
Origin, Migration, and Reproduction of Indigenous Domestic Animals with Special Reference to Their Sperm Quality


Abstract
:Simple Summary
Abstract
1. Introduction
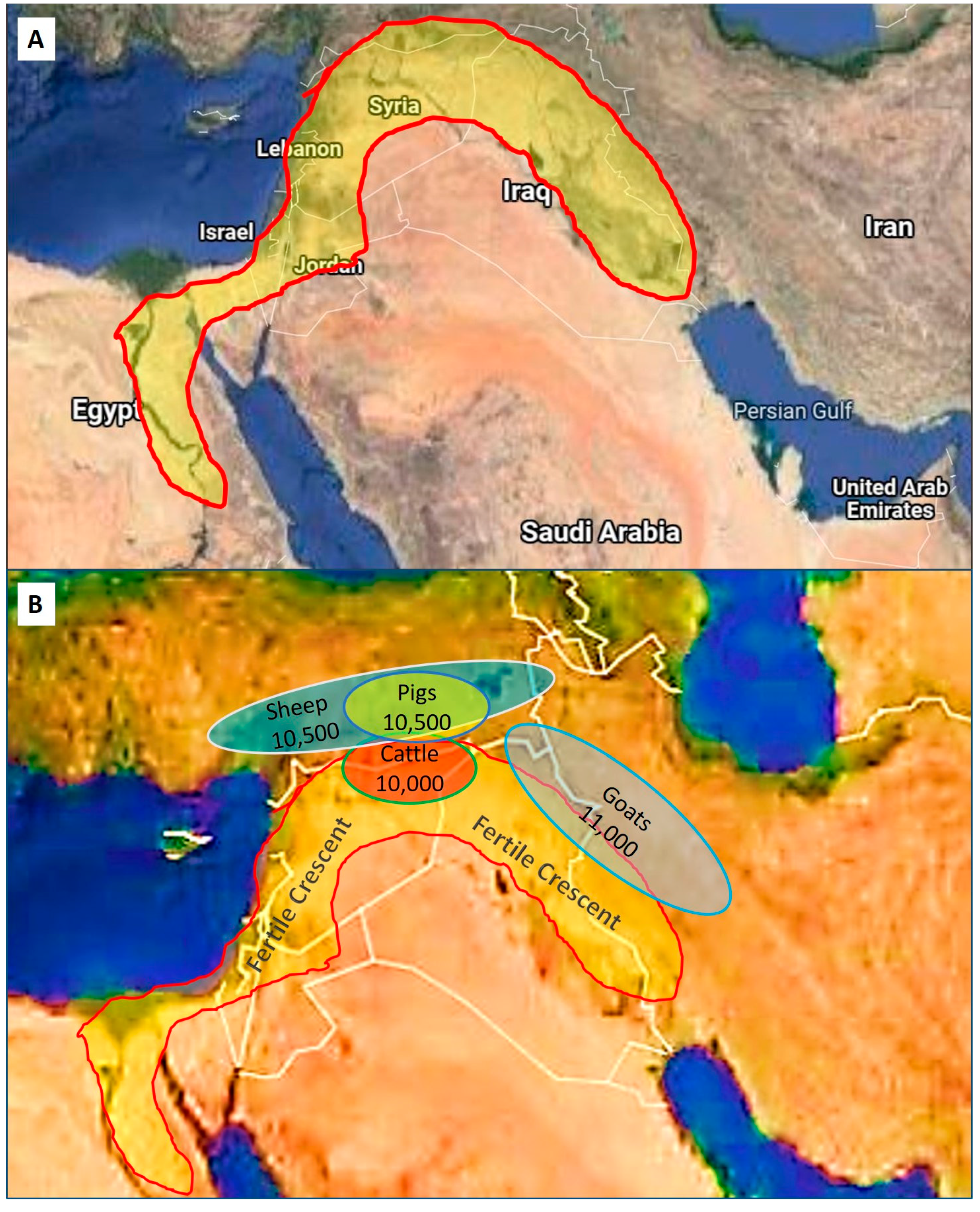
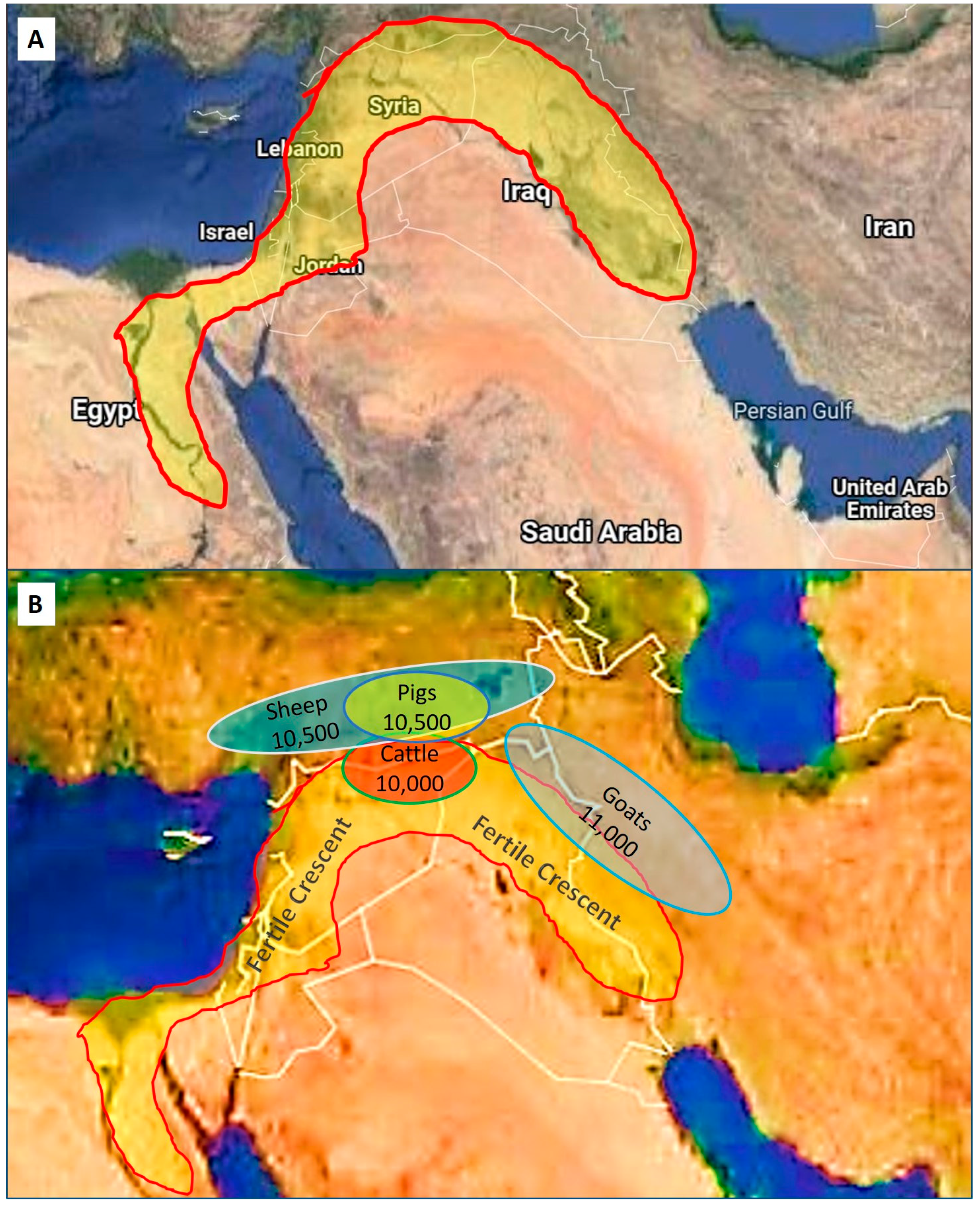
2. Historical Perspectives and Supportive Background
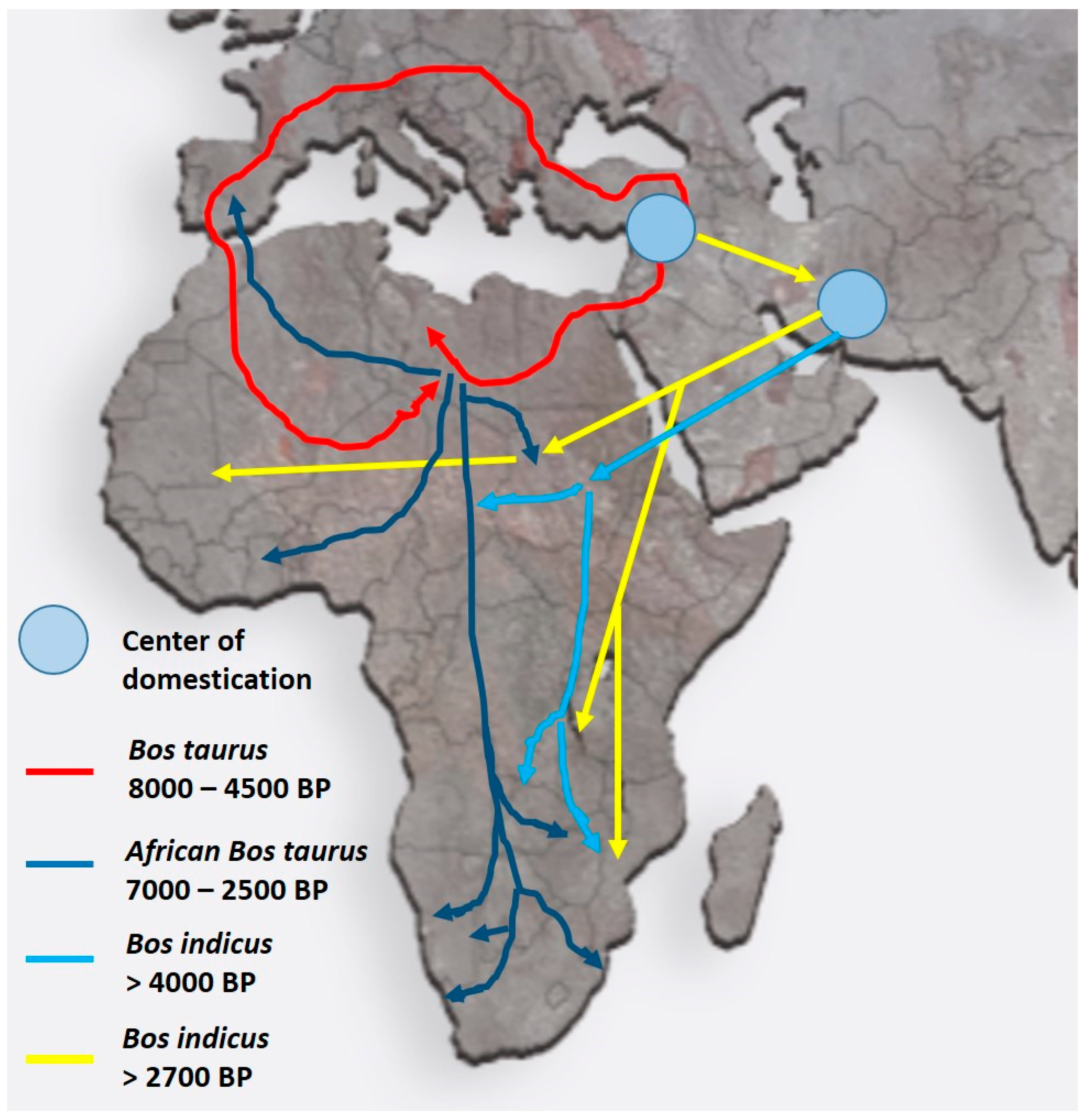
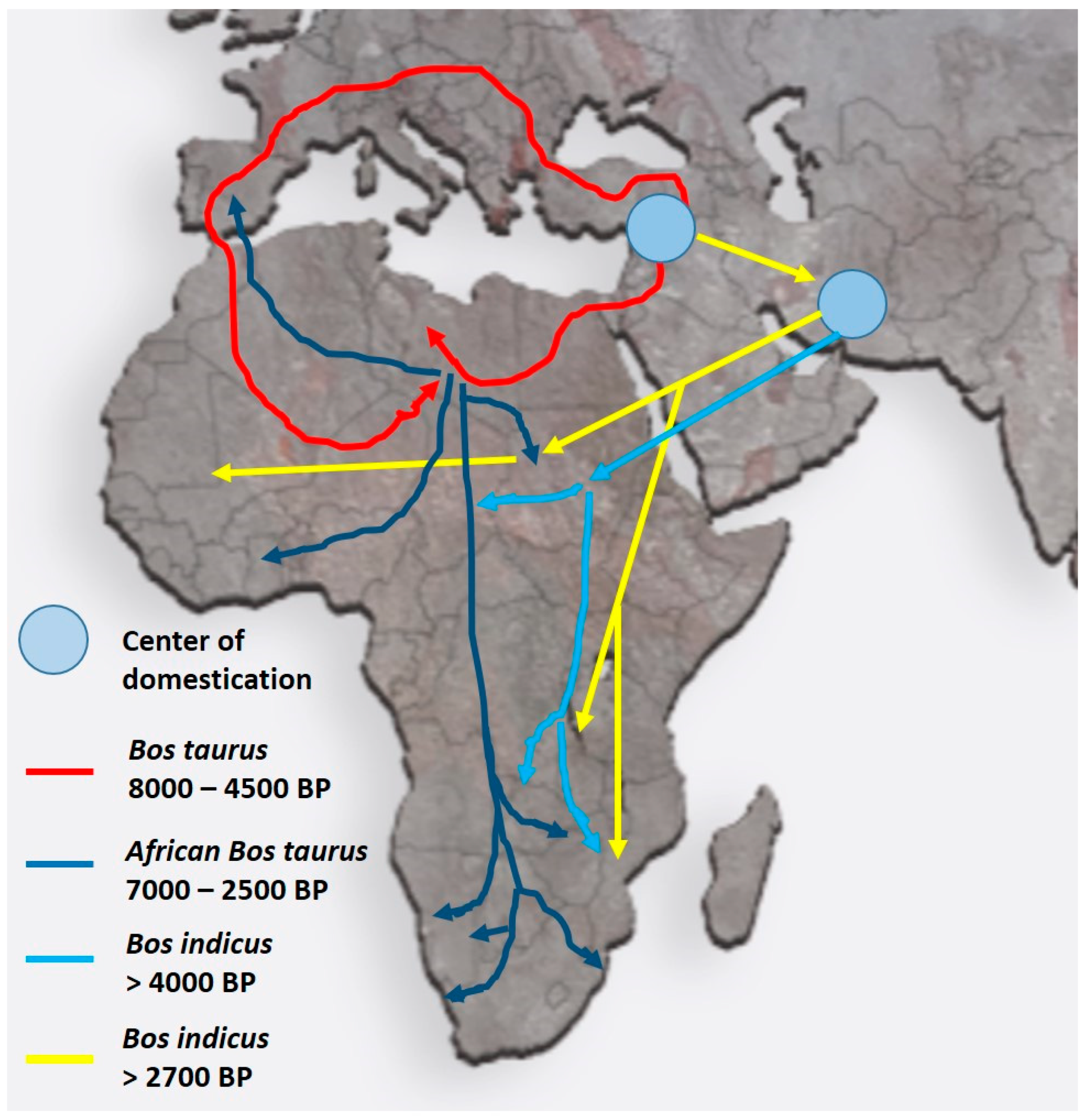
3. Migrations into Africa
4. Natural Selection vs. Artificial Selection for Reproduction
5. Advantages and Use of Indigenous Domestic Animals
6. Which Semen and Sperm Traits Have Been Recorded?
7. Do Semen Parameters Assessed during Breeding Soundness Relate to Fertility?
8. Is It Important to Protect Indigenous Domestic Breeds?
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Driscoll, C.A.; Macdonald, D.W.; O’Brien, S.J. From wild animals to domestic pets, an evolutionary view of domestication. Proc. Natl. Acad. Sci. USA 2009, 106, 9971–9978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daly, N. Domestic Animals Explained. National Geographic, 4 July. 2019. Available online: https://www.nationalgeographic.com/animals/article/domesticated-animals (accessed on 14 December 2021).
