


The Photoperiod Regulates Granulosa Cell Apoptosis through the FSH-Nodal/ALK7 Signaling Pathway in Phodopus sungorus

 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
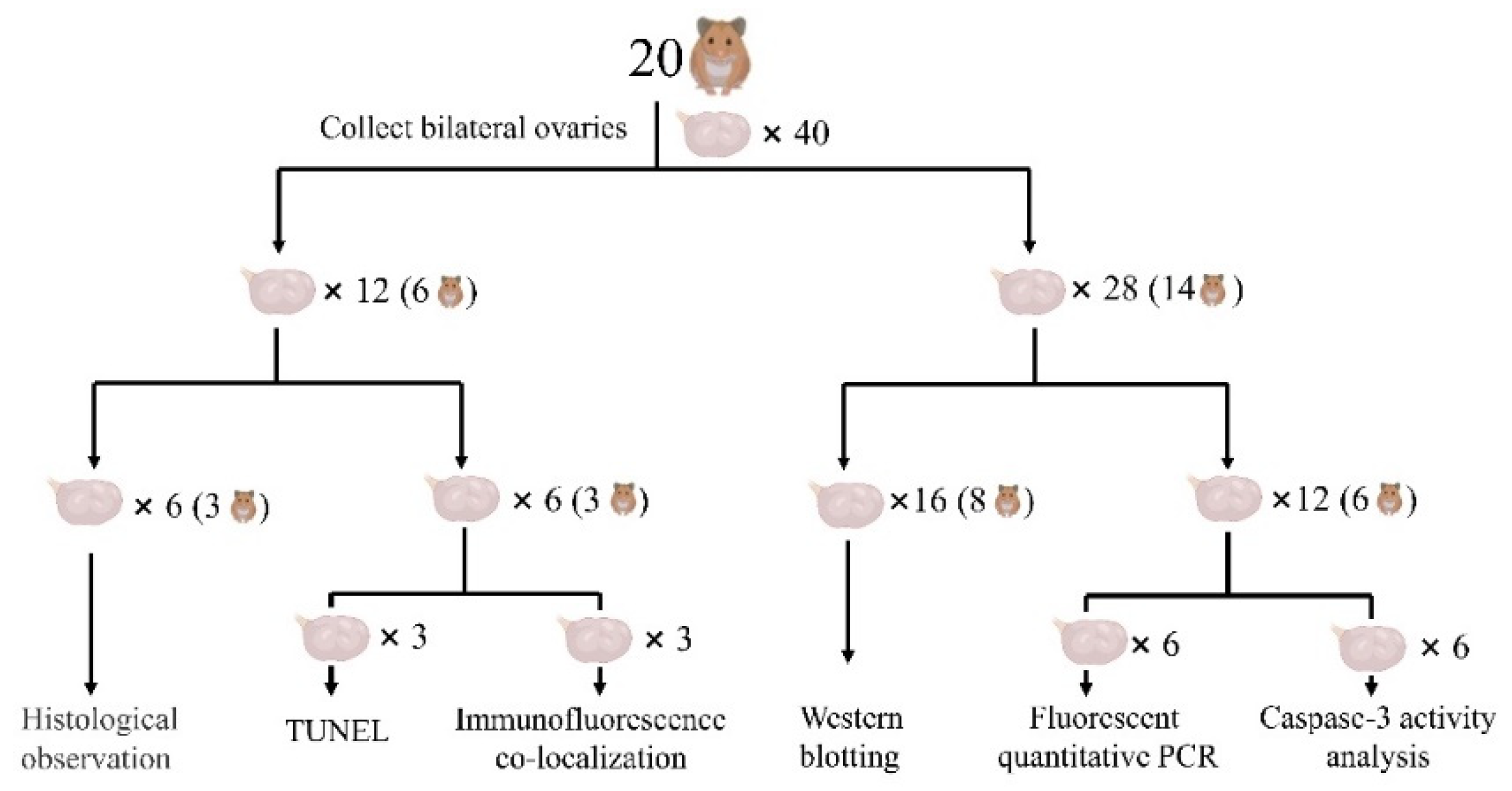
2.1. Sample Collection and Treatments
2.2. Sample Preparation
2.3. Histological Study
2.4. TUNEL Staining
2.5. Immunofluorescence Co-Localization
2.6. Quantitative Real-Time PCR
2.7. Western Blotting
2.8. Detection of the Caspase-3 Enzymatic Activity
2.9. Statistical Analyses
3. Results
3.1. Differences in the Body (BW) and Ovarian Weight (OW) in Hamsters under Different Photoperiods
3.2. Differences in the Serum Hormone Concentrations of the Hamsters under Different Photoperiods
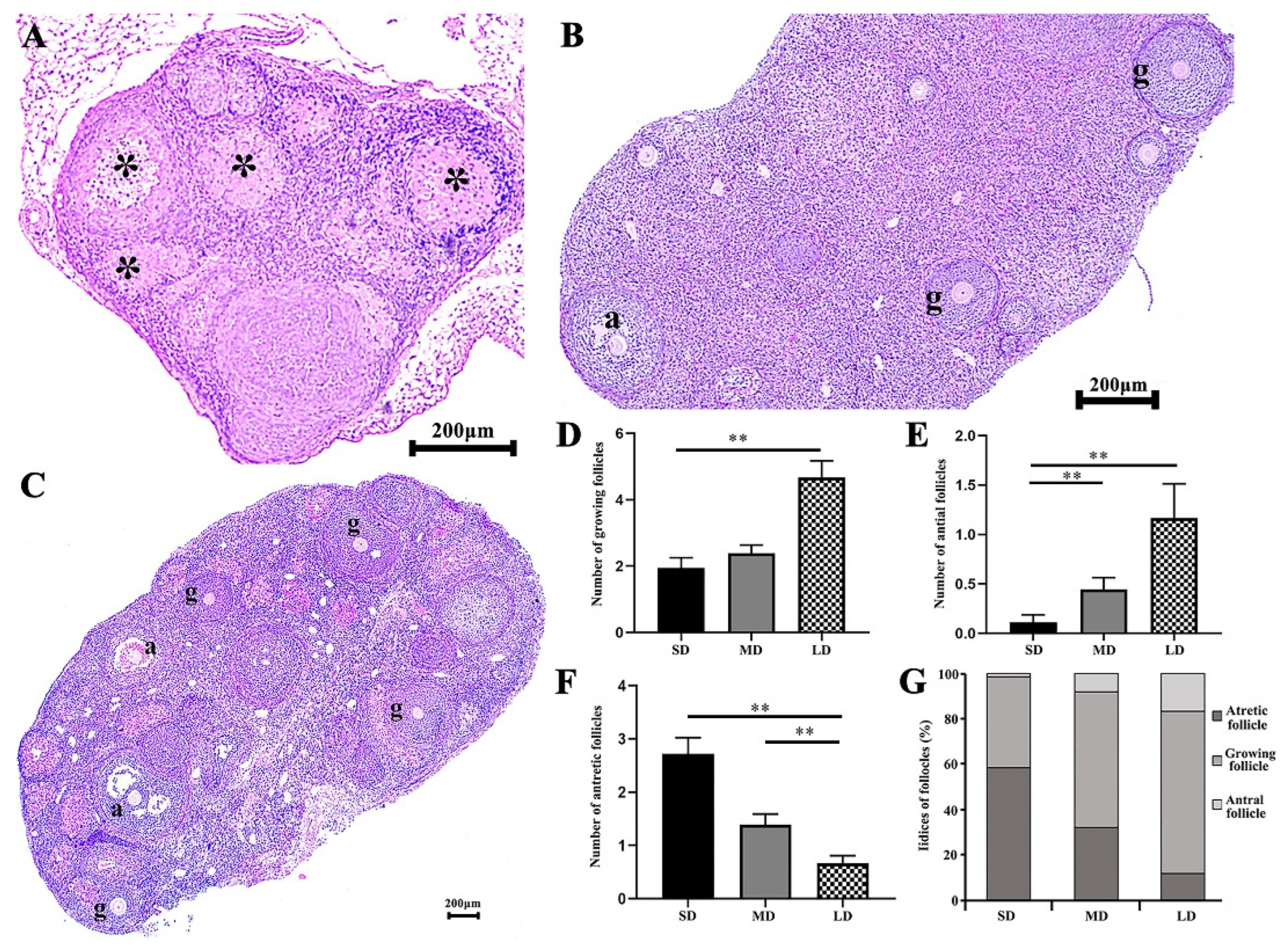
3.3. Differences in the Growing Follicles, Antral and Atretic Follicles in the Hamsters under Different Photoperiods
3.4. Apoptosis of the Granulosa Cells Examined by the TUNEL under Different Photoperiods
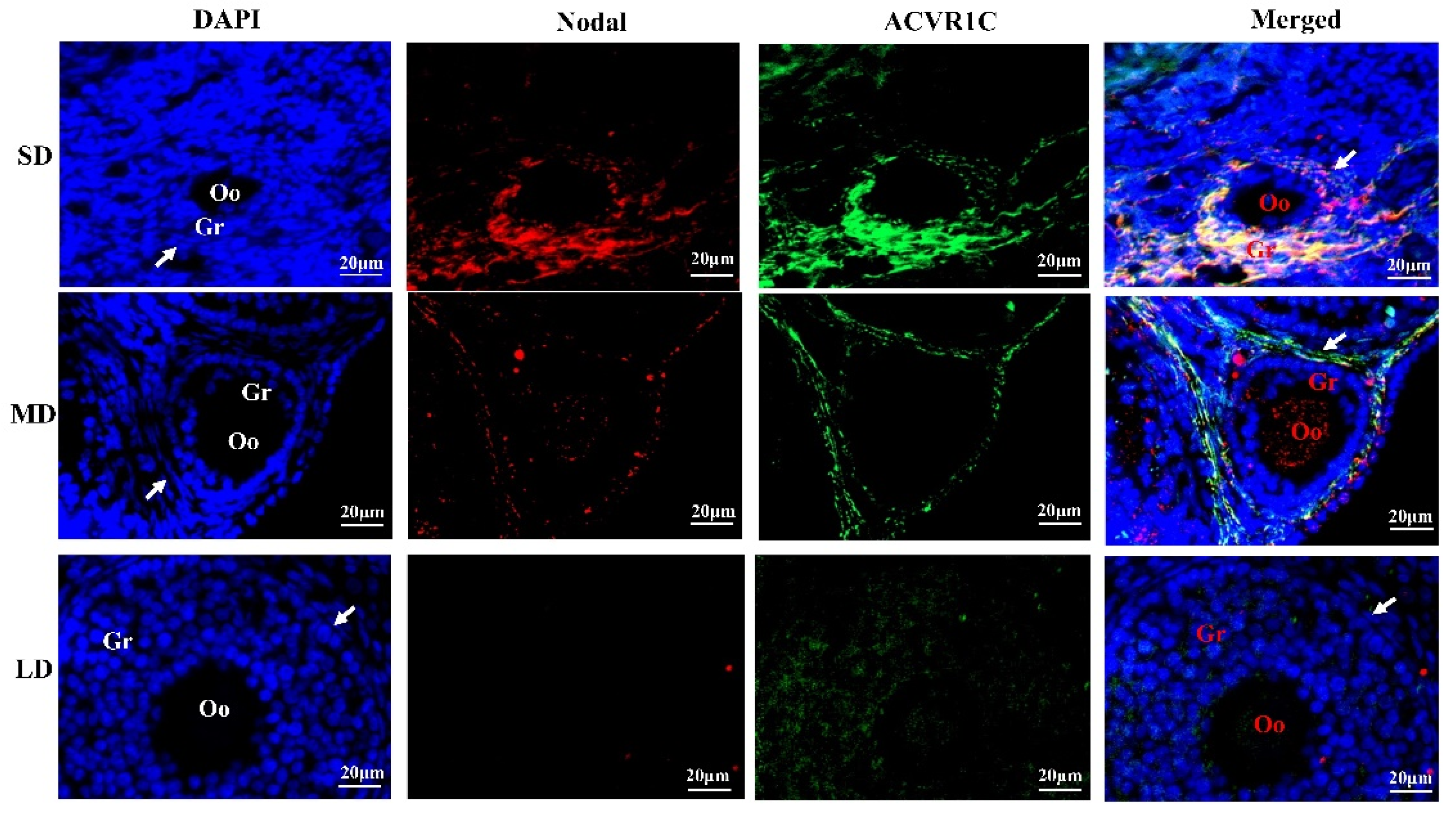
3.5. Interactions between Nodal and ACVR1C under Different Photoperiods
3.6. Differentiated Expression of the Key Genes at the mRNA Level in the Nodal/ALK7 Signaling Pathway
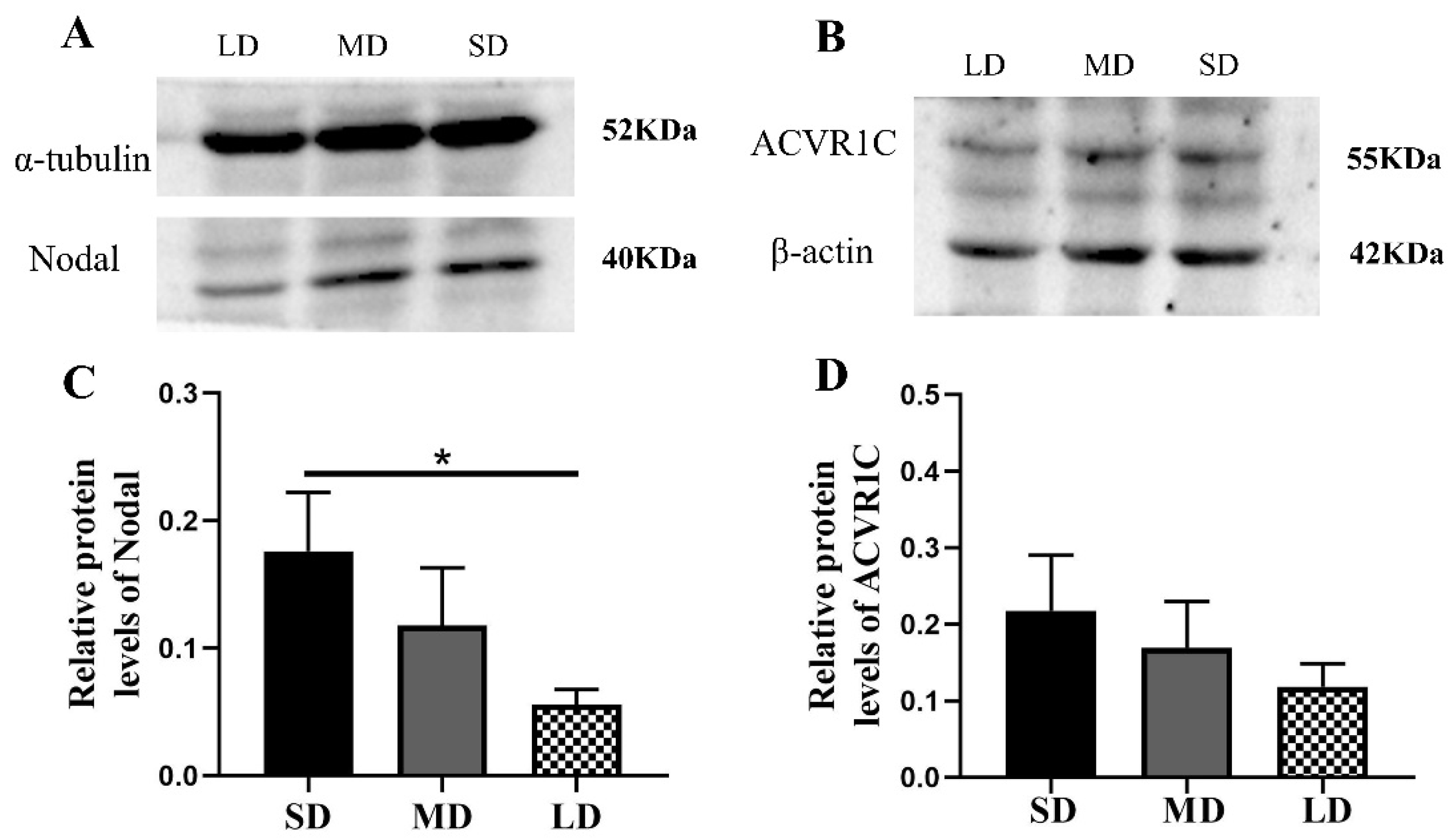
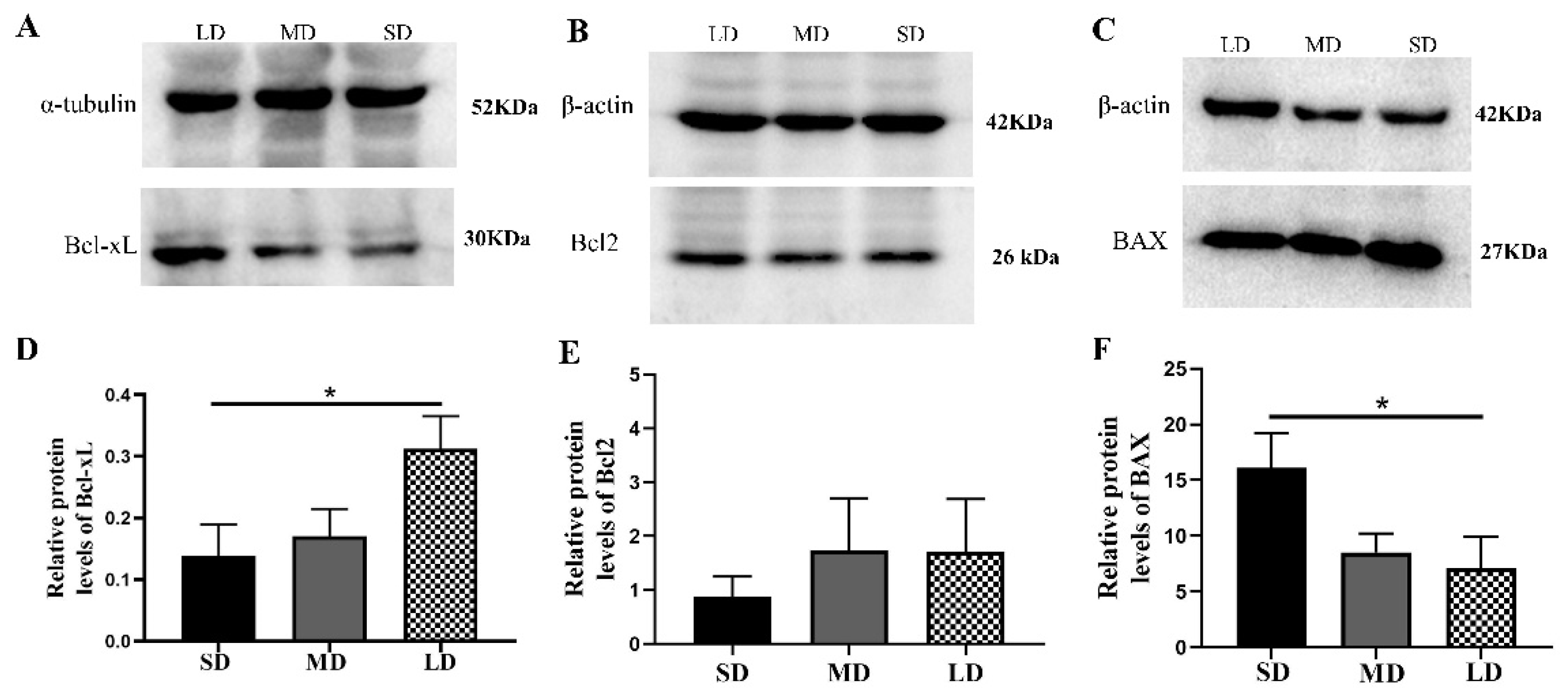
3.7. Differentiated Expression of the Key Genes at the Protein Level in the Nodal/ALK7 Signaling Pathway
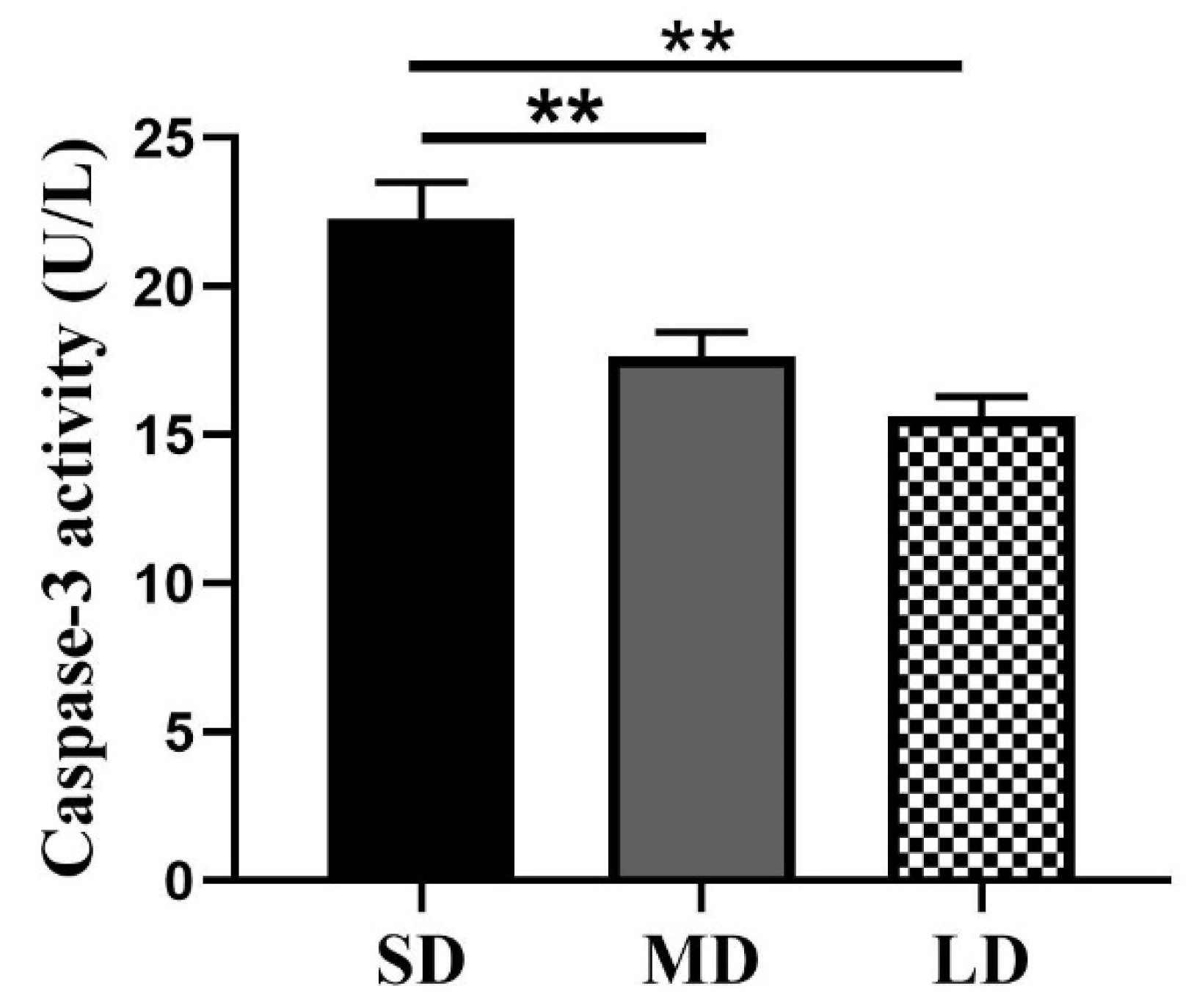
3.8. Differences in the Caspase -3 Enzyme Activity under the Different Photoperiods
4. Discussion
4.1. Hormone Synthesis and Ovarian Morphology Changes with the Photoperiods
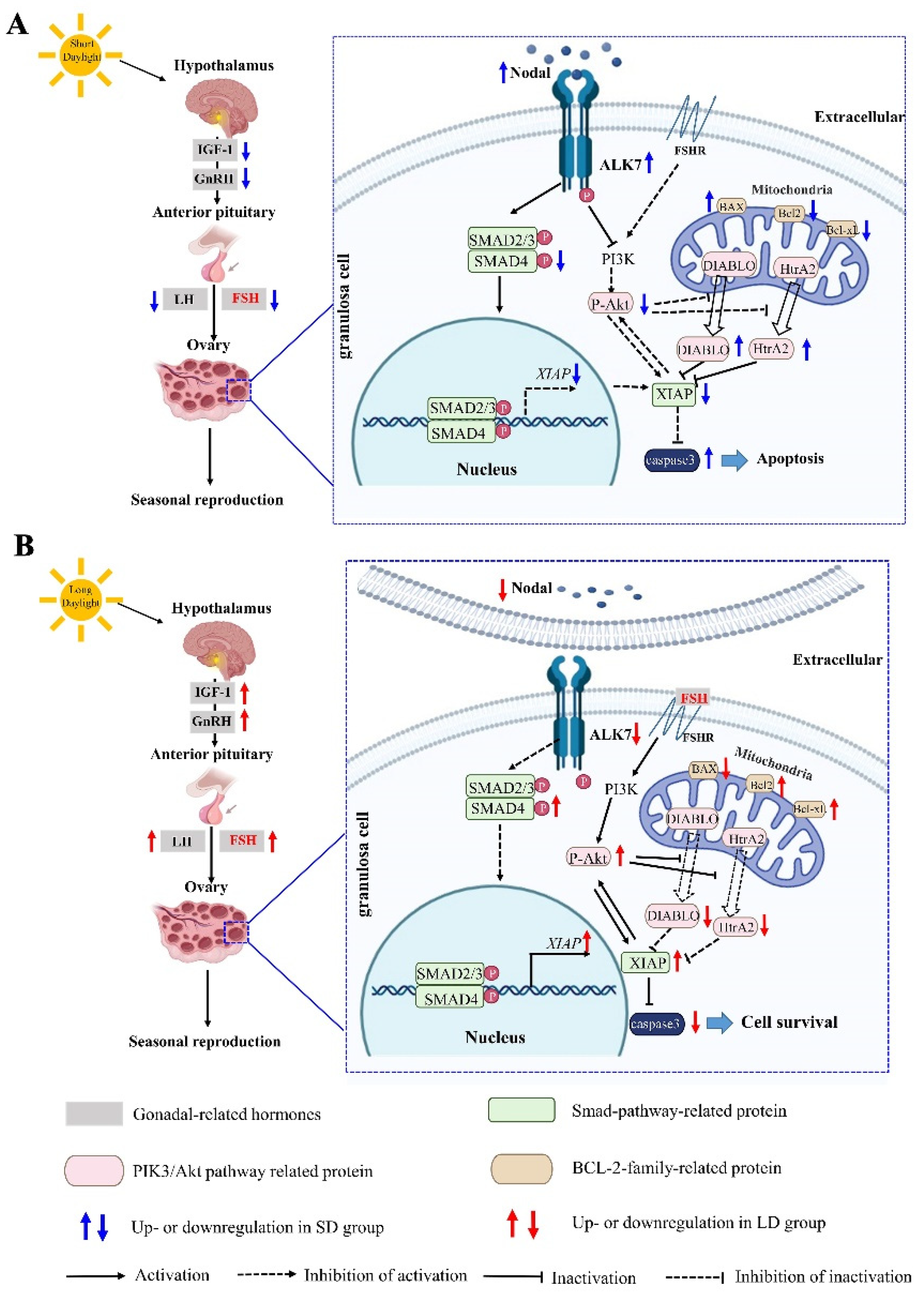
4.2. Changes in the Nodal/ALK7 Signaling Pathway in the Ovary with the Different Photoperiods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borniger, J.C.; Nelson, R.J. Photoperiodic regulation of behavior: Peromyscus as a model system. Semin. Cell Dev. Biol. 2017, 61, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Ming-Hui, X.; Yong-Zhi, Z.; Lu-Lu, L.; Guang-Ling, Z.; Wei-Hua, S.; Wan-Li, N.; Ming-Jing, Q.; Hao-Liang, C. Effect of Photoperiod on Longevity, Food Consumption, and Reproduction of Holotrichia oblita (Coleoptera: Scarabaeidae). Environ. Entomol. 2021, 50, 1151–1157. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, W.; Wang, Z.; Zheng, N.; Yuan, F.; Li, B.; Li, X.; Deng, L.; Lin, M.; Chen, X.; et al. Enhanced glycolysis in granulosa cells promotes the activation of primordial follicles through mTOR signaling. Cell Death Dis. 2022, 13, 87. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, R.; Ouyang, N.; Dai, K.; Yuan, P.; Zheng, L.; Wang, W. Investigating the impact of local inflammation on granulosa cells and follicular development in women with ovarian endometriosis. Fertil. Steril. 2019, 112, 882–891. [Google Scholar] [CrossRef]
