
Virulence Genes as Markers for Pseudomonas aeruginosa Biofilm Formation in Dogs and Cats



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolates
2.2. DNA Extraction
2.3. PCR Assay
2.4. Microtiter Plate Method
2.5. Statistical Analysis
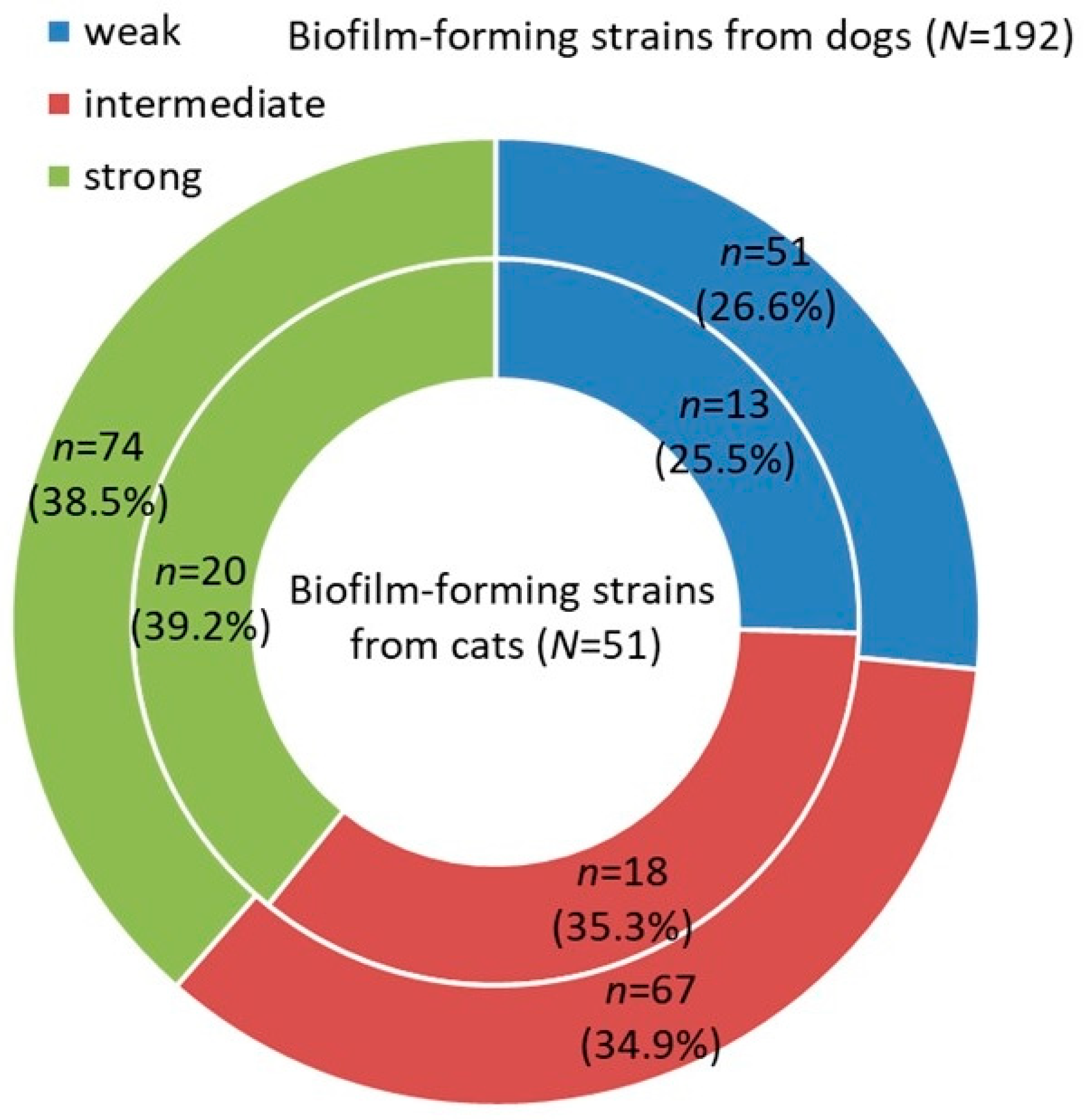
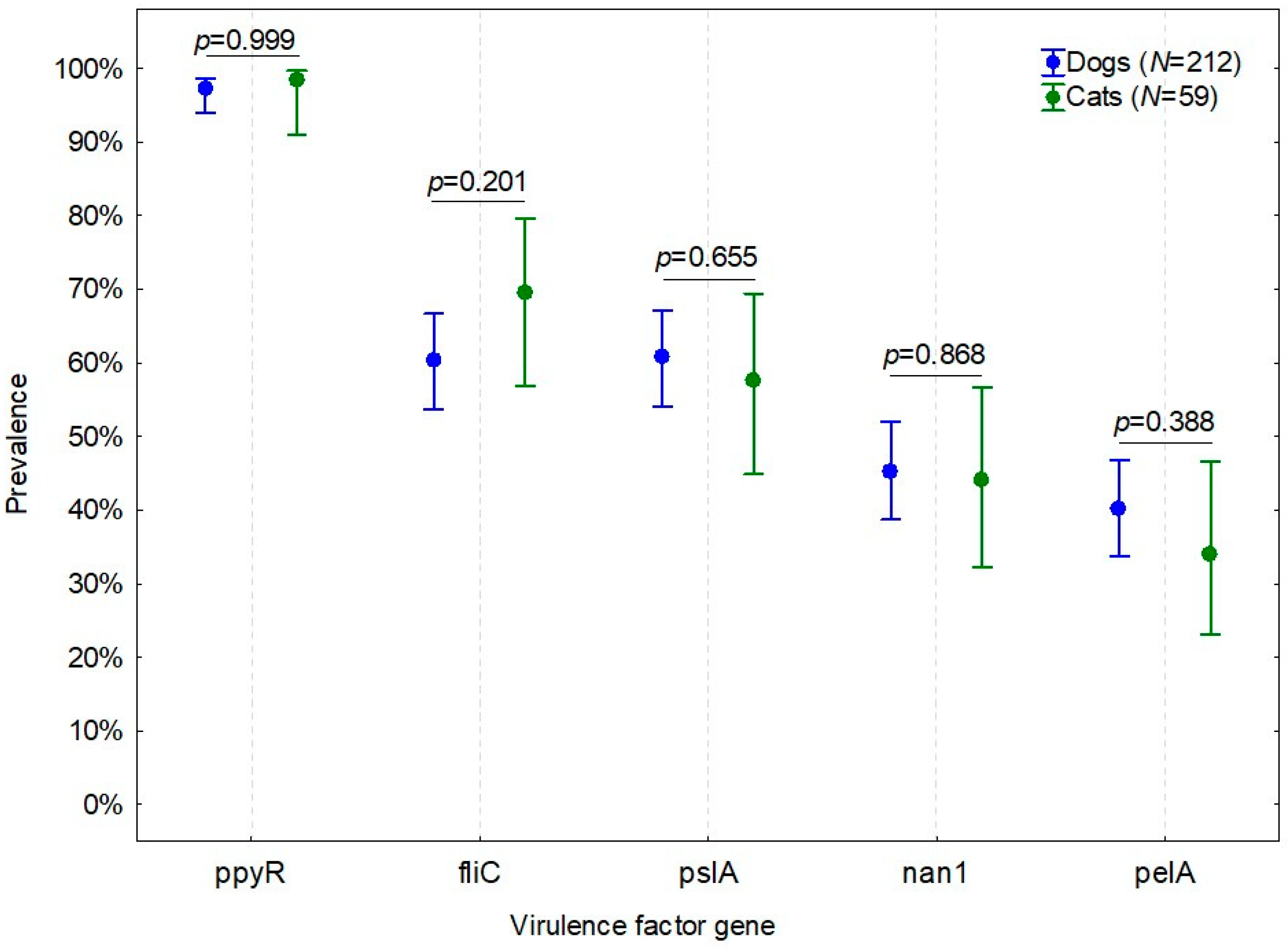
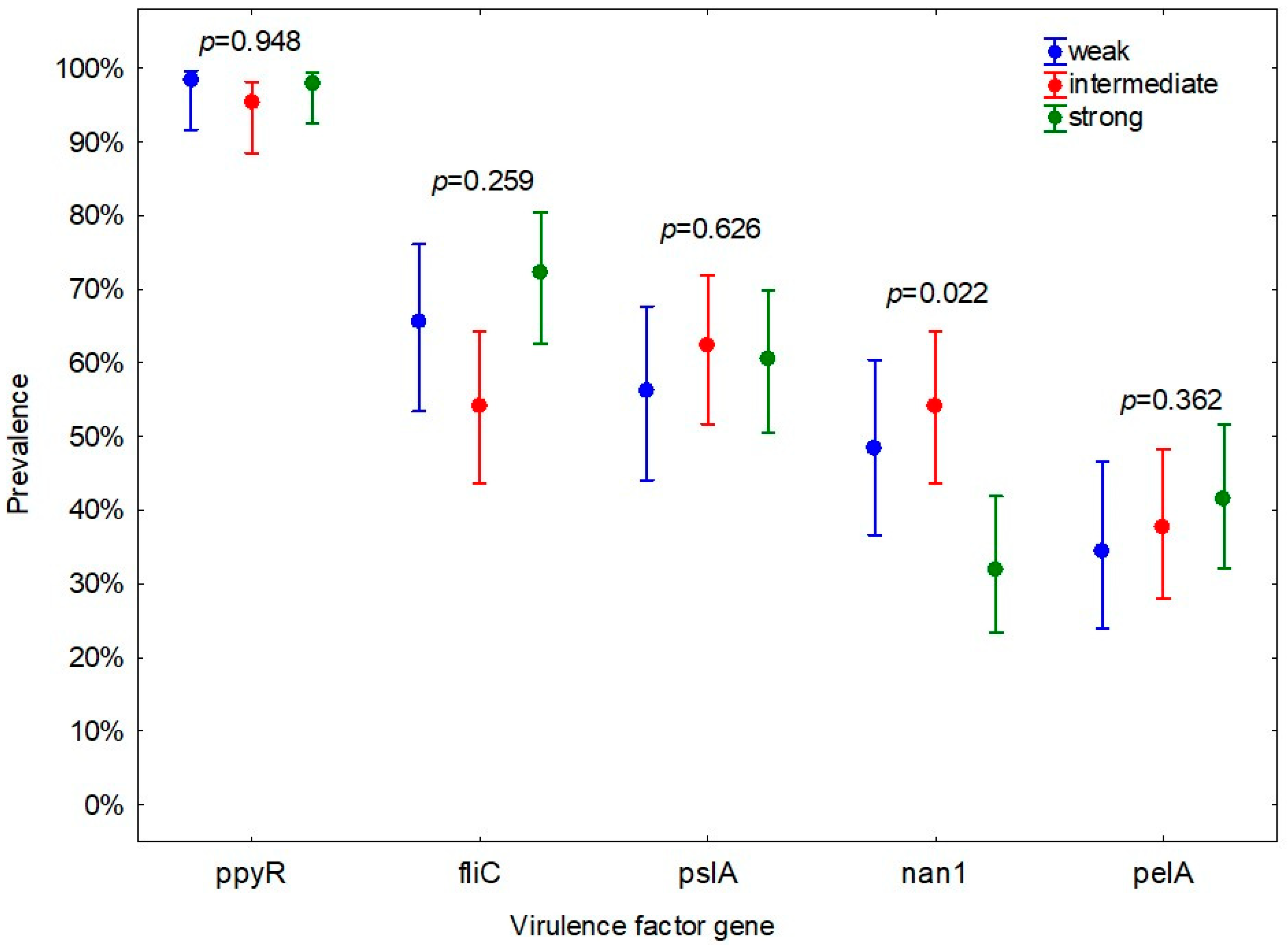
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Da Silva, L.C.A.; Pessoa, D.A.; Maia, L.A.; Matos, R.A.; Macêdo, M.M. Systemic infection by Pseudomonas aeruginosa in a dog. Act. Sci. Vet. 2016, 44, 1–5. [Google Scholar] [CrossRef]
- Hillier, A.; Alcorn, J.R.; Cole, L.K.; Kowalski, J.J. Pyoderma caused by Pseudomonas aeruginosa infection in dogs: 20 cases. Vet. Dermatol. 2006, 17, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Robinson, V.H.; Paterson, S.; Bennett, C.; Steen, S.I. Biofilm production of Pseudomonas spp. isolates from canine otitis in three different enrichment broths. Vet. Dermatol. 2019, 30, 218-e67. [Google Scholar] [CrossRef] [PubMed]
- Prado, M.R.; Rocha, M.F.G.; Brito, É.H.S.; Girão, M.D.; Monteiro, A.J.; Teixeira, M.F.S.; Sidrim, J.J.C. Survey of bacterial microorganisms in the conjunctival sac of clinically normal dogs and dogs with ulcerative keratitis in Fortaleza, Ceará, Brazil. Vet. Ophthalmol. 2005, 8, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Pakravanm, N.; Pritchard, J.C.; Hartmann, F.A.; Young, K.M. Mucoid Pseudomonas aeruginosa infection in a cat with severe chronic rhinosinusitis. Vet. Clin. Pathol. 2019, 48, 300–304. [Google Scholar] [CrossRef]
