
High Polymorphism in the Dmrt2a Gene Is Incompletely Sex-Linked in Spotted Scat, Scatophagus argus




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
2.2. RNA Isolation and cDNA Synthesis
2.3. Genotypic and Phenotypic Sex Identification
2.4. Cloning of Dmrt2a and Dmrt2b in S. argus
2.5. Sequence Analysis and Bioinformatics
2.6. Designing of Markers on Dmrt2a Genomic DNA Sequence
2.7. Tissue Distribution
2.8. Real-Time Quantitative PCR (qPCR)
2.9. Dmrt2a and Dmrt2b Expression in Transcriptome Data of Different Fish Species
3. Results
3.1. Cloning and Sequence Analysis of Spotted Scat Dmrt2a and Dmrt2b
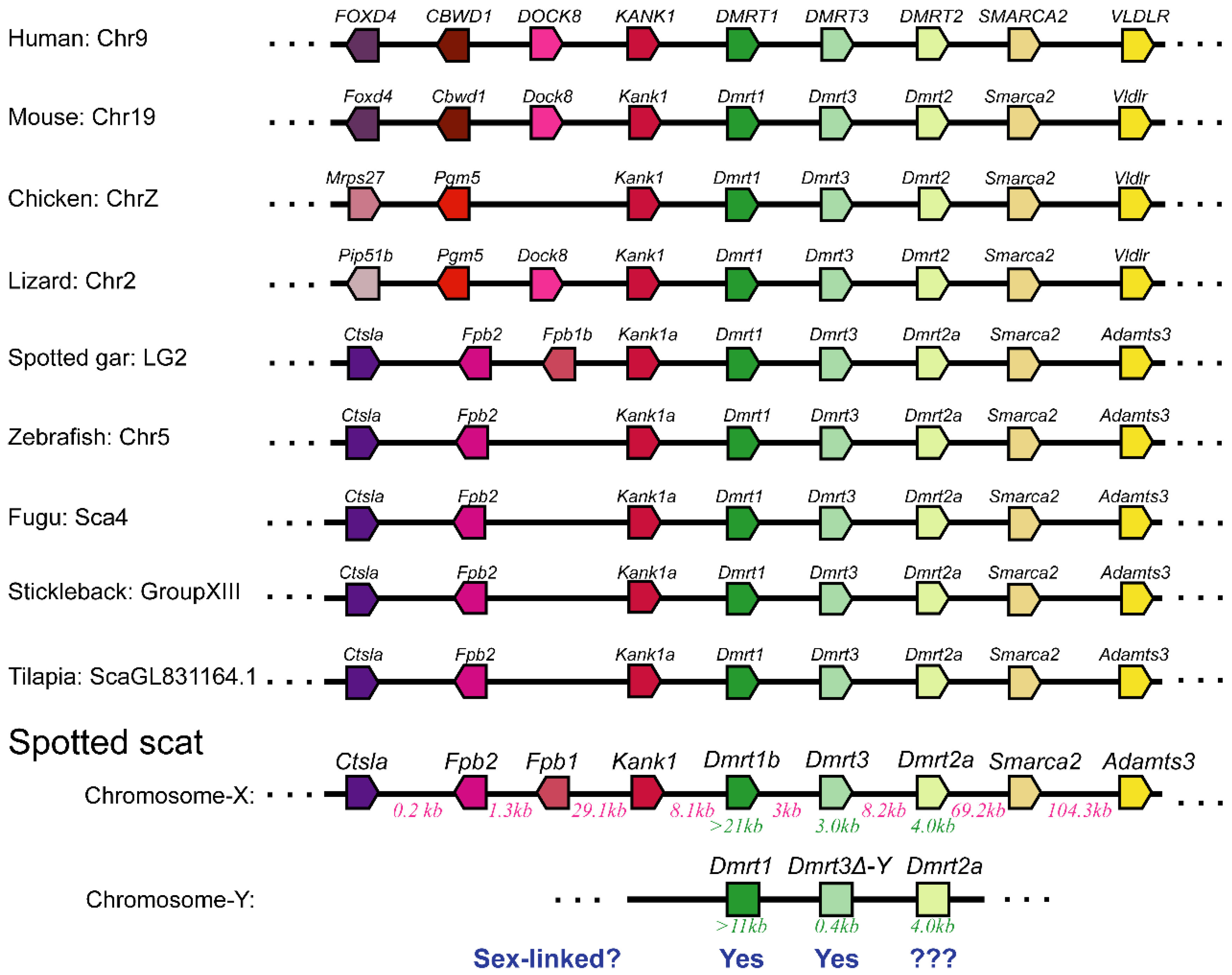
3.2. The Dmrt2a Genomic Sequence in Female and Male Spotted Scat Is Dissimilar
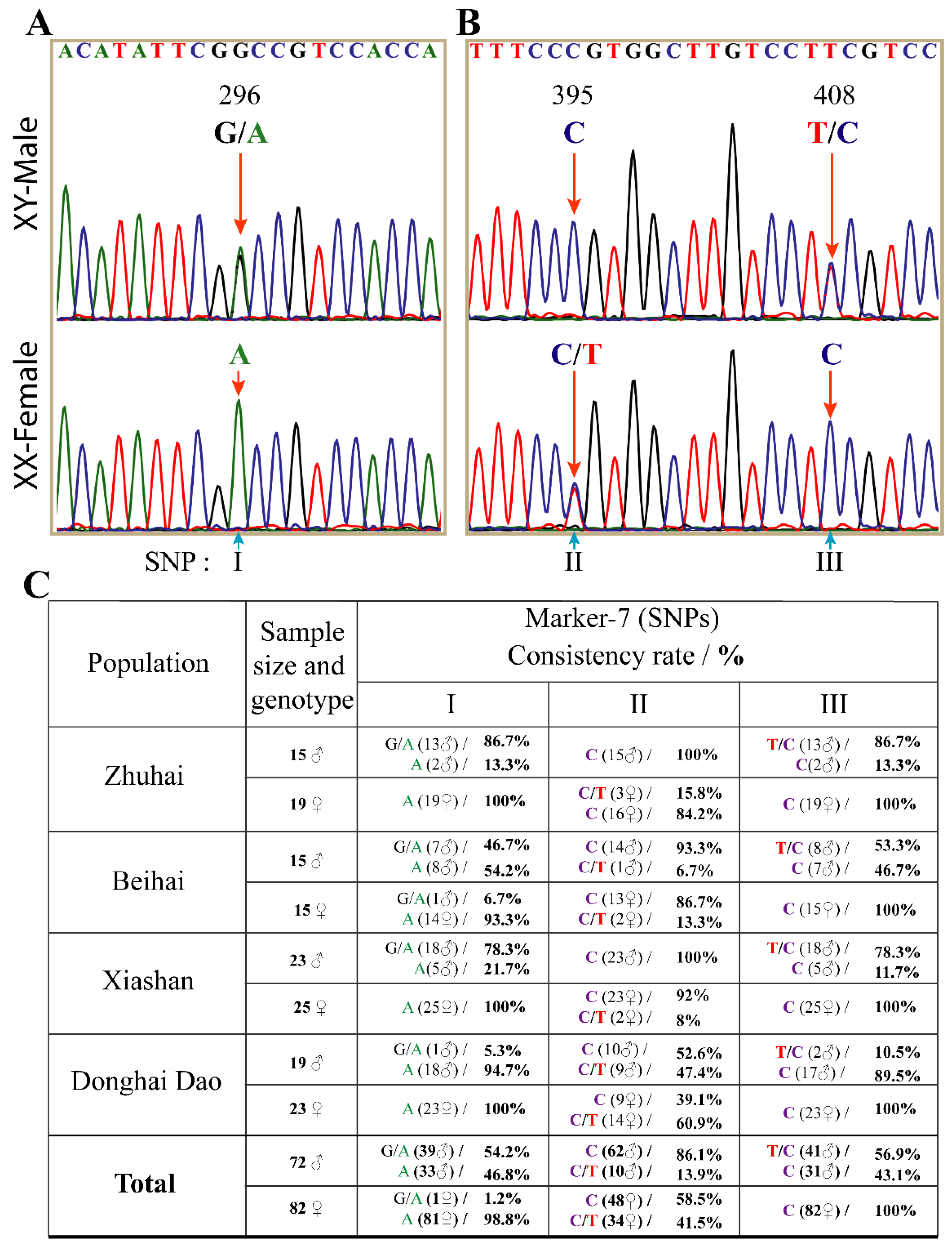
3.3. The Interspersed Indels and SNPs on Dmrt2a Are Sex-Linked at Different Rates
3.4. Dmrt2a Is Highly Expressed in Spotted Scat Testes
3.5. The Expression of Dmrt2a in Spotted Scat Gonads
4. Discussion
4.1. The Spotted Scat and the Incomplete Sex-Linked Dmrt2a Gene Are Excellent Models to Study the Evolution of Sex Chromosomes and the Differentiation of Sex-Linked Genes
4.2. Reason for the Sexually Dimorphic Expression of Dmrt2 in the Gonads of Spotted Scat
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R.; et al. Sex determination: Why so many ways of doing it? PLoS Biol. 2014, 12, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, J.; Gui, J.F. Genetic basis and biotechnological manipulation of sexual dimorphism and sex determination in fish. Sci. China Life Sci. 2015, 58, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Mustapha, U.F.; Jiang, D.N.; Liang, Z.H.; Gu, H.-T.; Yang, W.; Chen, H.P.; Deng, S.; Wu, T.L.; Tian, C.-X.X.; Zhu, C.H.; et al. Male-specific dmrt1 is a candidate sex determination gene in spotted scat (Scatophagus argus). Aquaculture 2018, 495, 351–358. [Google Scholar] [CrossRef]
- Bao, L.; Tian, C.; Liu, S.; Zhang, Y.; Elaswad, A.; Yuan, Z.; Khalil, K.; Sun, F.; Yang, Y.; Zhou, T.; et al. The Y Chromosome sequence of the channel catfish suggests novel sex determination mechanisms in teleost fish. BMC Biol. 2019, 17, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Sun, Y.; Zhao, J.; Shi, H.; Zeng, S.; Ye, K.; Jiang, D.; Zhou, L.; Sun, L.; Tao, W.; et al. A tandem duplicate of anti-müllerian hormone with a missense SNP on the Y chromosome is essential for male sex determination in nile tilapia, Oreochromis niloticus. PLoS Genet. 