- Han, Y.; Wang, S.; Wang, Y.; Zeng, S. IGF-1 inhibits apoptosis of porcine primary granulosa cell by targeting degradation of BimEL. Int. J. Mol. Sci. 2019, 20, 5356. [Google Scholar] [CrossRef]
- Kwintkiewicz, J.; Giudice, L.C. The interplay of insulin-like growth factors, gonadotropins, and endocrine disruptors in ovarian follicular development and function. Semin. Reprod. Med. 2009, 27, 43–51. [Google Scholar] [CrossRef]
- Chun, S.Y.; Eisenhauer, K.M.; Minami, S.; Billig, H.; Perlas, E.; Hsueh, A.J. Hormonal regulation of apoptosis in early antral follicles: Follicle-stimulating hormone as a major survival factor. Endocrinology 1996, 137, 1447–1456. [Google Scholar] [CrossRef]
- Galas, J.; Clarke, J.R.; Soltys, Z.; Stoklosowa, S. Effect of photoperiod on cultured granulosa cells of the bank vole, Clethrionomys glareolus. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2005, 140, 187–194. [Google Scholar] [CrossRef]
- Zhou, X.; Sasaki, H.; Lowe, L.; Hogan, B.L.; Kuehn, M.R. Nodal is a novel TGF-beta-like gene expressed in the mouse node during gastrulation. Nature 1993, 361, 543–547. [Google Scholar] [CrossRef]
- Tzavlaki, K.; Moustakas, A. TGF-beta signaling. Biomolecules 2020, 10, 487. [Google Scholar] [CrossRef]
- Reissmann, E.; Jornvall, H.; Blokzijl, A.; Andersson, O.; Chang, C.; Minchiotti, G.; Persico, M.G.; Ibanez, C.F.; Brivanlou, A.H. The orphan receptor ALK7 and the Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate development. Genes Dev. 2001, 15, 2010–2022. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, J.Y.; Zhu, C.; Peng, C.; Tsang, B.K. Role and regulation of nodal/activin receptor-like kinase 7 signaling pathway in the control of ovarian follicular atresia. Mol. Endocrinol. 2006, 20, 2469–2482. [Google Scholar] [CrossRef] [PubMed]
- Partheniou, F.; Kelsey, S.M.; Srinivasula, S.M.; Newland, A.C.; Alnemri, E.S.; Jia, L. c-IAP1 blocks TNFalpha-mediated cytotoxicity upstream of caspase-dependent and -independent mitochondrial events in human leukemic cells. Biochem. Biophys. Res. Commun. 2001, 287, 181–189. [Google Scholar] [CrossRef]
- Roy, N.; Deveraux, Q.L.; Takahashi, R.; Salvesen, G.S.; Reed, J.C. The c-IAP-1 and c-IAP-2 proteins are direct inhibitors of specific caspases. EMBO J. 1997, 16, 6914–6925. [Google Scholar] [CrossRef] [PubMed]