- Chawnan, N.; Na Lampang, K.; Mektrirat, R.; Awaiwanont, N.; Thongkorn, K. Cultivation of bacterial pathogens and antimicrobial resistance in canine periapical tooth abscesses. Vet. Integr. Sci. 2021, 19, 513–524. [Google Scholar] [CrossRef]
- Riggio, M.P.; Lennon, A.; Taylor, D.J.; Bennett, D. Molecular identification of bacteria associated with canine periodontal disease. Vet. Microbiol. 2011, 3–4, 394–400. [Google Scholar]
- Shehab, Z.; Al-Rubaii, B. Correlation of nan1 (Neuraminidase) and production of some type III secretion system in clinical isolates of Pseudomonas aeruginosa. Biosci. Res. 2018, 15, 1729–1738. [Google Scholar]
- Paterson, S. Biofilms: Their importance in veterinary medicine. Companion Anim. 2017, 22, 659–668. [Google Scholar] [CrossRef]
- Vestby, L.K.; Grønseth, T.; Simm, R.; Nesse, L.L. Bacterial Biofilm and its Role in the Pathogenesis of Disease. Antibiotics 2020, 9, 59. [Google Scholar] [CrossRef]
- Wei, Q.; Ma, L.Z. Biofilm Matrix and Its Regulation in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2013, 14, 20983–21005. [Google Scholar] [CrossRef] [PubMed]
- Jyot, J.; Ramphal, R. Flagella and Pili of Pseudomonas aeruginosa. In Pseudomonas: Model Organism, Pathogen, Cell Factory; Wiley-VCH: Weinheim, Germany, 2008; pp. 85–108. [Google Scholar]
- Mahenthiralingam, E.; Campbell, M.E.; Speert, D.P. Nonmotility and phagocytic resistance of Pseudomonas aeruginosa isolates from chronically colonized patients with cystic fibrosis. Infect. Immun. 1994, 62, 596–605. [Google Scholar] [CrossRef]
- Lanotte, P.; Watt, S.; Mereghetti, L.; Dartiguelongue, N.; Rastegar-Lari, A.; Goudeau, A.; Quentin, R. Genetic features of Pseudomonas aeruginosa isolates from cystic fibrosis patients compared with those of isolates from other origins. J. Med. Microbiol. 2004, 53, 73–81. [Google Scholar] [CrossRef]
