
A Review on the Marek’s Disease Outbreak and Its Virulence-Related meq Genovariation in Asia between 2011 and 2021

 ,
,  ,
,  , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Literature Screening
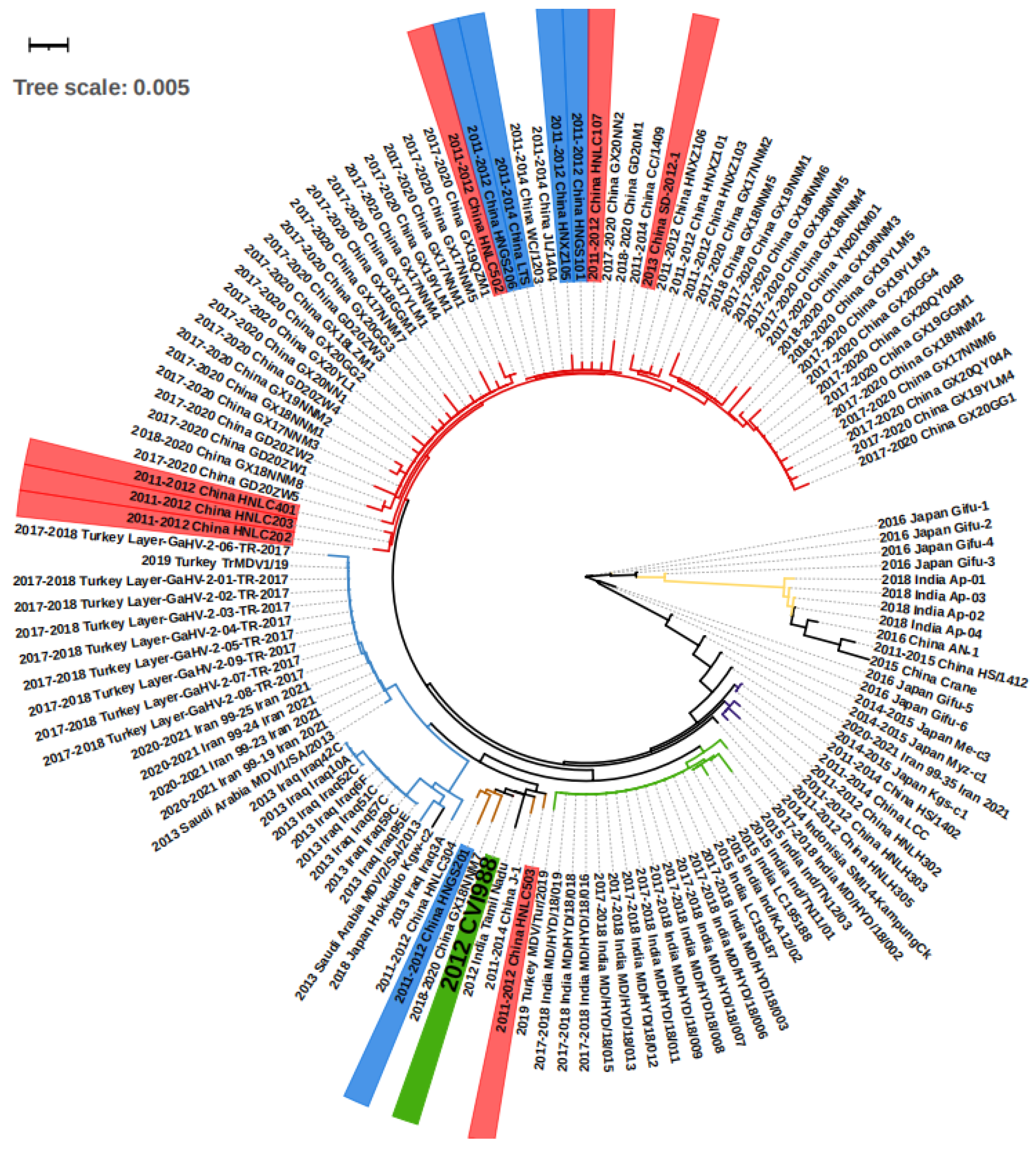
2.2. Phylogenetic Analyses
3. Results and Discussions
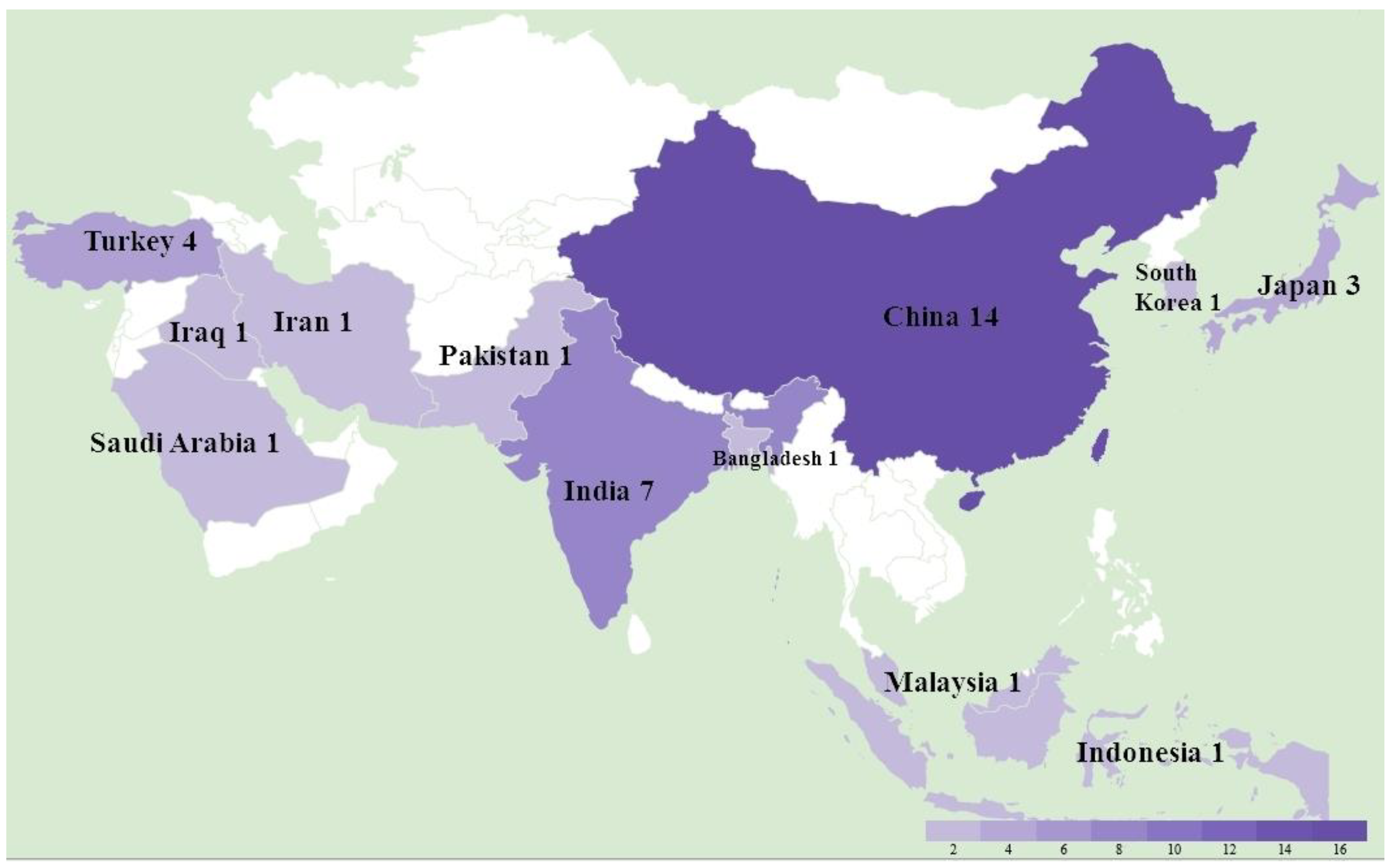
3.1. Overall MD Outbreaks in Asia between 2011 and 2021 and Associated Phylogeny Based on meq Gene
3.2. MD in Specific Asian Countries
3.2.1. China
3.2.2. India
3.2.3. Japan
3.2.4. Turkey
3.2.5. Other Regions
3.3. Amino Acid Changes of Virulent Strains
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kennedy, D.A.; Cairns, C.; Jones, M.J.; Bell, A.S.; Salathé, R.M.; Baigent, S.J.; Nair, V.K.; Dunn, P.A.; Read, A.F. Industry-Wide Surveillance of Marek’s Disease Virus on Commercial Poultry Farms. Avian Dis. 2017, 61, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Biggs, P.M.; Nair, V. The long view: 40 years of Marek’s disease research and Avian Pathology. Avian Pathol. 2012, 41, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, R.; Singh, A.; Singh, P.K.; Teja, E.S.; Varshney, R. Dynamics of Marek’s disease in poultry industry. Pharma Innov. 2021, 10, 80–83. [Google Scholar] [CrossRef]
- Baigent, S.J.; Davison, F. 6-Marek’s disease virus: Biology and life cycle. In Marek’s Disease; Davison, F., Nair, V., Eds.; Academic Press: Oxford, UK, 2004; pp. 62–77. [Google Scholar] [CrossRef]
- Najem, H.A. Clinical, Pathological and Molecular Study of Marek’s Disease in the Gallus gllus Domesticus and Brahma Chickens of Basra Province, Iraq. Egypt. J. Vet. Sci. 2021, 52, 63–71. [Google Scholar] [CrossRef]
- Das, S.; Das, D.; Panda, S.K.; Sagarika, S.; Jena, B. Clinico-Pathological Studies of Marek’s Disease in Chickens. Int. J. Livest. Res. 2018, 8, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Zhang, Y.; Sun, G.; Zhang, F.; Yu, Z.; Gao, Y.; Qi, X.; Li, K.; Wang, X.; Liu, C. Study on the injury of chicken nervous system caused by Marek’s disease virus infection. Zhongguo Yufang Shouyi Xuebao/Chin. J. Prev. Vet. Med. 2019, 41, 113–117. [Google Scholar]
- Osterrieder, N.; Kamil, J.P.; Schumacher, D.; Tischer, B.K.; Trapp, S. Marek’s disease virus: From miasma to model. Nat. Rev. Microbiol. 2006, 4, 283–294. [Google Scholar] [CrossRef]
- McPherson, M.C.; Delany, M.E. Virus and host genomic, molecular, and cellular interactions during Marek’s disease pathogenesis and oncogenesis. Poult. Sci. 2016, 95, 412–429. [Google Scholar] [CrossRef]
- Lupiani, B.; Lee, L.F.; Cui, X.; Gimeno, I.; Anderson, A.; Morgan, R.W.; Silva, R.F.; Witter, R.L.; Kung, H.-J.; Reddy, S.M. Marek’s disease virus-encoded Meq gene is involved in transformation of lymphocytes but is dispensable for replication. Proc. Natl. Acad. Sci. USA 2004, 101, 11815–11820. [Google Scholar] [CrossRef] [Green Version]