- Asselin, E.; Wang, Y.; Tsang, B.K. X-linked inhibitor of apoptosis protein activates the phosphatidylinositol 3-kinase/Akt pathway in rat granulosa cells during follicular development. Endocrinology 2001, 142, 2451–2457. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vincent, T.; Neve, E.P.; Johnson, J.R.; Kukalev, A.; Rojo, F.; Albanell, J.; Pietras, K.; Virtanen, I.; Philipson, L.; Leopold, P.L.; et al. A SNAIL1-SMAD3/4 transcriptional repressor complex promotes TGF-beta mediated epithelial-mesenchymal transition. Nat. Cell Biol. 2009, 11, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Du, X.; Wei, S.; Li, D.; Li, Q. A comprehensive transcriptomic view on the role of SMAD4 gene by RNAi-mediated knockdown in porcine follicular granulosa cells. Reproduction 2016, 152, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Zhang, L.; Li, X.; Pan, Z.; Liu, H.; Li, Q. TGF-beta signaling controls FSHR signaling-reduced ovarian granulosa cell apoptosis through the SMAD4/miR-143 axis. Cell Death Dis. 2016, 7, e2476. [Google Scholar] [CrossRef]
- Dan, H.C.; Sun, M.; Kaneko, S.; Feldman, R.I.; Nicosia, S.V.; Wang, H.G.; Tsang, B.K.; Cheng, J.Q. Akt phosphorylation and stabilization of X-linked inhibitor of apoptosis protein (XIAP). J. Biol. Chem. 2016, 291, 22846. [Google Scholar] [CrossRef]
- Du, C.; Fang, M.; Li, Y.; Li, L.; Wang, X. Smac, a mitochondrial protein that promotes cytochrome c-dependent caspase activation by eliminating IAP inhibition. Cell 2000, 102, 33–42. [Google Scholar] [CrossRef]
- Srinivasula, S.M.; Gupta, S.; Datta, P.; Zhang, Z.; Hegde, R.; Cheong, N.; Fernandes-Alnemri, T.; Alnemri, E.S. Inhibitor of apoptosis proteins are substrates for the mitochondrial serine protease Omi/HtrA2. J. Biol. Chem. 2003, 278, 31469–31472. [Google Scholar] [CrossRef] [PubMed]
- Bhola, P.D.; Letai, A. Mitochondria-Judges and Executioners of Cell Death Sentences. Mol. Cell 2016, 61, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.E.; Schlamp, C.L.; Nickells, R.W. BAX to basics: How the BCL2 gene family controls the death of retinal ganglion cells. Prog. Retin. Eye Res. 2017, 57, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Er, E.; Oliver, L.; Cartron, P.F.; Juin, P.; Manon, S.; Vallette, F.M. Mitochondria as the target of the pro-apoptotic protein BAX. Biochim. Biophys. Acta 2006, 1757, 1301–1311. [Google Scholar] [CrossRef] [PubMed]
- Antonsson, B.; Conti, F.; Ciavatta, A.; Montessuit, S.; Lewis, S.; Martinou, I.; Bernasconi, L.; Bernard, A.; Mermod, J.J.; Mazzei, G.; et al. Inhibition of BAX channel-forming activity by Bcl-2. Science 1997, 277, 370–372. [Google Scholar] [CrossRef] [PubMed]
- Manickam, P.; Kaushik, A.; Karunakaran, C.; Bhansali, S. Recent advances in cytochrome c biosensing technologies. Biosens. Bioelectron. 2017, 87, 654–668. [Google Scholar] [CrossRef]