- Soong, G.; Muir, A.; Gomez, M.I.; Waks, J.; Reddy, B.; Planet, P.; Singh, P.K.; Kanetko, Y.; Wolfgang, M.C.; Hsiao, Y.-S.; et al. Bacterial neuraminidase facilitates mucosal infection by participating in biofilm production. J. Clin. Investig. 2006, 116, 2297–2305. [Google Scholar] [CrossRef]
- Wolska, K.; Kot, B.; Mioduszewska, H.; Sempruch, C.; Borkowska, L.; Rymuza, K. Occurrence of the nan1 gene and adhesion of Pseudomonas aeruginosa isolates to human buccal epithelial cells. Biol. Lett. 2012, 49, 59–64. [Google Scholar] [CrossRef][Green Version]
- De Vos, D.; Lim, A.; Pirnay, J.P.; Struelens, M.; Vandenvelde, C.; Duinslaeger, L.; Vanderkelen, A.; Cornelis, P. Direct detection and identification of Pseudomonas aeruginosa in clinical samples such as skin biopsy specimens and expectorations by multiplex PCR based on two outer membrane lipoprotein genes, oprI and oprL. J. Clin. Microbiol. 1997, 35, 1295–1299. [Google Scholar] [CrossRef] [PubMed]
- Ghadaksaz, A.; Fooladi, A.A.I.; Hosseini, H.M.; Amin, M. The prevalence of some Pseudomonas virulence genes related to biofilm formation and alginate production among clinical isolates. J. Appl. Biomed. 2015, 13, 61–68. [Google Scholar] [CrossRef]
- Sattari, M.; Godarzi, G.H. The recognizing of fli C gene in Pseudomonas aeruginosa isolated from clinical sample with PCR. Med. Lab. J. 2009, 3. [Google Scholar]
- O’Toole, G.A. Microtiter Dish Biofilm Formation Assay. J. Vis. Exp. 2011, 47, e2437. [Google Scholar] [CrossRef]
- Darwish, S.F.; Asfour, H.A.E. Investigation of biofilm forming ability in staphylococci causing bovine mastitis using phenotypic and genotypic assays. Sci. World J. 2013, 2013, 1–9. [Google Scholar] [CrossRef]