- Suchodolski, P.F.; Izumiya, Y.; Lupiani, B.; Ajithdoss, D.K.; Lee, L.F.; Kung, H.-J.; Reddy, S.M. Both homo and heterodimers of Marek’s disease virus encoded Meq protein contribute to transformation of lymphocytes in chickens. Virology 2010, 399, 312–321. [Google Scholar] [CrossRef] [Green Version]
- Burgess, S.C.; Young, J.R.; Baaten, B.J.G.; Hunt, L.; Ross, L.N.J.; Parcells, M.S.; Kumar, P.M.; Tregaskes, C.A.; Lee, L.F.; Davison, T.F. Marek’s disease is a natural model for lymphomas overexpressing Hodgkin’s disease antigen (CD30). Proc. Natl. Acad. Sci. USA 2004, 101, 13879–13884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Lian, L.; Yang, N.; Qu, L. Temporal expression and DNA hypomethylation profile of CD30 in Marek’s disease virus-infected chicken spleens. Poult. Sci. 2015, 94, 1165–1169. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-L.; Ye, Y.; Qian, Z.; Qian, Y.; Templeton, D.J.; Lee, L.F.; Kung, H.-J. Functional Interactions between Herpesvirus Oncoprotein MEQ and Cell Cycle Regulator CDK2. J. Virol. 1999, 73, 4208–4219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.C.; Baigent, S.J.; Smith, L.P.; Chattoo, J.P.; Petherbridge, L.J.; Hawes, P.; Allday, M.J.; Nair, V. Interaction of MEQ protein and C-terminal-binding protein is critical for induction of lymphomas by Marek’s disease virus. Proc. Natl. Acad. Sci. USA 2006, 103, 1687–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, J.-Q.; Teng, M.; Yu, Z.-H.; Xu, H.; Su, J.-W.; Zhao, P.; Xing, G.-X.; Liang, H.-D.; Deng, R.-G.; Qu, L.-H.; et al. Marek’s disease virus-encoded analog of microRNA-155 activates the oncogene c-Myc by targeting LTBP1 and suppressing the TGF-β signaling pathway. Virology 2015, 476, 72–84. [Google Scholar] [CrossRef] [Green Version]
- Bertzbach, L.D.; Kheimar, A.; Ali, F.A.Z.; Kaufer, B.B. Viral Factors Involved in Marek’s Disease Virus (MDV) Pathogenesis. Curr. Clin. Microbiol. Rep. 2018, 5, 238–244. [Google Scholar] [CrossRef]
- Dunn, J.R.; Gimeno, I.M. Current Status of Marek’s Disease in the United States and Worldwide Based on a Questionnaire Survey. Avian Dis. 2013, 57, 483–490. [Google Scholar] [CrossRef]
- Gimeno, I.M. Marek’s disease vaccines: A solution for today but a worry for tomorrow? Vaccine 2008, 26, C31–C41. [Google Scholar] [CrossRef]
- Morrow, C.; Fehler, F. 5-Marek’s disease: A worldwide problem. In Marek’s Disease; Davison, F., Nair, V., Eds.; Academic Press: Oxford, UK, 2004; pp. 49–61. [Google Scholar] [CrossRef]
- Lv, H.; Zhang, Y.; Sun, G.; Bao, K.; Gao, Y.; Qi, X.; Cui, H.; Wang, Y.; Li, K.; Gao, L.; et al. Genetic evolution of Gallid herpesvirus 2 isolated in China. Infect. Genet. Evol. 2017, 51, 263–274. [Google Scholar] [CrossRef]
- Yanamala, P.; Sreedevi, B.; Nagaram, V.K.; Chintamaneni, S. Isolation and molecular characterization of a virulent Marek’s Disease virus serotype-1 from Andhra Pradesh, India. Vet. Ital. 2021, 57, 29–39. [Google Scholar] [CrossRef]