- Matikainen, T.; Perez, G.I.; Zheng, T.S.; Kluzak, T.R.; Rueda, B.R.; Flavell, R.A.; Tilly, J.L. Caspase-3 gene knockout defines cell lineage specificity for programmed cell death signaling in the ovary. Endocrinology 2001, 142, 2468–2480. [Google Scholar] [CrossRef]
- Munley, K.M.; Rendon, N.M.; Demas, G.E. Neural Androgen Synthesis and Aggression: Insights From a Seasonally Breeding Rodent. Front. Endocrinol. 2018, 9, 136. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Xue, H.; Li, S.; Xu, J.; Chen, L. Seasonal differential expression of KiSS-1/GPR54 in the striped hamsters (Cricetulus barabensis) among different tissues. Integr. Zool. 2017, 12, 260–268. [Google Scholar] [CrossRef]
- Ohba, Y. Form of the molar crown with especial reference to tooth wear, and occlusion in rats. Aichi Gakuin Daigaku Shigakkai Shi 1974, 12, 171–192. [Google Scholar]
- Crooks, P.V.; O’Reilly, C.B.; Owens, P.D. Microscopy of the dentine of enamel-free areas of rat molar teeth. Arch. Oral Biol. 1983, 28, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Barnett, N.A.; Rowe, D.J. A comparison of alveolar bone in young and aged mice. J. Periodontol. 1986, 57, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Ledevin, R.; Quéré, J.P.; Renaud, S. Morphometrics as an insight into processes beyond tooth shape variation in a bank vole population. PLoS ONE 2010, 5, e15470. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, K.; Illnerova, H. Photoperiodic effects in the Djungarian hamster. Rate of testicular regression and extension of pineal melatonin pattern depend on the way of change from long to short photoperiods. Neuroendocrinology 1986, 43, 317–321. [Google Scholar] [CrossRef]
- Knopper, L.D.; Boily, P. The energy budget of captive Siberian hamsters, Phodopus sungorus, exposed to photoperiod changes: Mass loss is caused by a voluntary decrease in food intake. Physiol. Biochem. Zool. 2000, 73, 517–522. [Google Scholar] [CrossRef]
- Prendergast, B.J.; Nelson, R.J. Spontaneous “regression” of enhanced immune function in a photoperiodic rodent Peromyscus maniculatus. Proc. Biol. Sci. 2001, 268, 2221–2228. [Google Scholar] [CrossRef]
- Tian, M.Y. Microstructure Observation on the Follicular Development of Blue Fox during the Estrous Period. J. Anhui Agric. Sci. 2011, 33, 221. [Google Scholar] [CrossRef]
- Moffatt-Blue, C.S.; Sury, J.J.; Young, K.A. Short photoperiod-induced ovarian regression is mediated by apoptosis in Siberian hamsters (Phodopus sungorus). Reproduction 2006, 131, 771–782. [Google Scholar] [CrossRef]
- Knigge, K.M.; Leathem, J.H. Growth and atresia of follicles in the ovary of the hamster. Anat. Rec. 1956, 124, 679–707. [Google Scholar] [CrossRef]