- Agresti, A. An Introduction to Categorical Data Analysis, 2nd ed.; John Wiley & Sons: London, UK, 2007; pp. 41–45. [Google Scholar]
- Newcombe, R.G.; Altman, D.G. Proportions and Their Differences. In Statistics with Confidence: Confidence Intervals and Statistical Guidelines, 2nd ed.; Altman, D.G., Machin, D., Bryant, T.N., Gardner, M.J., Eds.; BMJ Book: London, UK, 2000; pp. 45–48. [Google Scholar]
- Barnard, N.; Foster, A. Pseudomonas otitis in dogs: A general practitioner’s guide to treatment. Practice 2017, 39, 386–398. [Google Scholar] [CrossRef]
- Ceri, H.; Olson, M.; Morck, D.; Storey, D.; Read, R.; Buret, A.; Olson, B. The MBEC assay system: Multiple equivalent biofilms for antibiotic and biocide susceptibility testing. Method. Enzymol. 2001, 337, 377–385. [Google Scholar]
- Milivojevic, D.; Šumonja, N.; Medić, S.; Pavic, A.; Moric, I.; Vasiljevic, B.; Senerovic, L.; Nikodinovic-Runic, J. Biofilm-forming ability and infection potential of Pseudomonas aeruginosa strains isolated from animals and humans. Pathog. Dis. 2018, 76, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Pye, C.C.; Yu, A.A.; Weese, J.S. Evaluation of biofilm production by Pseudomonas aeruginosa from canine ears and the impact of biofilm on antimicrobial susceptibility in vitro. Vet. Dermatol. 2013, 24, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, G.; Dilhari, A.; Gayani, B.; Kottegoda, N.; Samaranayake, L.; Weerasekera, M. Influence of laboratory culture media on in vitro growth, adhesion, and biofilm formation of Pseudomonas aeruginosa and staphylococcus aureus. Med. Princ. Pract. 