
The Aleutians and Beyond: Distribution, Size Composition, and Catch Dynamics of the Aleutian Skate Bathyraja aleutica across the North Pacific



Abstract
Simple Summary
Abstract
1. Introduction
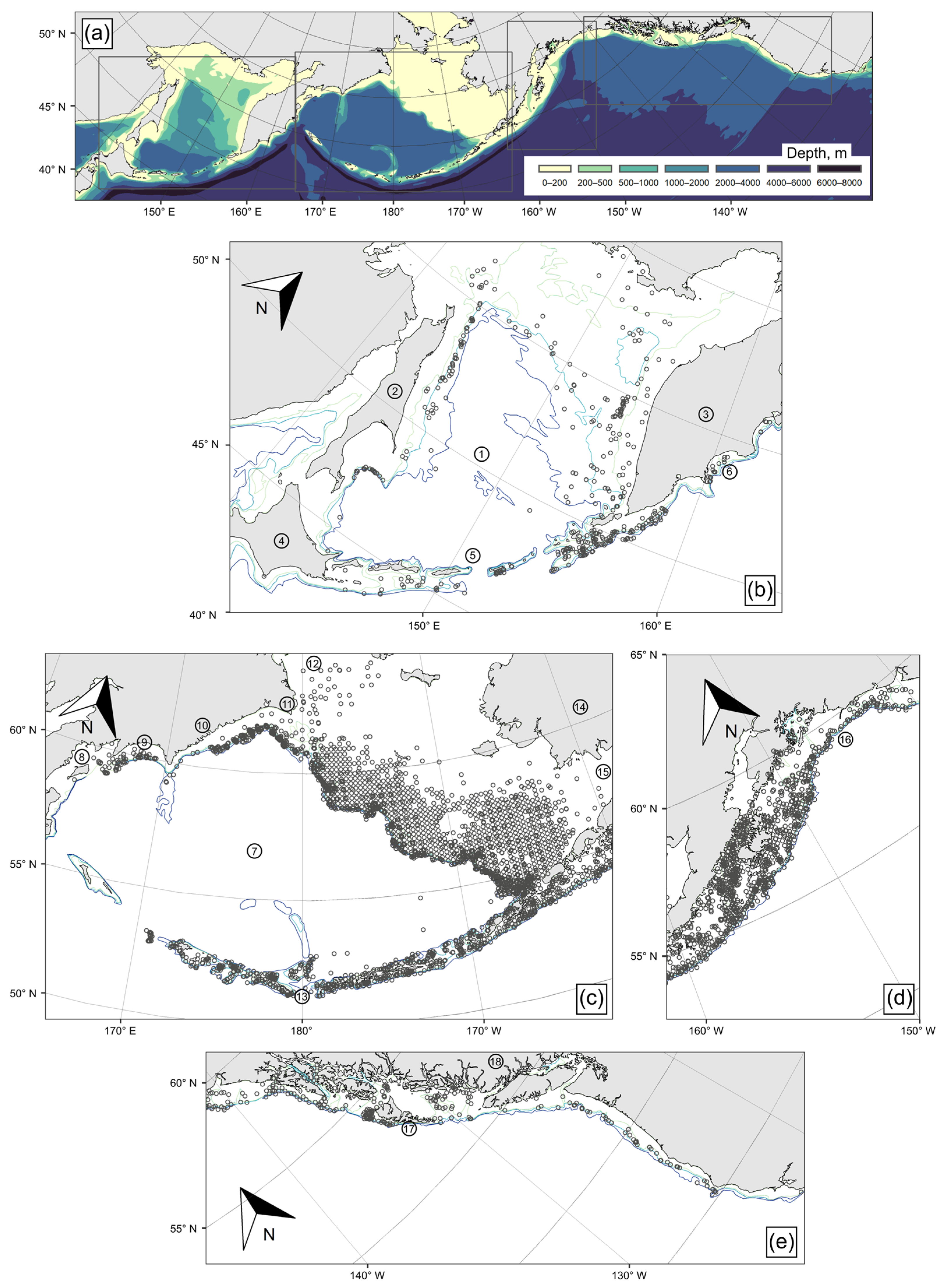