- Guldenaar, S.E.; Wang, K.; McCabe, J.T. Double immunofluorescence staining of Fos and Jun in the hypothalamus of the rat. Cell Tissue Res. 1994, 276, 1–6. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, J.H.; Mou, J.J.; Kong, X.T.; Wu, M.; Xue, H.L.; Xu, L.X. Photoperiod Affects Harderian Gland Morphology and Secretion in Female Cricetulus barabensis: Autophagy, Apoptosis, and Mitochondria. Front. Physiol. 2020, 11, 408. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Tan, H.; Ling, H.; Yuan, X. Detecting overexpression level of HER2 gene in NSCLC by real-time quantitative PCR and the 2[-Delta Delta C(T)] method. Zhongguo Fei Ai Za Zhi 2011, 14, 938–942. [Google Scholar] [CrossRef] [PubMed]
- Hnasko, T.S.; Hnasko, R.M. The western blot. Methods Mol. Biol. 2015, 1318, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xu, J.H.; Mou, J.J.; Kong, X.T.; Zou, J.W.; Xue, H.L.; Wu, M.; Xu, L.X. Novel ultrastructural findings on cardiac mitochondria of huddling Brandt’s voles in mild cold environment. Comp. Biochem. Physiol. Part A 2020, 249, 110766. [Google Scholar] [CrossRef]
- Masson-Pevet, M.; Naimi, F.; Canguilhem, B.; Saboureau, M.; Bonn, D.; Pevet, P. Are the annual reproductive and body weight rhythms in the male European hamster (Cricetus cricetus) dependent upon a photoperiodically entrained circannual clock? J. Pineal Res. 1994, 17, 151–163. [Google Scholar] [CrossRef]
- Bertoni, J.M.; Sprenkle, P.M.; Hanifin, J.P.; Stetson, M.H.; Brainard, G.C. Effects of short photoperiod on ATPase activities in the testis of the immature Siberian hamster. Biol. Reprod. 1992, 47, 509–513. [Google Scholar] [CrossRef]
- Herwig, A.; Petri, I.; Barrett, P. Hypothalamic gene expression rapidly changes in response to photoperiod in juvenile siberian hamsters (Phodopus sungorus). J. Neuroendocrinol. 2012, 24, 991–998. [Google Scholar] [CrossRef]
- Schramm, R.D.; Bavister, B.D. Effects of granulosa cells and gonadotrophins on meiotic and developmental competence of oocytes in vitro in non-stimulated rhesus monkeys. Hum. Reprod. 1995, 10, 887–895. [Google Scholar] [CrossRef]
- Amsterdam, A.; Keren-Tal, I.; Aharoni, D.; Dantes, A.; Land-Bracha, A.; Rimon, E.; Sasson, R.; Hirsh, L. Steroidogenesis and apoptosis in the mammalian ovary. Steroids 2003, 68, 861–867. [Google Scholar] [CrossRef]
- Clarke, H.J. Regulation of germ cell development by intercellular signaling in the mammalian ovarian follicle. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e294. [Google Scholar] [CrossRef]
- Hurwitz, A.; Adashi, E.Y. Ovarian follicular atresia as an apoptotic process: A paradigm for programmed cell death in endocrine tissues. Mol. Cell. Endocrinol. 1992, 84, C19–C23. [Google Scholar] [CrossRef]