2015, 11, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Pan, Q.; Feron, R.; Yano, A.; Jouanno, E.; Wen, M.; Bobe, J.; Concordet, J.-P.; Journot, L.; Klopp, C.; Schartl, M.; et al. Identification of the master sex determining gene in northern pike (Esox lucius) reveals restricted sex. bioRxiv 2019, 15, 549527. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, K.; Hamaguchi, S. Novel sex-determining genes in fish and sex chromosome evolution. Dev. Dyn. 2013, 242, 339–353. [Google Scholar] [CrossRef]
- Martínez, P.; Viñas, A.M.; Sánchez, L.; Díaz, N.; Ribas, L.; Piferrer, F. Genetic architecture of sex determination in fish: Applications to sex ratio control in aquaculture. Front. Genet. 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlesworth, D.; Charlesworth, B.; Marais, G. Steps in the evolution of heteromorphic sex chromosomes. Heredity 2005, 95, 118–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, M.W.; Lee, J.K.; Rojo, S.; Gearhart, M.D.; Kurahashi, K.; Banerjee, S.; Loeuille, G.A.; Bashamboo, A.; McElreavey, K.; Zarkower, D.; et al. An ancient protein-DNA interaction underlying metazoan sex determination. Nat. Struct. Mol. Biol. 2015, 22, 442–451. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Li, J.; Hu, J.; Sun, C.; Tian, Y.; Li, W.; Yan, N.; Sun, C.; Sheng, X.; Yang, S.; et al. Comparative genomics studies on the dmrt gene family in fish. Front. Genet. 2020, 11, 563947. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, H.; Andersen, Ø. Sex dimorphic expression of five dmrt genes identified in the atlantic cod genome. The fish-specific Dmrt2b diverged from Dmrt2a before the fish whole-genome duplication. Gene 2012, 505, 221–232. [Google Scholar] [CrossRef]
- Su, L.; Zhou, F.; Ding, Z.; Gao, Z.; Wen, J.; Wei, W.; Wang, Q.; Wang, W.; Liu, H. Transcriptional variants of Dmrt1 and expression of four dmrt genes in the blunt snout bream, Megalobrama amblycephala. Gene 2015, 573, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Portela-Bens, S.; Merlo, M.A.; Rodríguez, M.E.; Cross, I.; Manchado, M.; Kosyakova, N.; Liehr, T.; Rebordinos, L. Integrated gene mapping and synteny studies give insights into the evolution of a sex proto-chromosome in Solea senegalensis. Chromosoma 2017, 126, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Raghuveer, K.; Senthilkumaran, B. Identification of multiple Dmrt1s in catfish: Localization, dimorphic expression pattern, changes during testicular cycle and after methyltestosterone treatment. J. Mol. Endocrinol. 