- Kannaki, T.R.; Priyanka, E.; Nishitha, Y.; Krishna, S.V.; Haunshi, S.; Subbiah, M. Molecular detection and phylogenetic analysis of Marek’s disease virus virulence-associated genes from vaccinated flocks in southern India reveals circulation of virulent MDV genotype. Transbound. Emerg. Dis. 2021, 68, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Abd-Ellatieff, H.A.; Abou Rawash, A.A.; Ellakany, H.F.; Goda, W.M.; Suzuki, T.; Yanai, T. Molecular characterization and phylogenetic analysis of a virulent Marek’s disease virus field strain in broiler chickens in Japan. Avian Pathol. 2018, 47, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, A.; Turan, N.; Bayraktar, E.; Tali, H.E.; Aydin, O.; Umar, S.; Cakan, B.; Sadeyen, J.-R.; Baigent, S.; Iqbal, M.; et al. Molecular characterisation and phylogenetic analysis of Marek’s disease virus in Turkish layer chickens. Br. Poult. Sci. 2020, 61, 523–530. [Google Scholar] [CrossRef]
- Prathibha, Y.; Sreedevi, B.; Kumar, N.V.; Srilatha, C.H. Molecular characterization and phylogenetic analysis of oncogenes from virulent serotype-1 Marek’s disease virus in India. Acta Virol. 2018, 62, 277–286. [Google Scholar] [CrossRef]
- Wajid, S.J.; Katz, M.E.; Renz, K.G.; Walkden-Brown, S.W. Prevalence of Marek’s Disease Virus in Different Chicken Populations in Iraq and Indicative Virulence Based on Sequence Variation in the EcoRI-Q (meq) Gene. Avian Dis. 2013, 57, 562–568. [Google Scholar] [CrossRef]
- Machida, Y.; Murata, S.; Matsuyama-Kato, A.; Isezaki, M.; Taneno, A.; Sakai, E.; Konnai, S.; Ohashi, K. Isolation and purification of Gallid herpesvirus 2 strains currently distributed in Japan. J. Vet. Med. Sci. 2017, 79, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Gopal, S.; Manoharan, P.; Kathaperumal, K.; Chidambaram, B.; Divya, K.C. Differential Detection of Avian Oncogenic Viruses in Poultry Layer Farms and Turkeys by Use of Multiplex PCR. J. Clin. Microbiol. 2012, 50, 2668–2673. [Google Scholar] [CrossRef] [Green Version]
- Gong, Z.; Zhang, L.; Wang, J.; Chen, L.; Shan, H.; Wang, Z.; Ma, H. Isolation and analysis of a very virulent Marek’s disease virus strain in China. Virol. J. 2013, 10, 155. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.-H.; Teng, M.; Luo, J.; Wang, X.-W.; Ding, K.; Yu, L.-L.; Su, J.-W.; Chi, J.-Q.; Zhao, P.; Hu, B.; et al. Molecular characteristics and evolutionary analysis of field Marek’s disease virus prevalent in vaccinated chicken flocks in recent years in China. Virus Genes 2013, 47, 282–291. [Google Scholar] [CrossRef]
- Muniyellappa, H.K.; Satyanarayana, M.L.; Isloor, S.; Gowda, N.K.S. Marek’s disease outbreak among vaccinated commercial layer flocks in the mining area of Karnataka, India. Vet. Rec. 2013, 172, 452. [Google Scholar] [CrossRef]