2. Materials and Methods
3. Results
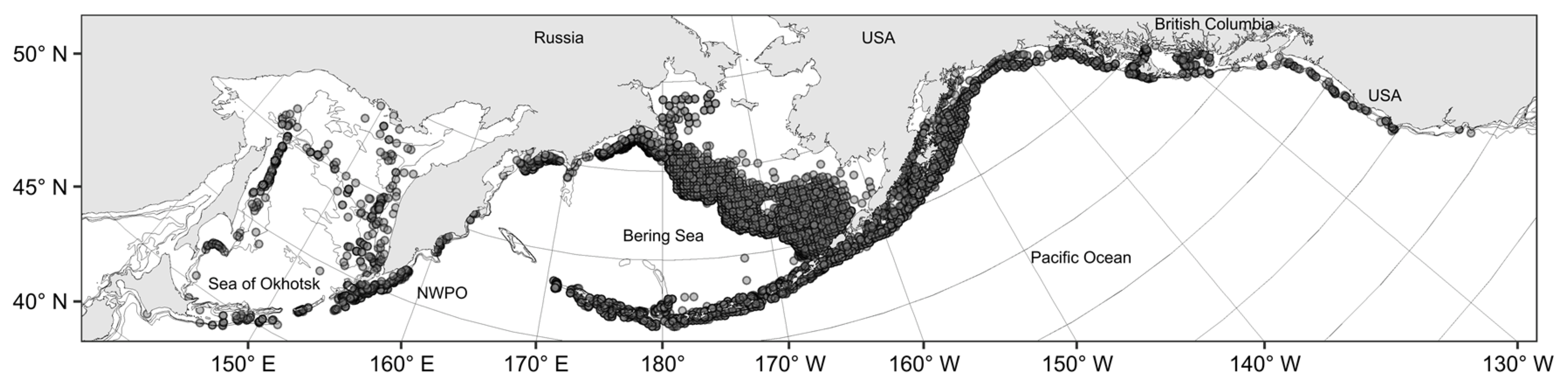