2009, 42, 437–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Mao, A.; Wang, P.; Ning, G.; Cao, Y.; Wang, Q. Endodermal pouch-expressed Dmrt2b Is important for pharyngeal cartilage formation. Biol. Open 2018, 7, bio035444. [Google Scholar] [CrossRef] [Green Version]
- Lyu, Q.; Hu, J.; Yang, X.; Liu, X.; Chen, Y.; Xiao, L.; Liu, Y.; Wang, Q.; Chen, J.; Huang, M.; et al. Expression profiles of dmrts and foxls during gonadal development and sex reversal induced by 17α-methyltestosterone in the orange-spotted grouper. Gen. Comp. Endocrinol. 2019, 274, 26–36. [Google Scholar] [CrossRef]
- Wexler, J.R.; Plachetzki, D.C.; Kopp, A. Pan-metazoan phylogeny of the DMRT gene family: A framework for functional studies. Dev. Genes Evol. 2014, 224, 175–181. [Google Scholar] [CrossRef]
- Hong, C.-S.; Park, B.-Y.; Saint-Jeannet, J.-P. The function of dmrt genes in vertebrate development: It is not just about sex. Dev. Biol. 2007, 310, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Brunner, B. Genomic organization and expression of the doublesex—Related gene cluster in vertebrates and detection of putative regulatory regions for DMRT1. Genomics 2001, 77, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Kettlewell, J.R.; Anderson, R.C.; Bardwell, V.J.; Zarkower, D. Sexually dimorphic expression of multiple doublesex-related genes in the embryonic mouse gonad. Gene Expr. Patterns 2003, 3, 77–82. [Google Scholar] [CrossRef]
- Kondo, M.; Froschauer, A.; Kitano, A.; Nanda, I.; Hornung, U.; Volff, J.-N.; Asakawa, S.; Mitani, H.; Naruse, K.; Tanaka, M.; et al. Molecular cloning and characterization of DMRT genes from the medaka oryzias latipes and the platyfish Xiphophorus maculatus. Gene 2002, 295, 213–222. [Google Scholar] [CrossRef]
- Winkler, C.; Hornung, U.; Kondo, M.; Neuner, C.; Duschl, J.; Shima, A.; Schartl, M. Developmentally regulated and non-sex-specific expression of autosomal dmrt genes in embryos of the medaka fish (Oryzias latipes). Mech. Dev. 2004, 121, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, H.; Li, M.; Cheng, Y.; Jiang, D.; Sun, L.; Tao, W.; Zhou, L.; Wang, Z.; Wang, D. Isolation of doublesex- and Mab-3-related transcription factor 6 and its involvement in spermatogenesis in tilapia1. Biol. Reprod. 