- Zhu, G.; Fang, C.; Li, J.; Mo, C.; Wang, Y.; Li, J. Transcriptomic diversification of granulosa cells during follicular development in chicken. Sci. Rep. 2019, 9, 5462. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.Y.; Cheung, C.K.; Wang, Y.; Tsang, B.K. Regulation of cell death and cell survival gene expression during ovarian follicular development and atresia. Front. Biosci. 2003, 8, 222–237. [Google Scholar] [CrossRef]
- Babu, S.R.; Sadhnani, M.D.; Swarna, M.; Padmavathi, P.; Reddy, P.P. Evaluation of FSH, LH and testosterone levels in different subgroups of infertile males. Indian J. Clin. Biochem. 2004, 19, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Xiao, L.; Ge, W.; Yang, S.; Jiang, Y.; Lv, J.; Hu, J.; Zhang, Y.; Zhao, X.; Hua, Y. Follicle-stimulating hormone and luteinizing hormone regulate the synthesis mechanism of dihydrotestosterone in sheep granulosa cells. Reprod. Domest. Anim. 2021, 56, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.F.; Lin, X.N.; Dai, Y.D.; Lin, X.; Lu, H.M.; Zhou, F.; Zhang, S.Y. The effect of follicular fluid from patients with endometriosis, follicle stimulating hormone and bone morphogenetic protein 15 on the proliferation and progesterone secretion of granular cells. Zhonghua Yi Xue Za Zhi 2017, 97, 3543–3547. [Google Scholar] [CrossRef] [PubMed]
- Daftary, S.S.; Gore, A.C. IGF-1 in the brain as a regulator of reproductive neuroendocrine function. Exp. Biol. Med. 2005, 230, 292–306. [Google Scholar] [CrossRef] [PubMed]
- Kanematsu, T.; Irahara, M.; Miyake, T.; Shitsukawa, K.; Aono, T. Effect of insulin-like growth factor I on gonadotropin release from the hypothalamus-pituitary axis in vitro. Acta Endocrinol. 1991, 125, 227–233. [Google Scholar] [CrossRef]
- Monniaux, D.; Pisselet, C. Control of proliferation and differentiation of ovine granulosa cells by insulin-like growth factor-I and follicle-stimulating hormone in vitro. Biol. Reprod. 1992, 46, 109–119. [Google Scholar] [CrossRef]
- Monniaux, D.; Huet, C.; Besnard, N.; Clement, F.; Bosc, M.; Pisselet, C.; Monget, P.; Mariana, J.C. Follicular growth and ovarian dynamics in mammals. J. Reprod. Fertil. Suppl. 1997, 51, 3–23. [Google Scholar]
- Wang, H.; Tsang, B.K. Nodal signalling and apoptosis. Reproduction 2007, 133, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chen, X.; Li, X.; Wang, L.; Zhang, H.; He, Y.; Wang, J.; Zhao, Y.; Zhang, B.; Xu, Y. Interference RNA-based silencing of endogenous SMAD4 in porcine granulosa cells resulted in decreased FSH-mediated granulosa cells proliferation and steroidogenesis. Reproduction 2011, 141, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, W.; Zhang, C.; Ji, G.; Ge, Y.; Xu, Y.; Zhao, Y. TGF-beta1 inhibits the growth and metastasis of tongue squamous carcinoma cells through Smad4. Gene 2011, 485, 160–166. [Google Scholar] [CrossRef]