- Rashid, H.; Xue, C.; Islam, R.; Islam, T.; Cao, Y. A longitudinal study on the incidence of mortality of infectious diseases of commercial layer birds in Bangladesh. Prev. Vet. Med. 2013, 109, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-P.; Li, Z.-J.; Bao, K.-Y.; Lv, H.-C.; Gao, Y.-L.; Gao, H.-L.; Qi, X.-L.; Cui, H.-Y.; Wang, Y.-Q.; Ren, X.-G.; et al. Pathogenic characteristics of Marek’s disease virus field strains prevalent in China and the effectiveness of existing vaccines against them. Vet. Microbiol. 2015, 177, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Zou, H.; Shi, H.; Shao, H.; Ye, J.; Miao, J.; Wu, G.; Qin, A. Outbreak of Marek’s disease in a vaccinated broiler breeding flock during its peak egg-laying period in China. BMC Vet. Res. 2015, 11, 157. [Google Scholar] [CrossRef] [Green Version]
- Suresh, P.; Johnson Rajeswar, J.; Sukumar, K.; Harikrishnan, T.J.; Srinivasan, P. Pathotyping of recent Indian field isolates of Marek’s disease virus serotype 1. Acta Virol. 2015, 59, 156–165. [Google Scholar] [CrossRef] [Green Version]
- Cui, N.; Su, S.; Sun, P.; Zhang, Y.; Han, N.; Cui, Z. Isolation and pathogenic analysis of virulent Marek’s disease virus field strain in China. Poult. Sci. 2016, 95, 1521–1528. [Google Scholar] [CrossRef]
- Hartawan, R.; Dharmayanti, N.L.P.I. The Meq Gene Molecular Profile of Marek’s Disease Virus Serotype 1 From Kampung and Arabic Chicken Farms in Sukabumi, West Java, Indonesia. HAYATI J. Biosci. 2016, 23, 160–167. [Google Scholar] [CrossRef]
- Mohamed, M.H.A.; El-Sabagh, I.M.; Al-Habeeb, M.A.; Al-Hammady, Y.M. Diversity of Meq gene from clinical Marek’s disease virus infection in Saudi Arabia. Vet. World 2016, 9, 572–578. [Google Scholar] [CrossRef]
- Sun, G.-R.; Zhang, Y.-P.; Lv, H.-C.; Zhou, L.-Y.; Cui, H.-Y.; Gao, Y.-L.; Qi, X.-L.; Wang, Y.-Q.; Li, K.; Gao, L.; et al. A Chinese Variant Marek’s Disease Virus Strain with Divergence between Virulence and Vaccine Resistance. Viruses 2017, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Yavuz, O.; Erer, H. Immunohistochemical and immunocytochemical findings associated with Marek’s disease virus in naturally infected laying hens. Biotech. Histochem. 2017, 92, 498–505. [Google Scholar] [CrossRef]
- Lian, X.; Ming, X.; Xu, J.; Cheng, W.; Zhang, X.; Chen, H.; Ding, C.; Jung, Y.-S.; Qian, Y. First molecular detection and characterization of Marek’s disease virus in red-crowned cranes (Grus japonensis): A case report. BMC Vet. Res. 2018, 14, 122. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.; Huang, Q.; Yang, C.; Pan, L.; Wang, G.; Qi, K.; Liu, H. Characterizing the histopathology of natural co-infection with Marek’s disease virus and subgroup J avian leucosis virus in egg-laying hens. Avian Pathol. 2018, 47, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Puro, K.-U.; Bhattacharjee, U.; Baruah, S.; Sen, A.; Das, S.; Ghatak, S.; Doley, S.; Sanjukta, R.; Shakuntala, I. Characterization of Marek’s disease virus and phylogenetic analyses of meq gene from an outbreak in poultry in Meghalaya of Northeast India. VirusDisease 2018, 29, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, Z.; Lan, X.; Zhang, F.; Wang, Q.; Li, K.; Pan, Q.; Gao, Y.; Qi, X.; Cui, H.-Y.; et al. A high frequency of Gallid herpesvirus-2 co-infection with Reticuloendotheliosis virusis associated with high tumor rates in Chinese chicken farms. Vet. Microbiol. 2019, 237, 108418. [Google Scholar] [CrossRef] [PubMed]