2014, 91, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ottolenghi, C.; Fellous, M.; Barbieri, M.; McElreavey, K. Novel paralogy relations among human chromosomes support a link between the phylogeny of doublesex-related genes and the evolution of sex determination. Genomics 2002, 79, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, Q.; Lu, H.; Chen, H.; Guo, Y.; Cheng, H.; Zhou, R. Fish specific duplication of Dmrt2: Characterization of zebrafish Dmrt2b. Biochimie 2008, 90, 878–887. [Google Scholar] [CrossRef]
- Zheng, Y.; Liang, H.; Xu, P.; Li, M.; Wang, Z. Molecular cloning of Pcc-Dmrt1s and their specific expression patterns in pengze crucian carp (Carassius auratus var. pengze) affected by 17α-methyltestosterone. Fish Physiol. Biochem. 2014, 40, 1141–1155. [Google Scholar] [CrossRef]
- Yu, F.-F.; Wang, M.-F.; Zhou, L.; Gui, J.-F.; Yu, X.-Y. Molecular cloning and expression characterization of Dmrt2 in akoya pearl oysters, Pinctada Martensii. J. Shellfish Res. 2011, 30, 247–254. [Google Scholar] [CrossRef]
- Lourenço, R.; Lopes, S.S.; Saúde, L. Left-right function of Dmrt2 genes is not conserved between zebrafish and mouse. PLoS ONE 2010, 5, e14438. [Google Scholar] [CrossRef] [Green Version]
- Seo, K.W.; Wang, Y.; Kokubo, H.; Kettlewell, J.R.; Zarkower, D.A.; Johnson, R.L. Targeted disruption of the dm domain containing transcription factor Dmrt2 reveals an essential role in somite patterning. Dev. Biol. 2006, 290, 200–210. [Google Scholar] [CrossRef]
- Sheng, Y.; Chen, B.; Zhang, L.; Luo, M.; Cheng, H.; Zhou, R. Identification of dmrt genes and their up-regulation during gonad transformation in the swamp eel (Monopterus albus). Mol. Biol. Rep. 2014, 41, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Jiang, D.; Li, M.; Mustapha, U.F.; Tian, C.; Chen, H.; Huang, Y.; Deng, S.; Wu, T.; Zhu, C.; et al. Genome survey of male and female spotted scat (Scatophagus argus). Animals 2019, 9, 1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Mustapha, U.F.; Huang, Y.; Tian, C.; Yang, W.; Chen, H.; Deng, S.; Zhu, C.; Jiang, D.; Li, G. A chromosome—Level genome assemblyof the spotted scat (Scatophagus Argus). Genome Biol. Evol. 2021, 13, evab092. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.N.; Mustapha, U.F.; Shi, H.J.; Huang, Y.Q.; Si-Tu, J.X.; Wang, M.; Deng, S.P.; Chen, H.P.; Tian, C.X.; Zhu, C.-H.C.H.; et al. Expression and transcriptional regulation of Gsdf in spotted scat (Scatophagus argus). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2019, 233, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Chen, H.; Cui, X.; Zhang, K.; Jiang, D.; Deng, S.; Zhu, C.; Li, G. Sequencing, de novo assembly and characterization of the spotted scat Scatophagus argus (Linnaeus 1766) transcriptome for discovery of reproduction related genes and SSRs. J. Oceanol. Limnol. 2018, 36, 1329–1341. [Google Scholar] [CrossRef]
- He, F.X.; Jiang, D.N.; Huang, Y.Q.; Mustapha, U.F.; Yang, W.; Cui, X.F.; Tian, C.X.; Chen, H.P.; Shi, H.J.; Deng, S.P.; et al. Comparative transcriptome analysis of male and female gonads reveals sex-biased genes in spotted scat (Scatophagus argus). Fish Physiol. Biochem. 2019, 45, 1963–1980. [Google Scholar] [CrossRef]
- Mustapha, U.F.; Huang, Y.; Huang, Y.-Q.; Assan, D.; Shi, H.-J.; Jiang, M.-Y.; Deng, S.-P.; Li, G.-L.; Jiang, D.-N. Gonadal development and molecular analysis revealed the critical window for sex differentiation, and E2 reversibility of XY-male spotted scat, scatophagus argus. Aquaculture 2021, 544, 737147. [Google Scholar] [CrossRef]
- Dong, Z.; Li, X.; Yao, Z.; Wang, C.; Guo, Y.; Wang, Q.; Shao, C.; Wang, Z. Oryzias curvinotus in sanya does not contain the male sex- determining gene dmy. Animals 2021, 11, 1327. [Google Scholar] [CrossRef]
- Tian, C.; Li, Z.; Dong, Z.; Huang, Y.; Du, T.; Chen, H.; Jiang, D.; Deng, S.; Zhang, Y.; Wanida, S.; et al. Transcriptome analysis of male and female mature gonads of silver sillago (Sillago sihama). Genes 2019, 10, 129. [Google Scholar] [CrossRef] [Green Version]
- Tao, W.; Yuan, J.; Zhou, L.; Sun, L.; Sun, Y.; Yang, S.; Li, M.; Zeng, S.; Huang, B.; Wang, D. Characterization of gonadal transcriptomes from nile tilapia (Oreochromis niloticus) reveals differentially expressed genes. PLoS ONE 2013, 8, 63604. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Zhou, D.; Zhang, X.; Li, G.; Zhang, Y.; Huang, C.; Zhang, Z.; Tian, C. A First insight into the gonad transcriptome of Hong Kong catfish (Clarias fuscus). Animals 2021, 11, 1131. [Google Scholar] [CrossRef]
- Chen, H.; Li, Z.; Wang, Y.; Huang, H.; Yang, X.; Li, S.; Yang, W.; Li, G. Comparison of gonadal transcriptomes uncovers reproduction-related genes with sexually dimorphic expression patterns in diodon hystrix. Animals 2021, 11, 1042. [Google Scholar] [CrossRef]
- Graves, J.A.M. The rise and fall of SRY. Trends Genet. 2002, 18, 259–264. [Google Scholar] [CrossRef]