- Illsley, C.L. What Are Native, Indigenous and Endemic Species. WorldAtlas, 25 September. 2018. Available online: https://www.worldatlas.com/articles/what-is-the-difference-between-a-native-an-indigenous-and-an-endemic-species.html (accessed on 14 December 2021).
- Zeder, M.A.; Emshwiller, E.; Smith, B.D.; Bradley, D.G. Documenting Domestication: New Genetic and Archaeological Paradigms; University of California Press: Berkeley, CA, USA, 2006; ISBN 9780520246386. [Google Scholar]
- Zeder, M.A. Domestication and early agriculture in the Mediterranean Basin: Origins, diffusion, and impact. Proc. Natl. Acad. Sci. USA 2008, 105, 11597–11604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, V.; Strömbergh, C.A.E.; Alimohammadian, A.; Sahni, A. Dinosaur coprolites and the early evolution of grasses and grazers. Science 2015, 310, 1177–1180. [Google Scholar] [CrossRef] [Green Version]
- Brugal, J.; Croitor, R. Evolution, ecology and biochronology of herbivore associations in Europe during the last 3 million years. Quaternaire 2007, 18, 129–152. [Google Scholar] [CrossRef] [Green Version]
- Ozimec, R. The importance of indigenous domestic animal breeds for the conservation karst polje grassland habitats. In Proceedings of the International Workshop on Dinaric Karst Fields as Wetlands of National and International Importance, Livno, Bosnia and Herzegovina, 1–3 October 2013. [Google Scholar]
- Gregory, T.R. Artificial selection and domestication: Modern lessons from Darwin’s enduring analogy. Evol. Educ. Outreach 2009, 2, 5–27. [Google Scholar] [CrossRef] [Green Version]
- Larson, G.; Piperno, D.R.; Allaby, R.G.; Purugganan, M.D.; Andersson, L.; Arroyo-Kalin, M.; Barton, L.; Vigueira, C.C.; Denham, T.; Dobney, K.; et al. Current perspectives and the future of domestication studies. Proc. Natl. Acad. Sci. USA 2014, 111, 6139–6146. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, H.I.; Ahmad, M.J.; Jabbir, F.; Ahmar, S.; Ahmad, N.; Elokil, A.A.; Chen, J. The domestication makeup: Evolution, survival, and challenges. Front. Ecol. Evol. 2020, 8, 103. [Google Scholar] [CrossRef]
- Mwai, O.; Hanotte, O.; Kwon, Y.; Cho, S. African indigenous cattle: Unique genetic resources in a rapidly changing world. Asian-Autralas. J. Anim. Sci. 2015, 28, 911–921. [Google Scholar] [CrossRef] [Green Version]
- Nyamushamba, G.B.; Mapiye, C.; Tada, O.; Halimani, T.E.; Muchenje, V. Conservation of indigenous cattle genetic resources in Southern Africa’s smallholder areas: Turning threats into opportunities—A review. Asian-Australas. J. Anim. Sci. 2017, 30, 603–621. [Google Scholar] [CrossRef]
- Molotsi, A.H.; Dube, B.; Cloete, S.W.P. The current status of indigenous ovine genetic resources in southern Africa and future sustainable utilisation to improve livelihoods. Diversity 2020, 12, 14. [Google Scholar] [CrossRef] [Green Version]
- Monau, P.; Raphaka, K.; Zvinorova-Chimboza, P.; Gondwe, T. Sustainable utilization of indigenous goats in Southern Africa. Diversity 2020, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Grobbelaar, J.A.N.; Sutherland, B.; Molalakgotla, N.M. Egg production potentials of certain indigenous chicken breeds from South Africa. Anim. Genet. Resour. 2010, 46, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Darwin, C. On the Origin of Species by Mean of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life; John Murray: London, UK, 1859. [Google Scholar]
- Darwin, C. The Variation of Animals and Plants under Domestication; John Murray: London, UK, 1868. [Google Scholar]
- Rischkowsky, B.; Pilling, D. The State of the World’s Animal Genetic Resources for Food and Agriculture; FAO: Rome, Italy, 2007; ISBN 978-92-5-105762-9. [Google Scholar]
- Parker, G.A. Sperm competition and its evolutionary consequences in the insects. Biol. Rev. 1970, 45, 525–567. [Google Scholar] [CrossRef]
- van der Horst, G.; Maree, L. Sperm form and function in the absence of sperm competition. Mol. Reprod. Dev. 2014, 8, 204–216. [Google Scholar] [CrossRef]