2018, 28, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Samad, A.; Khan, A.A.; Sajid, M.; Zahra, R. Assessment of biofilm formation by Pseudomonas aeruginosa and hydrodynamic evaluation of microtiter plate assay. J. Pak. Med Assoc. 2019, 69, 666–671. [Google Scholar]
- Ertugrul, B.; Oryasin, E.; Lipsky, B.A.; Willke, A.; Bozdogan, B. Virulence genes fliC, toxA and phzS are common among Pseudomonas aeruginosa isolates from diabetic foot infections. Infect. Dis. 2017, 50, 273–279. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | Overall (n = 271) | Dogs (n = 212) | Cats (n = 59) | p-Value |
|---|---|---|---|---|
| Demographic Characteristics | ||||
| Age a (years) | 7, 4–10 (0.2–18) | 7, 4–10 (0.7–18) | 5, 1–8 (0.2–16) | 0.001 * |
| Male sex (n (%)) | 146 (53.9) | 112 (52.8) | 34 (57.6) | 0.513 |
| Breed (n (%)) | <0.001 * | |||
| Crossbreed/Domestic shorthair | - | 43 (20.3) | 43 (72.9) | |
| Pedigree | - | 169 (79.7) | 16 (27.1) | |
| German Shepherd | - | 33 | - | |
| Cocker spaniel | - | 25 | - | |
| Beagle | - | 12 | - | |
| Golden retriever | - | 10 | - | |
| Shih-tzu | - | 10 | - | |
| Others | - | 79 | - | |
| Collection site | ||||
| External auditory canal | 121 (44.6) | 118 (55.7) | 3 (5.1) | <0.001 * |
| Respiratory system and oral cavity | 85 (31.4) | 36 (17.0) | 49 (83.1) | <0.001 * |
| Nasal cavity | 51 | 7 | 44 | |
| Conjunctival sac | 12 | 10 | 2 | |
| Throat | 10 | 8 | 2 | |
| Trachea and bronchi | 7 | 6 | 1 | |
| Oral cavity | 5 | 5 | 0 | |
| Skin and appendages | 33 (12.2) | 31 (14.6) | 2 (3.4) | 0.020 * |
| Skin | 27 | 25 | 2 | |
| Perianal glands | 6 | 6 | 0 | |
| Wounds | 20 (7.4) | 16 (7.5) | 4 (6.8) | 0.855 |