- Cortez, D.; Marin, R.; Toledo-Flores, D.; Froidevaux, L.; Liechti, A.; Waters, P.D.; Grützner, F.; Kaessmann, H. Origins and functional evolution of Y chromosomes across mammals. Nature 2014, 508, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Graves, M.J.A. Evolution of the testis-determining gene—The rise and fall of SRY. Novartis Found. Symp. 2002, 244, 86–101. [Google Scholar] [CrossRef]
- De Bello Cioffi, M.; Kejnovsk, E.; Marquioni, V.; Poltronieri, J.; Molina, W.F.; Diniz, D.; Bertollo, L.A.C. The key role of repeated DNAs in sex chromosome evolution in two fish species with ZW sex chromosome system. Mol. Cytogenet. 2012, 5, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, K.A.; Schach, U.; Ordaz, A.; Steinfeld, J.S.; Draper, B.W.; Siegfried, K.R. Dmrt1 Is Necessary for male sexual development in zebrafish. Dev. Biol. 2017, 422, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Inui, M.; Tamano, M.; Kato, T.; Takada, S. CRISPR/Cas9-Mediated simultaneous knockout of Dmrt1 and Dmrt3 does not recapitulate the 46,XY gonadal dysgenesis observed in 9p24.3 deletion patients. Biochem. Biophys. Rep. 2017, 9, 238–244. [Google Scholar] [CrossRef]
- Curzon, A.Y.; Shirak, A.; Dor, L.; Zak, T.; Perelberg, A.; Seroussi, E.; Ron, M. A duplication of the anti-müllerian hormone gene is associated with genetic sex determination of different oreochromis niloticus strains. Heredity 2020, 125, 317–327. [Google Scholar] [CrossRef]
- Liu, X.; Dai, S.; Wu, J.; Wei, X.; Zhou, X.; Chen, M.; Tan, D.; Pu, D.; Li, M.; Wang, D. Roles of anti-müllerian hormone and its duplicates in sex determination and germ cell proliferation of nile tilapia. Genetics 2021, iyab237. [Google Scholar] [CrossRef]
- Jiang, X.; Mu, B.; Huang, Z.; Zhang, M.; Wang, X.; Tao, S. Impacts of Mutation effects and population size on mutation rate in asexual populations: A simulation study. BMC Evol. Biol. 2010, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, R.A.; Almeida-Santos, J.; Lourenço, R.; Saúde, L. Identification of Dmrt2a downstream genes during zebrafish early development using a timely controlled approach. BMC Dev. Biol. 2018, 18, 4. [Google Scholar] [CrossRef] [Green Version]