- Diamond, J. Evolution, consequences and future of plant and animal domestication. Nature 2002, 418, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Dzama, K. Is the Livestock Sector in Southern Africa Prepared for Climate Change; South African Institute of International Affairs (SAIIA) Policy Briefing: Johannesburg, South Africa, 2016; Volume 153, pp. 1–4. [Google Scholar]
- Poland, M.; Hammond-Tooke, D.; Voigt, L. The Abundant Herds: A Celebration of the Nguni Cattle of the Zulu People; Fernwood Press: Vlaeberg, South Africa, 2003; ISBN 978-1-874950-69-1. [Google Scholar]
- Butler, M.L.; Bormann, J.M.; Weaber, R.L.; Grieger, D.M.; Rolf, M.M. Selection for bull fertility: A review. Transl. Anim. Sci. 2020, 4, 423–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utt, M.D. Prediction of bull fertility. Anim. Reprod. Sci. 2016, 169, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi-Arpanahi, R.; Morota, G.; Penagaricano, F. Predicting bull fertility using genomic data and biological information. J. Dairy Sci. 2017, 100, 9656–9666. [Google Scholar] [CrossRef] [Green Version]
- Usman, T.H.; Sir, S.M.; Sani, M.B. Comparison of semen characteristics between indigenous and Amo cockerel of Gombe Sate, Nigeria. Malays. J. Fundam. Appl. Sci. 2019, 303–306. [Google Scholar] [CrossRef]
- Mavi, G.K.; Dubey, P.P.; Cheema, R.S. Comparison of sperm attributes in two indigenous layer breeds and their relationship with fertility. Theriogenology Insight Int. J. Reprod. All Anim. 2017, 7, 123–129. [Google Scholar] [CrossRef]
- Makhafola, M.B.; Lehloenya, K.C.; Mphaphathi, M.L.; Dinnyes, A.; Nedambale, T.L. The effect of breed on the survivability and motility rate of cryopreserved cock semen. S. Afr. J. Anim. Sci. 2009, 39 (Suppl. 1), 242–245. [Google Scholar] [CrossRef]
- Molekwa, J.T.; Umesiobi, D.O. Relationships between cock semen viability and the fertility of artificially inseminated South African indigenous chicken breeds. S. Afr. J. Anim. Sci. 2009, 39 (Suppl. 1), 24–28. [Google Scholar] [CrossRef]
- Masenya, M.B.; Mphaphathi, M.L.; Mapeka, M.H.; Munyai, P.H.; Makhafola, M.B.; Ramukhithi, F.V.; Malusi, P.P.; Umesiobi, D.O.; Nedambale, T.L. Comparative study on semen characteristics of Kolbroek and Large White boars following computer aided sperm analysis® (CASA). Afr. J. Biotechnol. 2011, 10, 14223–14229. [Google Scholar] [CrossRef]
- Seshoka, M.M.; Mphaphathi, M.L.; Nedambale, T.L. Comparison of four different permitting and combination of two best cryoprotectants on freezing Nguni sperm evaluated with the aid of computer aided sperm analysis. Cryobiology 2016, 72, 232–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isnaini, N.; Wahjuningsih, S.; Adhitama, E. Seasonal effects on semen quality of Ongole crossbred and Simmental bulls used for artificial insemination. Livest. Res. Rural Dev. 2019, 31, 16. [Google Scholar]
- Bandeira, A.M.P.; Matos, J.E.; Maria, A.N.; Carneiro, P.C.F.; Purdy, P.H.; Azevedo, H.C. The effects of gelatin supplementation prior to cooling on ram semen quality and fertility. Anim. Reprod. 2018, 15, 23–28. [Google Scholar] [CrossRef]
- Boshoff, N.H.; Lambrechts, H.; Maree, L.; Cloete, S.W.P.; van der Horst, G. A novel flush technique to simulate natural dispersal of spermatozoa in the female reproductive tract and expedite motility assessment of fresh ejaculated Merino (Ovis aries) sperm. S. Afr. J. Anim. Sci. 2018, 48, 469–476. [Google Scholar] [CrossRef]
- Letsoalo, P.T. Characterisation and Cryopreservation of Semen from the Indigenous Namaqua Afrikaner Sheep Breed, in Comparison with the Dorper and Dohne Merino Breeds. Master’s Thesis, University of Fort Hare, Alice, South Africa, 2017. [Google Scholar]
- van der Horst, G. Computer aided sperm analysis (CASA) in domestic animals: Current status, three D tracking and flagellar analysis. Anim. Reprod. Sci. 2020, 220, 106350. [Google Scholar] [CrossRef]
- Sundararaman, M.N.; Edwin, M.J. Changes in Motility characteristics of goat spermatozoa during glycerol-equilibration and the relevance to cryopreservation. Asian J. Cell Biol. 2008, 3, 22–33. [Google Scholar] [CrossRef] [Green Version]
- Ncgauzele, A.; van der Horst, G.; Kotze, A.; Jonker, T.; Maree, L. Quantitative sperm characteristics of Tankwa goats with special reference to hyperactivated motility. S. Afr. J. Anim. Sci. 2020, 50, 687–699. [Google Scholar] [CrossRef]