- Othman, I.; Aklilu, E. Marek’s disease herpesvirus serotype 1 in broiler breeder and layer chickens in Malaysia. Vet. World 2019, 12, 472–476. [Google Scholar] [CrossRef]
- Oh, S.-I.; Do, Y.J.; Kim, E.; Yi, S.W.; Yoo, J.G. Prevalence of poultry red mite (Dermanyssus gallinae) in Korean layer farms and the presence of avian pathogens in the mite. Exp. Appl. Acarol. 2020, 81, 223–238. [Google Scholar] [CrossRef]
- Deng, Q.; Shi, M.; Li, Q.; Wang, P.; Li, M.; Wang, W.; Gao, Y.; Li, H.; Lin, L.; Huang, T.; et al. Analysis of the evolution and transmission dynamics of the field MDV in China during the years 1995–2020, indicating the emergence of a unique cluster with the molecular characteristics of vv+ MDV that has become endemic in southern China. Transbound. Emerg. Dis. 2021, 68, 3574–3587. [Google Scholar] [CrossRef]
- Shi, M.; Li, M.; Wang, P.; Wang, W.; Li, H.; Gao, Y.; Lin, L.; Huang, T.; Wei, P. An outbreak in three-yellow chickens with clinical tumors of high mortality caused by the coinfection of reticuloendotheliosis virus and Marek’s disease virus: A speculated reticuloendotheliosis virus contamination plays an important role in the case. Poult. Sci. 2021, 100, 19–25. [Google Scholar] [CrossRef]
- Li, M.; Wang, P.; Li, Q.; Deng, Q.; Shi, M.; Mo, M.; Wei, T.; Huang, T.; Wei, P. Reemergence of reticuloendotheliosis virus and Marek’s disease virus co-infection in Yellow-Chickens in Southern China. Poult. Sci. 2021, 100, 101099. [Google Scholar] [CrossRef]
- Murata, S.; Yamamoto, E.; Sakashita, N.; Maekawa, N.; Okagawa, T.; Konnai, S.; Ohashi, K. Research Note: Characterization of S-Meq containing the deletion in Meq protein’s transactivation domain in a Marek’s disease virus strain in Japan. Poult. Sci. 2021, 100, 101461. [Google Scholar] [CrossRef]
- Abayli, H.; Karabulut, B.; Ozbek, R.; Ongor, H.; Timurkaan, N.; Tonbak, S. Detection and molecular characterization of a highly oncogenic Marek’s disease virus from vaccinated hens in Turkey. Acta Virol. 2021, 65, 212–220. [Google Scholar] [CrossRef]
- Ozan, E.; Muftuoglu, B.; Sahindokuyucu, I.; Kurucay, H.N.; Inal, S.; Kuruca, N.; Elhag, A.E.; Karaca, E.; Tamer, C.; Gumusova, S.; et al. Marek’s disease virus in vaccinated poultry flocks in Turkey: Its first isolation with molecular characterization. Arch. Virol. 2021, 166, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Khatoon, N.; Akram, M.; Nisa, N.; Waheed, S. Histopathological study of spleen infected by Marek’s Disease Virus (MDV) in broiler chicken. Ceylon J. Sci. 2021, 50, 97–100. [Google Scholar] [CrossRef]