| Genito-urinary tract | 11 (4.1) | 10 (4.7) | 1 (1.7) | 0.301 |
| Vagina | 7 | 7 | 0 | |
| Urine | 4 | 3 | 1 | |
| Joint fluid | 1 (0.4) | 1 (0.5) | 0 | 0.586 |
| Target Genes | Primer Sequence 5′-3′ | Product Size (bp) | Reference |
|---|---|---|---|
| pslA | F: 5′-TCCCTACCTCAGCAGCAAGC-3′ R: 5′–TGTTGTAGCCGTAGCGTTTCTG-3′ | 656 | Ghadaksaz et al., 2015 [18] |
| pelA | F: 5′-CATACCTTCAGCCATCCGTTCTTC-3′ R: 5′-CGCATTCGCCGCACTCAG-3′ | 786 | Ghadaksaz et al., 2015 [18] |
| ppyR | F: 5′-CGTGATCGCCGCCTATTTCC-3′ R: 5′-ACAGCAGACCTCCCAACCG-3′ | 160 | Ghadaksaz et al., 2015 [18] |
| fliC | F: 5′-TGAACGTGGCTACCAAGAACG-3′ R: 5′-TCTGCAGTTGCTTCACTTCGC-3′ | 180 | Immani et al., 2009 [19] |
| nan1 | F: 5′-ACGCTCCGTCCAGCCGGA-3′ R: 5′-GTCTGGACGACGGCGGCA-3′ | 221 | Lanotte et al., 2004 [14] |
| oprI | F: 5′ATGAACAACGTTCTGAAATTCTCTGCT-3′ R: 5′-CTTGCGGCTGGCTTTTTCCAG-3′ | 249 | De Vos et al., 1997 [17] |
| oprL | F: 5′-ATGGAAATGCTGAAATTCGGC-3′ R: 5′-CTTCTTCAGCTCGACGCGACG-3′ | 504 | De Vos et al., 1997 [17] |
| Collection Site | Biofilm-Forming P. aeruginosa Strains | |
|---|---|---|
| No. of Strains | Prevalence (CI 95%) | |
| Dogs (n = 212) | ||
| External auditory canal (n = 118) | 111 | 94.1 (88.3–97.1) |
| Skin and appendages (n = 31) | 28 | 90.3 (75.1–96.7) |
| Respiratory system and oral cavity (n = 36) | 28 | 77.8 (61.9–88.3) |
| Genito-urinary tract (n = 10) | 10 | 100 (72.2–100) |
| Wounds (n = 16) | 14 | 87.5 (64.0–96.5) |
| Cats (n = 59) | ||
| Respiratory system (n = 49) | 42 | 85.7 (73.3–92.9) |
| Others (n = 10) | 9 | 90.0 (59.6–98.2) |
| Virulence Factor Gene | Pseudomonas aeruginosa Strains | p-Value | |||
|---|---|---|---|---|---|
| Biofilm-Forming | Non-Biofilm-Forming | ||||