- Kumaresan, A.; Elango, K.; Datta, T.K.; Morrell, J.M. Cellular and molecular insight into the etiology of subfertility/infertility in crossbred bulls (Bos taurus × Bos indicus): A review. Front. Cell Dev. Biol. 2021, 9, 696637. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.K.M.; Kumaresan, A.; Rajak, S.K.; Tajmul, M.; Datta, T.K.; Mohanty, T.K.; Srinivasan, A.; Yadav, S. Comparative proteomic analysis of Taurine, Indicine, and crossbred (Bos taurus × Bos indicus) bull spermatozoa for identification of proteins related to sperm malfunctions and subfertility in crossbred bulls. Theriogenology 2015, 84, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Goss, D.M.; Ayad, B.; Vlok, M.; Hattingh, S.M.; van der Horst, G.; du Plessis, S.S. Proteomics and biomarker identification in improved sperm motility parameters after 4 h of ejaculatory abstinence. In XIIIth International Symposium on Spermatology; Björndahl, L., Flanagan, J., Holmberg, R., Kvist, U., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Alipour, H.; Duus, R.K.; Wimmer, R.; Dardmeh, F.; Du Plessis, S.S.; Jørgensen, N.; Christiansen, O.B.; Hnida, C.; Nielsen, H.I.; Van Der Horst, G. Seminal plasma metabolomics profiles following long (4–7 days) and short (2 h) sexual abstinence periods. Eur. J. Obstet. Gynecol. Reprod. Biol. 2021, 264, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Patterson, D.; Perry, G. Determining Reproductive Fertility in Herd Bulls; University of Missouri Extension: Columbia, MO, USA, 2011; pp. 1–4. [Google Scholar]
- Tibary, A.; Boukhliq, R.; Allali, K. Ram and buck breeding soundness examination. Rev. Mar. Sci. Agron. Vét. 2018, 6, 241–255. [Google Scholar]
- Keyser, S.; van der Horst, G.; Maree, L. New approaches to define the functional competency of human sperm subpopulations and its relationship to semen quality. Int. J. Fertil. Steril. 2021; in press. [Google Scholar]
- Nagy, A.; Polichronopoulos, T.; Gáspárdy, A.; Solti, L.; Cseh, S. Correlation between bull fertility and sperm cell velocity parameters generated by computer-assisted semen analysis. Acta Vet. Hung. 2015, 63, 370–381. [Google Scholar] [CrossRef] [Green Version]
- van der Horst, G.; du Plessis, S.S. Not just the marriage of figaro: But the marriage of WHO/ESHRE semen analysis criteria with sperm functionality. Postęp. Androl. Online 2017, 4, 6–21. [Google Scholar]
- van der Horst, G.; Maree, L.; du Plessis, S.S. Current perspectives of CASA applications in diverse mammalian sperm. Reprod. Fertil. Dev. 2018, 30, 875–888. [Google Scholar] [CrossRef] [Green Version]
- Petrunkina, A.M.; Waberski, D.; Günzel-Apel, A.R.; Töpfer-Petersen, E. Determinants of sperm quality and fertility in domestic species. Reproduction 2007, 134, 3–17. [Google Scholar] [CrossRef]
- Keyser, S.; van der Horst, G.; Maree, L. Progesterone, myo-inositol, dopamine and prolactin present in follicular fluid have differential effects on sperm motility subpopulations. Life 2021, 11, 1250. [Google Scholar] [CrossRef]
- Erenpreiss, J.; Jepson, K.; Giwercman, A.; Tsarev, I.; Erenpreisa, J.; Spano, M. Toluidine blue cytometry test for sperm DNA conformation: Comparison with the flow cytometric sperm chromatin structure and TUNEL assays. Hum. Reprod. 2004, 19, 2277–2282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dovie, B.D.K.; Shackleton, C.M.; Witkowski, E.T.F. Valuation of communal area livestock benefits, rural livelihoods and related policy issues. Land Use Policy 2006, 23, 260–271. [Google Scholar] [CrossRef]
- Simela, L.; Montshwe, B.D.; Mahanjana, A.M.; Tshuwa, M.P. The livestock production environment in the South African smallholder sector. New Challenges for the Animal Science Industries. In Proceedings of the SASAS 41st Congress, Bloemfontein, South Africa, 3–6 April 2006; p. 66. [Google Scholar]
- Bezjian, M.; Abou-Madi, N.; Kollias, G.V.; Parks, J.E.; Cheong, S.H.; Beltaire, K.A. Characterization and cryopreservation of semen from endangered markhor goats (Capra falconeri heptneri) with evaluation of reproductive seasonality. J. Zoo Wildl. Med. 2013, 44, 672–685. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Martinez, H. State of the art in farm animal sperm evaluation. Reprod. Fertil. Dev. 2007, 19, 91–101. [Google Scholar] [CrossRef] [PubMed]