- Molouki, A.; Ghalyanchilangeroudi, A.; Abdoshah, M.; Shoushtari, A.; Abtin, A.; Eshtartabadi, F.; Mahmoudzadeh Akhijahani, M.; Ziafatikafi, Z.; Babaeimarzango, S.S.; Allahyari, E.; et al. Report of a new meq gene size: The first study on genetic characterisation of Marek’s disease viruses circulating in Iranian commercial layer and backyard chicken. Br. Poult. Sci. 2021, 20, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Lupiani, B.; Izumiya, Y.; Reddy, S.M. Marek’s disease virus Meq oncoprotein interacts with chicken HDAC 1 and 2 and mediates their degradation via proteasome dependent pathway. Sci. Rep. 2021, 11, 637. [Google Scholar] [CrossRef] [PubMed]
- Shamblin, C.E.; Greene, N.; Arumugaswami, V.; Dienglewicz, R.L.; Parcells, M.S. Comparative analysis of Marek’s disease virus (MDV) glycoprotein-, lytic antigen pp38- and transformation antigen Meq-encoding genes: Association of meq mutations with MDVs of high virulence. Vet. Microbiol. 2004, 102, 147–167. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Zhao, Y.; Lin, Y.; Zou, N.; Liu, C.; Liu, P.; Cao, S.; Wen, X.; Huang, Y. Comparative analysis of oncogenic genes revealed unique evolutionary features of field Marek’s disease virus prevalent in recent years in China. Virol. J. 2011, 8, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, X.; Li, X.; Shen, Y.; Qiu, Y.; Shi, Z.; Shao, D.; Jin, Y.; Chen, H.; Ding, C.; Li, L.; et al. The Meq oncoprotein of Marek’s disease virus interacts with p53 and inhibits its transcriptional and apoptotic activities. Virol. J. 2010, 7, 348. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
| Year of Study | Outbreak Year | Region | Number of Chicken Farms | MD Incidence Rate in Farm | Mortality Rate in Farm | Vaccine | Pathology Types | Number of Strains | Method | References |
|---|---|---|---|---|---|---|---|---|---|---|
| 2012 | * | India, | * | * | * | * | Visceral | 1 | PCR, pathology | [29] |
| 2013 | 2013 | China | 1 | * | 38.30% | FC126 | Visceral | 1 | PCR, pathology | [30] |
| 2013 | 2011–2012 | China | 18 | * | * | * | * | 17 | PCR | [31] |
| 2013 | * | India | 12 | * | * | unknown vaccine | Visceral | * | Pathology | [32] |
| 2013 | 2011 | Bangladesh | * | * | * | * | Visceral | * | Pathology | [33] |
| 2013 | 2013 | Iraq | * | * | 36.8% and 65% | * | * | 9 | PCR | [27] |
| 2015 | 2011 | China | * | * | * | * | * | * | PCR | [34] |
| 2015 | 2014 | China | 1 | 5% | 80% | CVI988/Rispens | Visceral | * | PCR, pathology | [35] |
| 2015 | 2012 | India | 5 | * | * | * | Visceral | * | PCR, pathology | [36] |
| 2016 | 2013 | China | 1 | * | 22.30% | CVI988/Rispens | * | * | PCR | [37] |
| 2016 | 2014 | Indonesia | 38.8%, 35%, and 20.8% | * | * | * | 1 | PCR | [38] | |
| 2016 | 2016 | Saudi Arabia | * | * | * | CVI988, HVT and FC126 | 2 | PCR | [39] | |
| 2017 | 2011–2014 | China | * | 20–40% | * | CVI988/Rispens | * | 6 | PCR | [21] |
| 2017 | 2015 | China | 1 | 36% | * | CVI988/Rispens | * | 1 | PCR | [40] |
| 2017 | 2014 | Japan | * | * | * | * | * | 3 | PCR, pathology | [28] |
| 2017 | * | Turkey | 11 | * | * | * | Neural, visceral | * | PCR, pathology | [41] |
| 2018 | 2015 | China | * | * | * | * | Visceral | 1 | PCR, pathology | [42] |