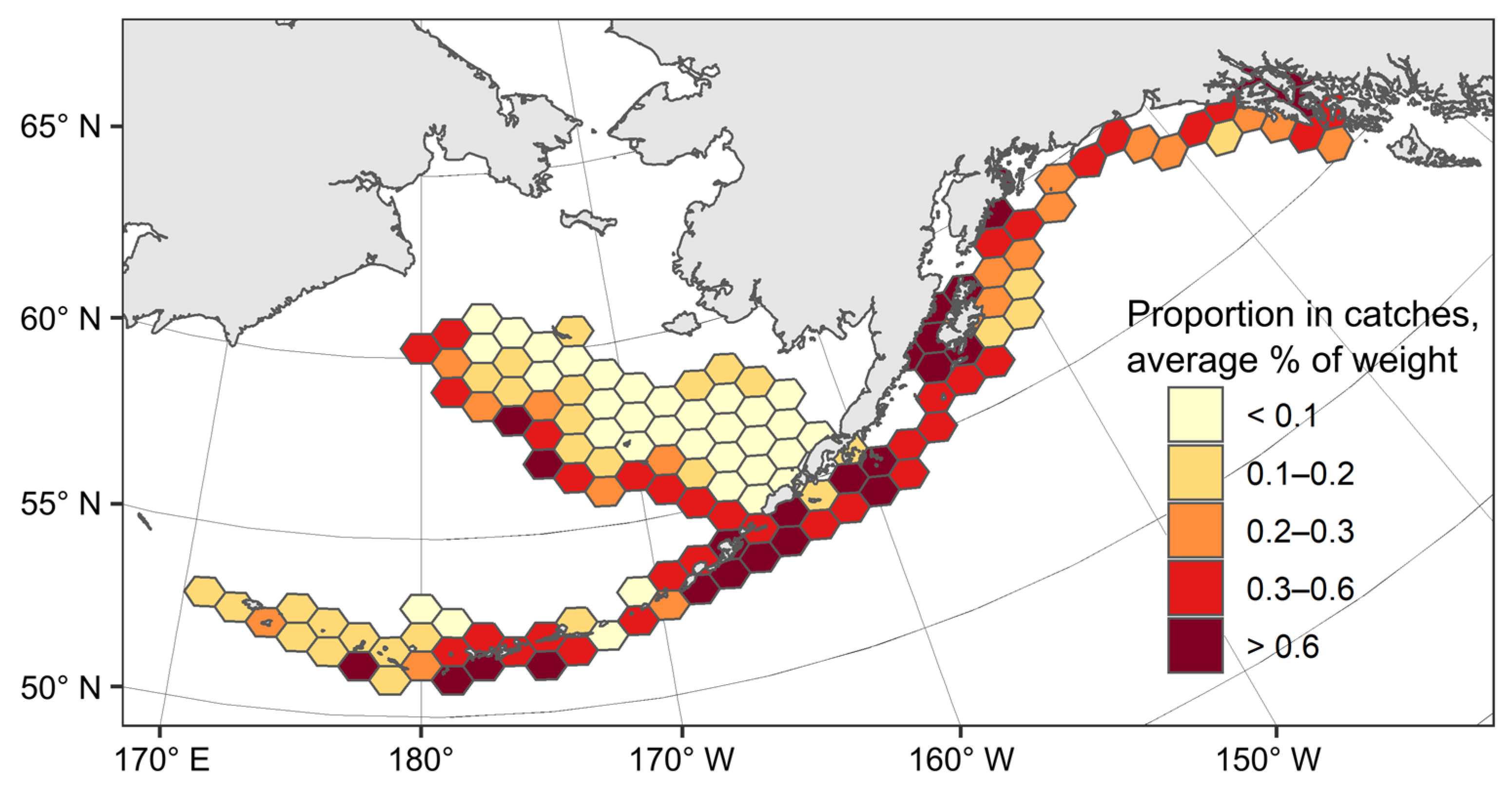
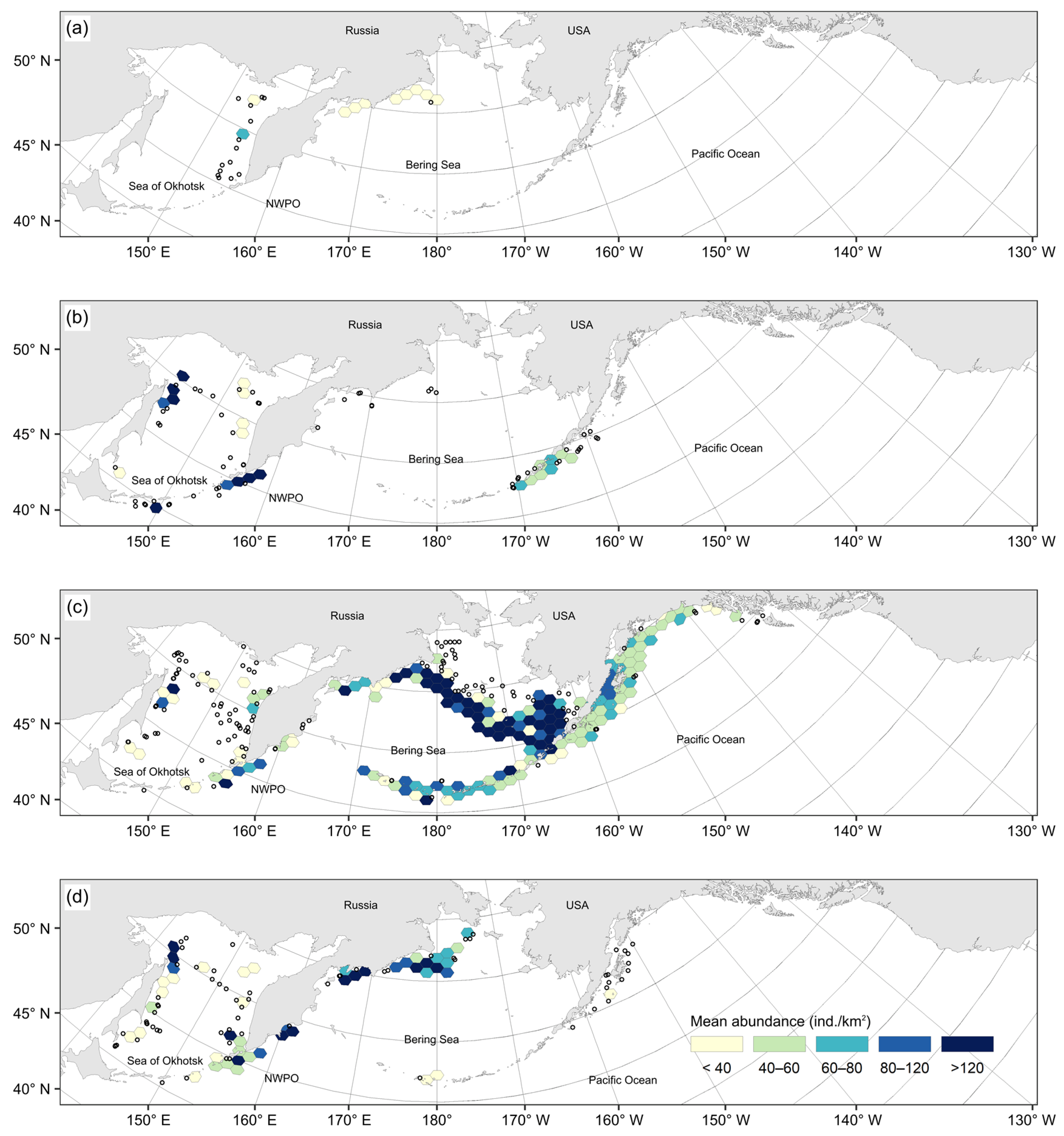
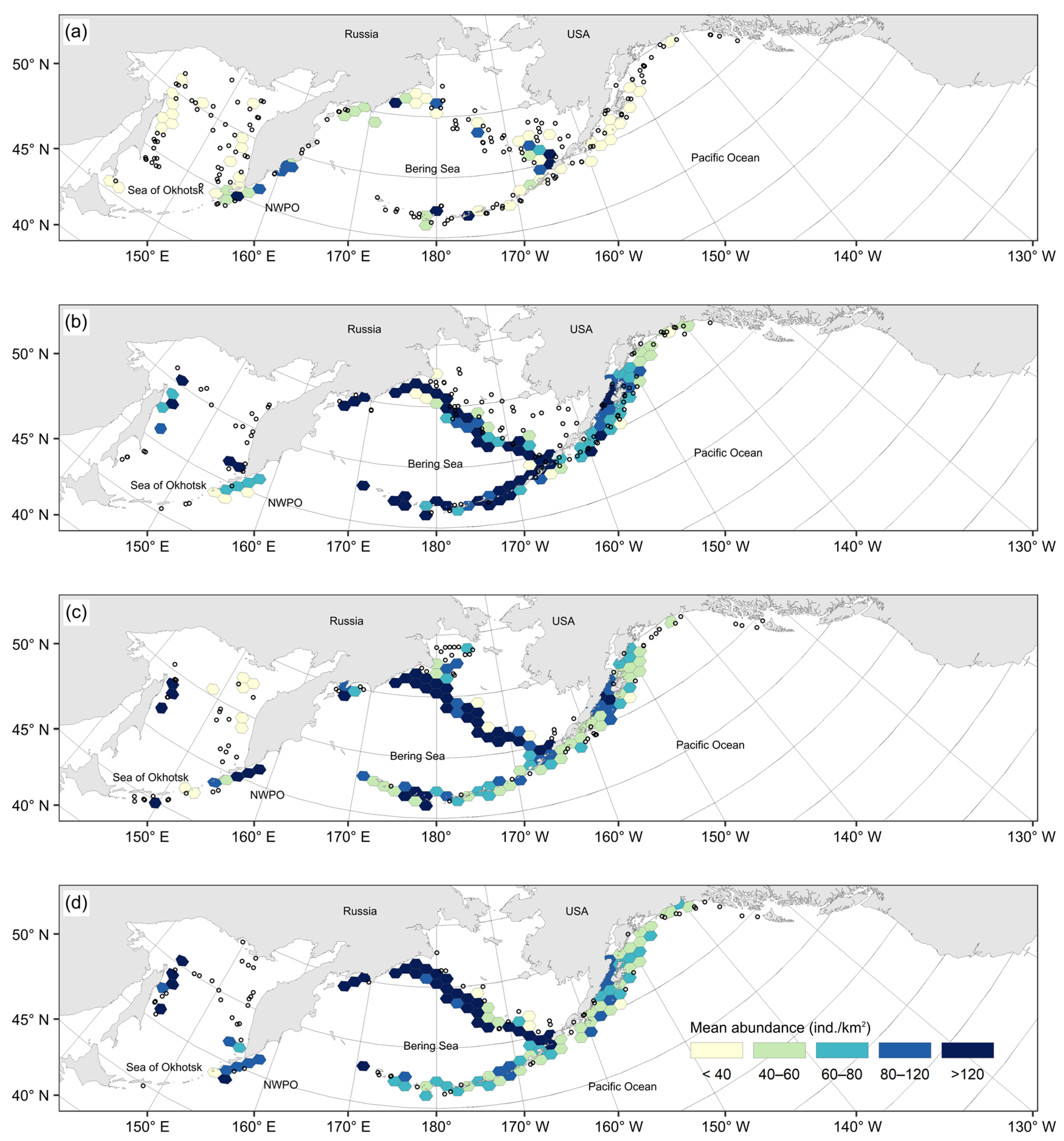