- Ebbing, A.; Vértesy, Á.; Betist, M.C.; Spanjaard, B.; Junker, J.P.; Berezikov, E.; van Oudenaarden, A.; Korswagen, H.C. Spatial transcriptomics of C. elegans males and hermaphrodites identifies sex-specific differences in gene expression patterns. Dev. Cell 2018, 47, 801–813.e6. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.L.J.; Horsfield, J.A.; Black, M.A.; Rutherford, K.; Fisher, A.; Gemmell, N.J. Histological and transcriptomic effects of 17α-methyltestosterone on zebrafish gonad development. BMC Genom. 2017, 18, 557. [Google Scholar] [CrossRef] [PubMed]
- Ribas, L.; Robledo, D.; Gómez-Tato, A.; Viñas, A.; Martínez, P.; Piferrer, F. Comprehensive transcriptomic analysis of the process of gonadal sex differentiation in the turbot (Scophthalmus maximus). Mol. Cell. Endocrinol. 2016, 422, 132–149. [Google Scholar] [CrossRef]
- Kobayashi, T.; Matsuda, M.; Kajiura-Kobayashi, H.; Suzuki, A.; Saito, N.; Nakamoto, M.; Shibata, N.; Nagahama, Y. Two DM domain genes, DMY and DMRT1, involved in testicular differentiation and development in the medaka, Oryzias latipes. Dev. Dyn. 2004, 231, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.I.; Young, R.A. Transcriptional regulation and its misregulation in disease. Cell 2013, 152, 1237. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Sun, F.; Wan, Z.Y.; Yang, Z.; Tay, Y.X.; Lee, M.; Ye, B.; Wen, Y.; Meng, Z.; Fan, B.; et al. Transposon-induced epigenetic silencing in the X chromosome as a novel form of dmrt1 expression regulation during sex determination in the fighting fish. BMC Biol. 2022, 20, 1–16. [Google Scholar] [CrossRef]
- Barbosa, C.; Peixeiro, I.; Romão, L. Gene expression regulation by upstream open reading frames and human disease. PLoS Genet. 2013, 9, e1003529. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Sample Size and Genotype | Marker-2/Indel2-8 bp (Intron1) | Marker-3/Indel3-12 bp (Intron3) | ||
|---|---|---|---|---|---|
| Consistency | Average Concordance Rate (%) | Consistency | Average Concordance Rate (%) | ||
| Donghai Dao | a/b (1  ) ) | 5.3 | |||
| 19 | a/b (19 ) | 100 | b (18 ) | 94.7 | |
25  | a/b (25 ) | 100 | b (25 ) | 100 | |
| Beihai | 17 | a (10 ) a/b (7 ) | 58.8 41.2 | a (1 ) | 5.9 |
| a/b (7 ) | 41.2 | ||||
| b (9 ) | 52.9 | ||||
| 13 | a (5 ) | 38.5 | |||
| a/b (8 ) | 61.5 | b (13 ) | 100 | ||
| Xiashan | 24 | a (17 ) | 70.8 | a/b (18 ) | 75 |
| a/b (7 ) | 29.2 | b (6 ) | 25 | ||
| 24 | a (6 ) | 25 | b (24 ) | 100 | |
| a/b (11 ) | 45.8 | ||||
| b (7 ) | 29.2 | ||||
| Zhuhai | 15 | a (9 ) | 60 | a/b (13 ) | 86.7 |
| a/b (6 ) | 40 | b (2 ) | 13.3 | ||
| 19 | a (1 ) | 5.3 | b (19 ) | 100 | |
| a/b (9 ) | 47.4 | ||||