| 2018 | 2016 | China | 1 | 0.10% | * | CVI988/Rispens | 1 | PCR | [43] | |
| 2018 | * | India | * | * | * | * | * | * | PCR | [26] |
| 2018 | 2015 | India | 2 | * | 5.5% and 34% | unknown vaccine | Neural, visceral | 5 | PCR, pathology | [44] |
| 2018 | 2016 | Japan | 6 | * | * | CVI988 and HVT | Neural, visceral | 6 | PCR, pathology | [24] |
| 2019 | 2011–2015 | China | 165 | * | * | * | * | 1 | PCR | [45] |
| 2019 | 2017–2018 | Turkey | 49 | * | * | CVI988 and HVT | * | 9 | PCR | [25] |
| 2019 | 2018 | Malaysia | 4 | * | * | * | Visceral | * | PCR | [46] |
| 2020 | 2018–2019 | South Korea | * | * | * | * | * | * | PCR | [47] |
| 2021 | 2017–2020 | China | * | * | * | * | * | 37 | PCR | [48] |
| 2021 | 2018 | China | 3 | 20% | 10% | CVI988/Rispens | Visceral | 1 | PCR, pathology | [49] |
| 2021 | 2018–2020 | China | * | 5–20% | 2–10% | CVI988/Rispens | * | 5 | PCR | [50] |
| 2021 | 2017–2018 | India | 5 | * | * | * | * | 13 | PCR | [23] |
| 2021 | 2018 | India | 1 | * | * | * | * | 4 | PCR | [22] |
| 2021 | 2018 | Japan | 1 | * | * | * | * | 1 | PCR, pathology | [51] |
| 2021 | 2019 | Turkey | 1 | * | * | * | * | 1 | PCR | [52] |
| 2021 | 2019 | Turkey | 1 | * | 1–2% | CVI988, HVT and FC126 | Visceral | 1 | PCR, pathology | [53] |
| 2021 | 2019 | Pakistan | * | * | * | * | Neural, visceral | * | Pathology | [54] |
| 2021 | 2020–2021 | Iran | * | * | * | * | * | 5 | PCR | [55] |
| References | Region | Species | Liver | Spleen | Bursa of Fabricius | Kidney | Skin | Heart | Gizzard | Intestine | Lung |
|---|---|---|---|---|---|---|---|---|---|---|---|
| [35] | China | layer | y | y | y | * | toe | * | * | * | * |
| [42] | China | Crane | y | * | * | * | leg | y | y | * | * |
| [30] | China | broiler | y | y | * | y | * | y | y | * | * |
| [49] | China | broiler | y | * | * | * | * | y | * | y | * |
| [29] | India | layer | y | * | y | * | * | * | * | * | * |
| [32] | India | layer | * | * | * | * | leg | * | * | * | y |
| [36] | India | layer | y | y | y | * | * | * | * | y | * |
| [44] | India | layer | y | y | * | y | * | * | * | * | y |
| [41] | Turkey | layer | y | y | * | y | * | * | y | * | * |
| [53] | Turkey | broiler | y | y | * | * | * | y | * | * | * |
| [24] | Japan | broiler | y | y | * | y | * | * | y | y | * |
| [33] | Bangladesh | broiler | * | * | y | * | y | * | * | * | * |
| [46] | Malaysia | broiler | y | y | y | * | * | * | * | * | * |
| [54] | Pakistan | broiler | * | y | * | * | * | * | * | * | * |
| Viral Strain | Amino Acid Site | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 27 | 59 | 71 | 80 | 88 | 115 | 133 | 139 | 176 | 217 | 237 | 254 | 277 | 293 | 318 | 320 | 332 | |
| 2012 CVI988 | S | K | S | D | A | V | T | T | P | P | C | I | L | P | I | I | W |
| 2011–2012 China HNXZ105 | . | . | A | Y | . | A | P | A | R | A | . | . | . | . | . | . | . |
| 2011–2012 China HNXZ101 | . | . | A | Y | T | A | . | A | R | A | . | . | . | L | . | . | . |
| 2011–2014 China LTS | . | . | A | Y | . | A | . | A | R | A | . | . | . | . | . | . | . |