| n | Prevalence (CI 95%) | n | Prevalence (CI 95%) | ||
| Dogs | |||||
| pslA | 117 | 60.9 (53.9–67.6) | 12 | 60.0 (38.7–78.1) | 0.935 |
| pelA | 76 | 39.6 (32.9–46.6) | 9 | 45.0 (25.8–65.8) | 0.638 |
| ppyR | 186 | 96.9 (93.4–98.6) | 20 | 100 (83.9–100) | 0.999 |
| fliC | 121 | 63.0 (56.0–69.5) | 7 | 35.0 (18.1–56.7) | 0.015 * |
| nan1 | 88 | 45.8 (38.9–52.9) | 8 | 40.0 (21.9–61.3) | 0.617 |
| Cats | |||||
| pslA | 29 | 56.9 (43.3–69.5) | 5 | 62.5 (30.6–86.3) | 0.999 |
| pelA | 17 | 33.3 (22.0–47.0) | 3 | 37.5 (13.7–69.4) | 0.999 |
| ppyR | 50 | 98.0 (89.7–99.7) | 8 | 100 (67.6–100) | 0.999 |
| fliC | 35 | 68.6 (55.0–79.7) | 6 | 75.0 (40.9–92.9) | 0.999 |
| nan1 | 19 | 37.3 (25.3–51.0) | 7 | 87.5 (52.9–97.8) | 0.017 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Płókarz, D.; Czopowicz, M.; Bierowiec, K.; Rypuła, K. Virulence Genes as Markers for Pseudomonas aeruginosa Biofilm Formation in Dogs and Cats. Animals 2022, 12, 422. https://doi.org/10.3390/ani12040422
Płókarz D, Czopowicz M, Bierowiec K, Rypuła K. Virulence Genes as Markers for Pseudomonas aeruginosa Biofilm Formation in Dogs and Cats. Animals. 2022; 12(4):422. https://doi.org/10.3390/ani12040422
Chicago/Turabian StylePłókarz, Daria, Michał Czopowicz, Karolina Bierowiec, and Krzysztof Rypuła. 2022. "Virulence Genes as Markers for Pseudomonas aeruginosa Biofilm Formation in Dogs and Cats" Animals 12, no. 4: 422. https://doi.org/10.3390/ani12040422
APA StylePłókarz, D., Czopowicz, M., Bierowiec, K., & Rypuła, K. (2022). Virulence Genes as Markers for Pseudomonas aeruginosa Biofilm Formation in Dogs and Cats. Animals, 12(4), 422. https://doi.org/10.3390/ani12040422