| b (9 ) | 47.4 | ||||
| TOTAL | 75 | a (36 ) a/b (39 ) | 48 52 | a (1 ) | 1.3 |
| a/b (39 ) | 52 | ||||
| b (35 ) | 46.7 | ||||
| 81 | a (12 ) | 14.8 | b (81 ) | 100 | |
| a/b (53 ) | 65.4 | ||||
| b (16 ) | 19.8 | ||||
| Population | Sample Size and Genotype | Marker-4/Indel4-99 bp (Intron1) | Sample Size and Genotype | Marker-6/Del6-14 bp (Intron2) | ||
|---|---|---|---|---|---|---|
| Consistency | Average Concordance Rate (%) | Consistency | Average Concordance Rate (%) | |||
| Donghai Dao | 23 | a (13 ) | 56.5 | a (3 ) | 14.3 | |
| a/b (10 ) | 43.5 | 21 | a/b (18 ) | 85.7 | ||
| 25 | a/b (2 ) | 8 | a (25 ) | 100 | ||
| b (23 ) | 92 | 25 | ||||
| Beihai | 17 | a (9 ) | 52.9 | a (7 ) | 46.7 | |
| a/b (8 ) | 47.1 | 15 | a/b (8 ) | 53.3 | ||
| 13 | a (2 ) | 15.4 | 15 | a (15 ) | 100 | |
| a/b (6 ) | 46.2 | |||||
| b (5 ) | 48.5 | |||||
| Xiashan | 24 | a (9 ) | 37.5 | a (16 ) | 64 | |
| a/b (15 ) | 62.5 | 25 | a/b (9 ) | 36 | ||
| 24 | a (4 ) | 16.7 | 30 | a (30 ) | 100 | |
| a/b (8 ) | 33.3 | |||||
| b (12 ) | 50 | |||||
| Zhuhai | 16 | a (4 ) | 25 | a (14 ) | 87.5 | |
| a/b (12 ) | 75 | 16 | a/b (2 ) | 12.5 | ||
| 22 | a (1 ) | 4.5 | 23 | a (23 ) | 100 | |
| a/b (2 ) | 9.1 | |||||
| b (19 ) | 86.4 | |||||
| TOTAL | 80 | a (35 ) | 43.7 | a (40 ) | 51.9 | |
| a/b (45 ) | 56.3 | 77 | a/b (37 ) | 48.1 | ||
| 84 | a (7 ) | 8.3 | 93 | a (93 ) | 100 | |
| a/b (18 ) | 21.4 | |||||
| b (59 ) | 70.2 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mustapha, U.F.; Assan, D.; Huang, Y.-Q.; Li, G.-L.; Jiang, D.-N. High Polymorphism in the Dmrt2a Gene Is Incompletely Sex-Linked in Spotted Scat, Scatophagus argus. Animals 2022, 12, 613. https://doi.org/10.3390/ani12050613
Mustapha UF, Assan D, Huang Y-Q, Li G-L, Jiang D-N. High Polymorphism in the Dmrt2a Gene Is Incompletely Sex-Linked in Spotted Scat, Scatophagus argus. Animals. 2022; 12(5):613. https://doi.org/10.3390/ani12050613
Chicago/Turabian StyleMustapha, Umar Farouk, Daniel Assan, Yuan-Qing Huang, Guang-Li Li, and Dong-Neng Jiang. 2022. "High Polymorphism in the Dmrt2a Gene Is Incompletely Sex-Linked in Spotted Scat, Scatophagus argus" Animals 12, no. 5: 613. https://doi.org/10.3390/ani12050613
APA StyleMustapha, U. F., Assan, D., Huang, Y.-Q., Li, G.-L., & Jiang, D.-N. (2022). High Polymorphism in the Dmrt2a Gene Is Incompletely Sex-Linked in Spotted Scat, Scatophagus argus. Animals, 12(5), 613. https://doi.org/10.3390/ani12050613