- van der Horst, G.; Maree, L. Important Focus on Sperm of Indigenous Domestic Animals in Africa. Microptic Blog. 17 June 2021. Available online: https://www.micropticsl.com/indigenous-domestic-animals/ (accessed on 14 December 2021).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Palaeolithic Era | Mesolithic Era | Neolithic Era | |
|---|---|---|---|
| Circa 70,000–12,200 BP | Circa 12,000–10,500 BP | 11,000–3800 BP | |
| Methods of obtaining food | Hunting | Hunting | Raising and herding animals |
| Gathering fruits and grains | Gathering plants and storing food | Farming in permanent villages | |
| Domestication of animals | Symbiosis of humans and wolves | First attempts at domestication and taming of “wild” animals | Domestication of most livestock species |
| Domestic Animal | Scientific Name | Domesticated BP | Geographical Area |
|---|---|---|---|
| Dog | Canis familiaris | 36,000–15,000 | Mainly Eurasia |
| Goat | Capra hircus | 11,000 | Middle East |
| Sheep | Ovis aries | 10,500 | Middle East |
| Pig | Sus domesticus | 10,500 | Middle East |
| Cattle | Bos taurus/Bos indicus | 10,000 | Middle East |
| Cat | Felis silvestris catus | 9500 | Middle East |
| Horse | Equus caballus | 5500 | Asia |
| Chicken | Gallus gallus domesticus | 4000 | Asia |
| Nigeria [28] | India [29] | South Africa [30,31] | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Indigenous | Amo | Aseel | Kadaknath | Naked Neck | Ovambo | P Koekoek | Venda | W Leghorn * | |
| Semen and Sperm Traits | (n = 9) | (n = 9) | (n = 10) | (n = 10) | (n = 8) | (n = 8) | (n = 8) | (n = 8) | (n = 6) |
| Macroscopic | |||||||||
| Semen colour (Scale 1–4) | 2.85 ± 0.07 | 2.0 ± 0.9 | Creamy white | Creamy white | − | − | − | − | − |
| Semen pH | 7.0 ± 0.0 | 7.0 ± 0.0 | 7.1 | 7.2 | − | − | − | − | 6.9 |
| Semen volume (mL) | 0.21 ± 0.17 | 0.2 ± 0.2 | − | − | 0.3 | 0.4 | 0.7 | 0.2 | 0.2 |
| Microscopic | |||||||||
| Mass motility (Score 1–5) | 4.85 ± 0.27 | 4.37 ± 0.19 | − | − | − | − | − | − | − |
| Total motility (%) | − | − | 75.8 ± 7.99 | 75.7 ± 4.1 | 65 ± 1.3 | 80 ± 9.4 | 90 ± 9.4 | 96 ± 5.3 | 80 ± 9.4 |
| Progressive motility (%) | 95.0 ± 0.43 | 82.0 ± 1.15 | − | − | − | − | − | − | − |
| Sperm concentration (×109/mL) | 4.93 ± 1.84 | 3.4 ± 1.07 | − | − | 1.3 ± 0.01 | 2.1 ± 0.1 | 3.5 ± 0.04 | 2.1 ± 0.45 | 5.61 |
| Live sperm (%) | 88.9 ± 0.58 | 72.7 ± 0.4 | 78.4 ± 5.37 | 82.0 ± 4.47 | 54.6 ± 0.6 | 72.9 ± 0.9 | 88.4 ± 0.7 | 50.3 | 56 |
| Normal morphology (%) | 81.0 ± 0.78 | 66.2 ± 0.1 | 90.0 ± 3.5 | 93.0 ± 2.8 | − | − | − | − | − |
| Kinematics | |||||||||
| Average VCL (µm/s) | − | − | − | − | − | − | − | 125.8 | 55 |
| Average VSL (µm/s) | − | − | − | − | − | − | − | 75.5 | 23 |
| Average VAP (µm/s) | − | − | − | − | − | − | − | 93.4 | 39 |
| Boar [32] | Bull [33,34] | ||||
|---|---|---|---|---|---|
| SA Kolbroek | SA Large White | Nguni | Bonsmara | Ongole Crossbr | |
| Semen and Sperm Traits | n = 4 | n = 4 | n = 4 | n = 4 | n = 1 |
| Macroscopic | |||||
| Semen pH | 7.0 ± 0.0 | 7.0 ± 0.0 | 7.1 ± 0.2 | 7.4 ± 0.3 | 6.3 |
| Semen volume (mL) | 140.4 ± 48.6 | 177.5 ± 60.4 | 3.7 ± 1.8 | 4.5 ± 0.8 | 6.4 |
| Microscopic | |||||
| Total motility (%) | 95.2 ± 4.2 | 91.4 ± 6.2 | 93.0 ± 1.7 | 91.3 ± 4.2 | 70.6 |
| Progressive motility (%) | 36.8 ± 15.2 | 22.8 ± 6.8 | 48.6 ± 12.4 | 35.3 ± 15.5 | − |
| Rapid motility (%) | 55.2 ± 17.3 | 42.0 ± 13.4 | 83.6 ± 7.1 | 54.7 ± 22.2 | − |
| Sperm concentration (×109/mL) | 0.72 ± 3.4 | 0.76 ± 3.7 | 4.95 ± 1.89 | 4.88 ± 4.2 | 8.5 |
| Live sperm (%) | 84.6 ± 6.1 | 81.7 ± 7.1 | 92.9 ± 4.2 | 92.7 ± 4.6 | − |
| Normal morphology (%) | − | − | 92.1 ± 4.4 | 89.4 ± 4.1 | − |
| Kinematics | |||||
| Average VCL (µm/s) | 143.3 ± 22.8 | 129.1 ± 24.8 | 168.0 ± 20.4 | 128.3 ± 27.8 | − |
| Average VSL (µm/s) | 40.9 ± 10.6 | 32.8 ± 7.0 | 76.6 ± 16.5 | 66.7 ± 18.1 | − |
| Average VAP (µm/s) | 90.6 ± 16.1 | 81.1 ± 17.7 | 113.7 ± 14.7 | 53.0 ± 23.8 | − |
| Average LIN (%) | 29.2 ± 9.7 | 25.7 ± 5.1 | 45.5 ± 7.5 | 52.7 ± 12.6 | − |