| 2011–2012 China HNGS206 | . | . | A | Y | . | A | . | A | R | A | . | . | . | . | . | . | . |
| 2011–2012 China HNGS201 | . | E | . | . | . | . | . | . | . | . | S | V | . | . | . | M | . |
| 2011–2012 China HNLC503 | . | . | . | . | . | . | . | . | . | A | S | . | . | . | . | . | . |
| 2011–2012 China HNLC401 | P | . | A | Y | . | A | . | . | R | A | . | . | . | . | . | . | . |
| 2011–2012 China HNLC502 | . | . | A | Y | . | A | . | A | R | A | . | . | . | . | . | . | . |
| 2011–2012 China HNLC203 | . | . | A | Y | . | A | . | . | R | A | . | . | . | . | . | . | . |
| 2011–2012 China HNLC202 | . | . | A | Y | . | A | I | . | R | A | . | . | . | . | . | . | G |
| 2011–2012 China HNLC107 | . | . | A | Y | . | A | . | A | R | A | . | . | F | . | . | . | . |
| 2013 China SD-2012-1 | . | . | A | Y | T | A | . | . | R | A | . | . | . | . | V | . | . |
| Viral Strain | Amino Acid Site | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 32 | 39 | 69 | 71 | 80 | 88 | 93 | 112 | 115 | 139 | 176 | 217 | 234 | 249 | 340 | |
| 2012 CVI988 | S | K | I | D | S | D | A | Q | S | V | T | P | P | P | P | - |
| 2018 China GX18NNM5 | A | . | . | . | A | Y | T | R | . | A | A | R | A | . | . | . |
| 2018–2020 China GX18NNM8 | A | . | . | . | A | Y | . | . | . | A | A | R | A | . | T | R |
| 2018–2020 China GX19YLM5 | . | . | . | . | A | Y | T | R | . | A | A | R | A | . | . | . |
| 2018–2020 China GX19NNM3 | . | R | . | G | A | Y | T | R | P | A | A | R | A | . | . | . |
| 2018–2020 China GD20M1 | . | . | V | . | A | Y | . | . | . | A | A | R | A | L | . | . |
| 2019 Turkey MDV/Tur/2019 | - | - | . | . | A | Y | T | R | . | L | A | . | . | . | . | . |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, B.; Zeb, J.; Hussain, S.; Aziz, M.U.; Circella, E.; Casalino, G.; Camarda, A.; Yang, G.; Buchon, N.; Sparagano, O. A Review on the Marek’s Disease Outbreak and Its Virulence-Related meq Genovariation in Asia between 2011 and 2021. Animals 2022, 12, 540. https://doi.org/10.3390/ani12050540
Song B, Zeb J, Hussain S, Aziz MU, Circella E, Casalino G, Camarda A, Yang G, Buchon N, Sparagano O. A Review on the Marek’s Disease Outbreak and Its Virulence-Related meq Genovariation in Asia between 2011 and 2021. Animals. 2022; 12(5):540. https://doi.org/10.3390/ani12050540
Chicago/Turabian StyleSong, Baolin, Jehan Zeb, Sabir Hussain, Muhammad Umair Aziz, Elena Circella, Gaia Casalino, Antonio Camarda, Guan Yang, Nicolas Buchon, and Olivier Sparagano. 2022. "A Review on the Marek’s Disease Outbreak and Its Virulence-Related meq Genovariation in Asia between 2011 and 2021" Animals 12, no. 5: 540. https://doi.org/10.3390/ani12050540
APA StyleSong, B., Zeb, J., Hussain, S., Aziz, M. U., Circella, E., Casalino, G., Camarda, A., Yang, G., Buchon, N., & Sparagano, O. (2022). A Review on the Marek’s Disease Outbreak and Its Virulence-Related meq Genovariation in Asia between 2011 and 2021. Animals, 12(5), 540. https://doi.org/10.3390/ani12050540