- Vyas, S.; Juin, P.; Hancock, D.; Suzuki, Y.; Takahashi, R.; Triller, A.; Evan, G. Differentiation-dependent sensitivity to apoptogenic factors in PC12 cells. J. Biol. Chem. 2004, 279, 30983–30993. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.H. Mitochondrial dysfunction and oxidative stress as determinants of cell death/survival in stroke. Ann. N. Y. Acad. Sci. 2005, 1042, 203–209. [Google Scholar] [CrossRef]
- Sun, C.; Cai, M.; Gunasekera, A.H.; Meadows, R.P.; Wang, H.; Chen, J.; Zhang, H.; Wu, W.; Xu, N.; Ng, S.C.; et al. NMR structure and mutagenesis of the inhibitor-of-apoptosis protein XIAP. Nature 1999, 401, 818–822. [Google Scholar] [CrossRef]
- Martins, L.M.; Iaccarino, I.; Tenev, T.; Gschmeissner, S.; Totty, N.F.; Lemoine, N.R.; Savopoulos, J.; Gray, C.W.; Creasy, C.L.; Dingwall, C.; et al. The serine protease Omi/HtrA2 regulates apoptosis by binding XIAP through a reaper-like motif. J. Biol. Chem. 2002, 277, 439–444. [Google Scholar] [CrossRef]
- van Loo, G.; van Gurp, M.; Depuydt, B.; Srinivasula, S.M.; Rodriguez, I.; Alnemri, E.S.; Gevaert, K.; Vandekerckhove, J.; Declercq, W.; Vandenabeele, P. The serine protease Omi/HtrA2 is released from mitochondria during apoptosis. Omi interacts with caspase-inhibitor XIAP and induces enhanced caspase activity. Cell Death Differ. 2002, 9, 20–26. [Google Scholar] [CrossRef]
- Winkler, J.; Rand, M.L.; Schmugge, M.; Speer, O. Omi/HtrA2 and XIAP are components of platelet apoptosis signalling. Thromb. Haemost. 2013, 109, 532–539. [Google Scholar] [CrossRef]
- Wu, G.; Chai, J.; Suber, T.L.; Wu, J.W.; Du, C.; Wang, X.; Shi, Y. Structural basis of IAP recognition by Smac/DIABLO. Nature 2000, 408, 1008–1112. [Google Scholar] [CrossRef]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Edlich, F. BCL-2 proteins and apoptosis: Recent insights and unknowns. Biochem. Biophys. Res. Commun. 2018, 500, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Morales, E.; Ferrer, C.; Zuasti, A.; Garcia-Borron, J.C.; Canteras, M.; Pastor, L.M. Apoptosis and molecular pathways in the seminiferous epithelium of aged and photoinhibited Syrian hamsters (Mesocricetus auratus). J. Androl. 2007, 28, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Russell, L.D.; Chiarini-Garcia, H.; Korsmeyer, S.J.; Knudson, C.M. Bax-dependent spermatogonia apoptosis is required for testicular development and spermatogenesis. Biol. Reprod. 2002, 66, 950–958. [Google Scholar] [CrossRef]