| Ram [35,36,37] | Goat [38,39,40] | |||||||
|---|---|---|---|---|---|---|---|---|
| Santa Inéz | Merino * | Namaqua | Dorper | Dohne Merino | Saanen * | Tankwa | Boer | |
| Semen and Sperm Traits | (n = 24) | (n = 10) | (n = 12) | (n = 10) | (n = 12) | (n = 3) | (n = 7) | (n = 3) |
| Macroscopic | ||||||||
| Semen pH | − | − | − | − | − | − | − | 6.94 |
| Semen volume (mL) | − | − | 1.09 ± 0.08 | 1.37± 0.08 | 1.2 ± 0.08 | 0.64 ± 0.11 | ||
| Microscopic | ||||||||
| Total motility (%) | − | 74.1 ± 10.7 | 70–85 | 70–85 | 70–85 | 74.1 ± 10.7 | 78.0 ± 12.5 | 78 |
| Sperm concentration (×106/mL) | − | 3846.1 ± 2773.0 | 1220 ± 5200 | 1100 ± 5290 | 1140 ± 5200 | 2306.1 ± 1669 | 2134.0 ± 1200 | 3.02 |
| Live sperm (%) | − | − | 67.67 ± 1.94 | 72.82 ± 1.98 | 68.59 ± 1.94 | − | − | 89.2 |
| Abnormal morphology (%) | − | − | − | − | − | − | − | 4.6 |
| Intact plasma membrane (%) | − | − | − | − | − | − | − | 88.2 |
| Kinematics | ||||||||
| Average VCL (µm/s) | 357 ± 30.7 | 231.3 ± 48.4 | − | − | − | 166.0 ± 36.4 | 157.0 ± 28.7 | 200.0 ± 14.1 |
| Rapid VCL (µm/s) | − | 246.6 ± 38.4 | − | − | − | 237.3 ± 44.4 | 225.0 ± 33.4 | − |
| Average VSL (µm/s) | 197.8 ± 20.6 | 182.3 ± 38.6 | − | − | − | 88.5 ± 33.6 | 88.9 ± 12.6 | 117.0 ± 5.94 |
| Rapid VSL (µm/s) | − | 188.9 ± 28.4 | − | − | − | 143.4 ± 28.4 | 165.0 ± 12.1 | − |
| Average VAP (%) | 249.5 ± 23.4 | 208.3 ± 39.1 | − | − | − | 104.0 ± 33.1 | 125.0 ± 18.5 | 126.0 ± 6.79 |
| Rapid VAP (%) | − | 211.9 ± 34.8 | − | − | − | 153.0 ± 27.8 | 178.0 ± 19.1 | − |
| Average LIN (%) | 55.5 ± 4.8 | 74.4 ± 12.0 | − | − | − | 54.3 ± 14.0 | 70.0 ± 11.1 | 58.3 ± 1.56 |
| Rapid LIN (%) | − | 82.5 ± 5.1 | − | − | − | 60.6 ± 5.1 | 73.8 ± 4.5 | − |
| Average STR (%) | 79.3 ± 3.5 | 87.2 ± 5.1 | − | − | − | 81.8 ± 7.2 | 87.4 ± 13.2 | 90.1 ± 0.69 |
| Rapid STR (%) | − | 89.3 ± 1.3 | − | − | − | 93.4 ± 2.4 | 92.7 ± 12.3 | − |
| Average WOB (%) | − | 84.8 ± 10.3 | − | − | − | 64.0 ± 10.3 | 79.5 ± 6.3 | − |
| Rapid WOB (%) | − | 92.4 ± 5.1 | − | − | − | 65.0 ± 5.1 | 79.7 ± 7.1 | − |
| Average ALH (µm) | 3.0 ± 0.2 | 3.6 ± 0.8 | − | − | − | 3.4 ± 0.7 | 2.3 ± 0.6 | 6.1 ± 0.29 |
| Rapid ALH (µm) | 2.3 ± 0.2 | 3.8 ± 0.9 | − | − | − | 4.5 ± 0.9 | 3.25 ± 0.6 | − |
| Average BCF (Hz) | 53.7 ± 2.0 | 17.4 ± 2.9 | − | − | − | 28.8 ± 3.1 | 32.3 ± 12.1 | 44.6 ± 0.57 |
| Rapid BCF (Hz) | − | 18.1 ± 3.2 | − | − | − | 28.7 ± 3.2 | 34.0 ± 15.9 | − |
| Hyperactivation (%) | − | − | − | − | − | − | 25.3 | − |
| Indigenous | Crossbreeds (with Simmental * (S)) | |||||
|---|---|---|---|---|---|---|
| Nguni (N) | Bonsmara (B) | Afrikaner (A) | S × N | S × B | S × A | |
| Sperm functionalities | (n = 2) | (n = 2) | (n = 2) | (n = 2) | (n = 2) | (n = 1) |
| Total motility (%) | 82.5 | 71 | 72 | 85 | 76 | 83 |
| Progressive motility (%) | 62 | 55 | 38.5 | 76 | 24.5 | 62 |
| Rapid progressive motility (%) | 11 | 10.6 | 8 | 8 | 11 | 12 |
| Rapid progressive VCL (µm/s) | 229.1 | 217 | 205 | 219 | 233 | 194 |
| Rapid progressive VAP (µm/s) | 130.2 | 108 | 115.8 | 98.3 | 122 | 108 |
| Rapid Progressive VSL (µm/s) | 115 | 98.2 | 101.5 | 85.9 | 109.5 | 95 |
| Rapid progressive LIN (%) | 50.9 | 45.1 | 49.5 | 39 | 47.2 | 49.3 |
| Hyperactivation (%) | 41.3 | 25 | 37 | 76 | 9 | 32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van der Horst, G.; Maree, L. Origin, Migration, and Reproduction of Indigenous Domestic Animals with Special Reference to Their Sperm Quality. Animals 2022, 12, 657. https://doi.org/10.3390/ani12050657
van der Horst G, Maree L. Origin, Migration, and Reproduction of Indigenous Domestic Animals with Special Reference to Their Sperm Quality. Animals. 2022; 12(5):657. https://doi.org/10.3390/ani12050657
Chicago/Turabian Stylevan der Horst, Gerhard, and Liana Maree. 2022. "Origin, Migration, and Reproduction of Indigenous Domestic Animals with Special Reference to Their Sperm Quality" Animals 12, no. 5: 657. https://doi.org/10.3390/ani12050657
APA Stylevan der Horst, G., & Maree, L. (2022). Origin, Migration, and Reproduction of Indigenous Domestic Animals with Special Reference to Their Sperm Quality. Animals, 12(5), 657. https://doi.org/10.3390/ani12050657