- Shannonhouse, J.L.; Urbanski, H.F.; Woo, S.L.; Fong, L.A.; Goddard, S.D.; Lucas, W.F.; Jones, E.R.; Wu, C.; Morgan, C. Aquaporin-11 control of testicular fertility markers in Syrian hamsters. Mol. Cell. Endocrinol. 2014, 391, 1–9. [Google Scholar] [CrossRef]
- Chai, J.; Shiozaki, E.; Srinivasula, S.M.; Wu, Q.; Datta, P.; Alnemri, E.S.; Shi, Y. Structural basis of caspase-7 inhibition by XIAP. Cell 2001, 104, 769–780. [Google Scholar] [CrossRef]
- Cossu, F.; Milani, M.; Vachette, P.; Malvezzi, F.; Grassi, S.; Lecis, D.; Delia, D.; Drago, C.; Seneci, P.; Bolognesi, M.; et al. Structural insight into inhibitor of apoptosis proteins recognition by a potent divalent smac-mimetic. PLoS ONE 2012, 7, e49527. [Google Scholar] [CrossRef]
- Hu, G.; Cao, C.; Deng, Z.; Li, J.; Zhou, X.; Huang, Z.; Cen, C. Effects of matrine in combination with cisplatin on liver cancer. Oncol. Lett. 2021, 21, 66. [Google Scholar] [CrossRef]
- Huang, Y.; Park, Y.C.; Rich, R.L.; Segal, D.; Myszka, D.G.; Wu, H. Structural basis of caspase inhibition by XIAP: Differential roles of the linker versus the BIR domain. Cell 2001, 104, 781–790. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | SD | MD | LD | |
|---|---|---|---|---|
| Prior to the photoperiod | BW (g) | 32.5750 ± 0.28982 | 32.4800 ± 0.35715 | 32.2200 ± 0.26937 |
| Following the photoperiod | BW (g) | 32.5050 ± 0.411640 | 33.0250 ± 0.25944 | 33.200 ± 0.22478 |
| OW (g) | 0.0264 ± 0.00071 | 0.0283 ± 0.00059 * | 0.02882 ± 0.00065 * | |
| OW/BW (mg/g) | 0.8002 ± 0.02402 | 0.8579 ± 0.02045 | 0.8697 ± 0.02217 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, Y.; Xue, H.-L.; Zheng, Y.-J.; Yin, Y.-F.; Xu, W.-L.; Xu, J.-H.; Wu, M.; Chen, L.; Xu, L.-X. The Photoperiod Regulates Granulosa Cell Apoptosis through the FSH-Nodal/ALK7 Signaling Pathway in Phodopus sungorus. Animals 2022, 12, 3570. https://doi.org/10.3390/ani12243570
Qi Y, Xue H-L, Zheng Y-J, Yin Y-F, Xu W-L, Xu J-H, Wu M, Chen L, Xu L-X. The Photoperiod Regulates Granulosa Cell Apoptosis through the FSH-Nodal/ALK7 Signaling Pathway in Phodopus sungorus. Animals. 2022; 12(24):3570. https://doi.org/10.3390/ani12243570
Chicago/Turabian StyleQi, Yan, Hui-Liang Xue, Yun-Jiao Zheng, Yun-Fei Yin, Wen-Lei Xu, Jin-Hui Xu, Ming Wu, Lei Chen, and Lai-Xiang Xu. 2022. "The Photoperiod Regulates Granulosa Cell Apoptosis through the FSH-Nodal/ALK7 Signaling Pathway in Phodopus sungorus" Animals 12, no. 24: 3570. https://doi.org/10.3390/ani12243570
APA StyleQi, Y., Xue, H.-L., Zheng, Y.-J., Yin, Y.-F., Xu, W.-L., Xu, J.-H., Wu, M., Chen, L., & Xu, L.-X. (2022). The Photoperiod Regulates Granulosa Cell Apoptosis through the FSH-Nodal/ALK7 Signaling Pathway in Phodopus sungorus. Animals, 12(24), 3570. https://doi.org/10.3390/ani12243570