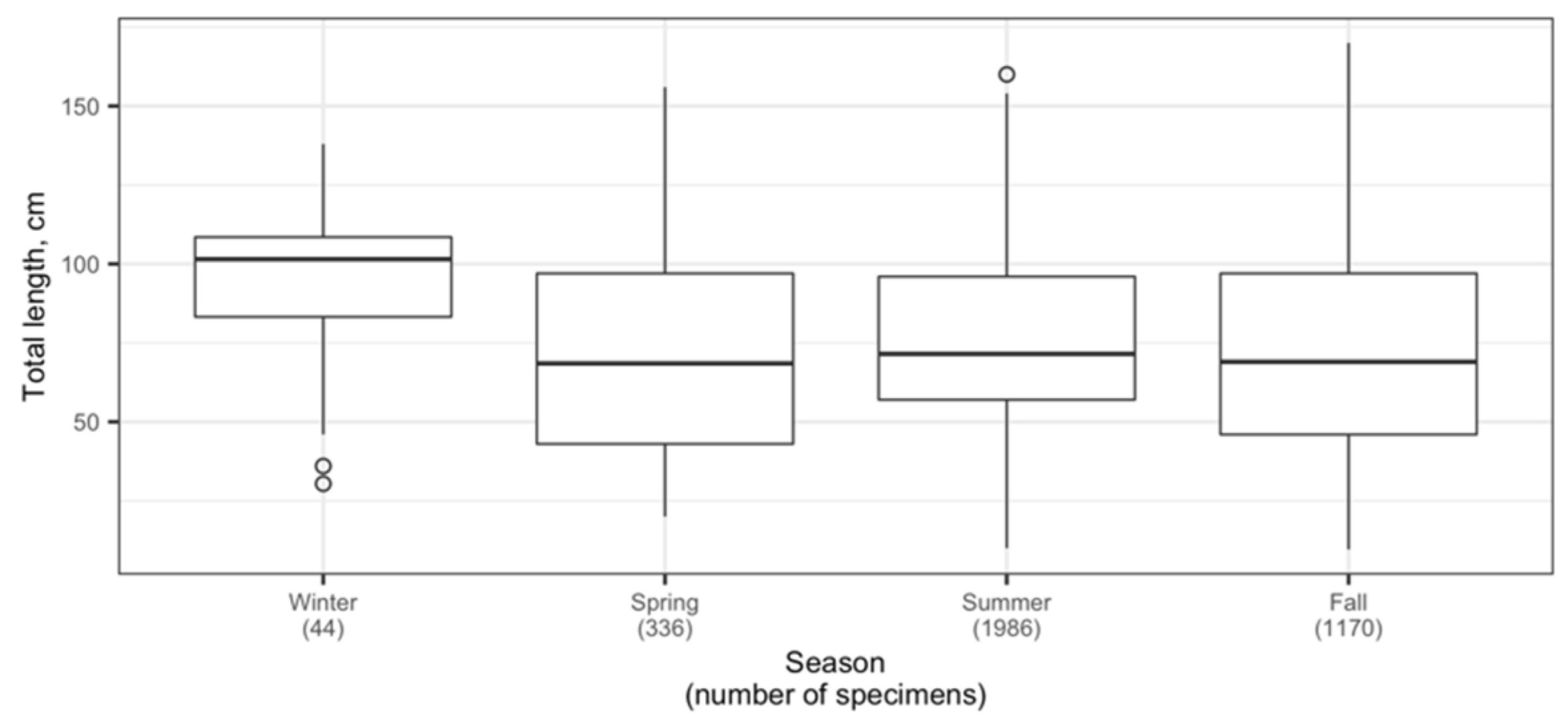
3.1. Spatial Distribution of Catches and Their Seasonal and Long-Term Changes
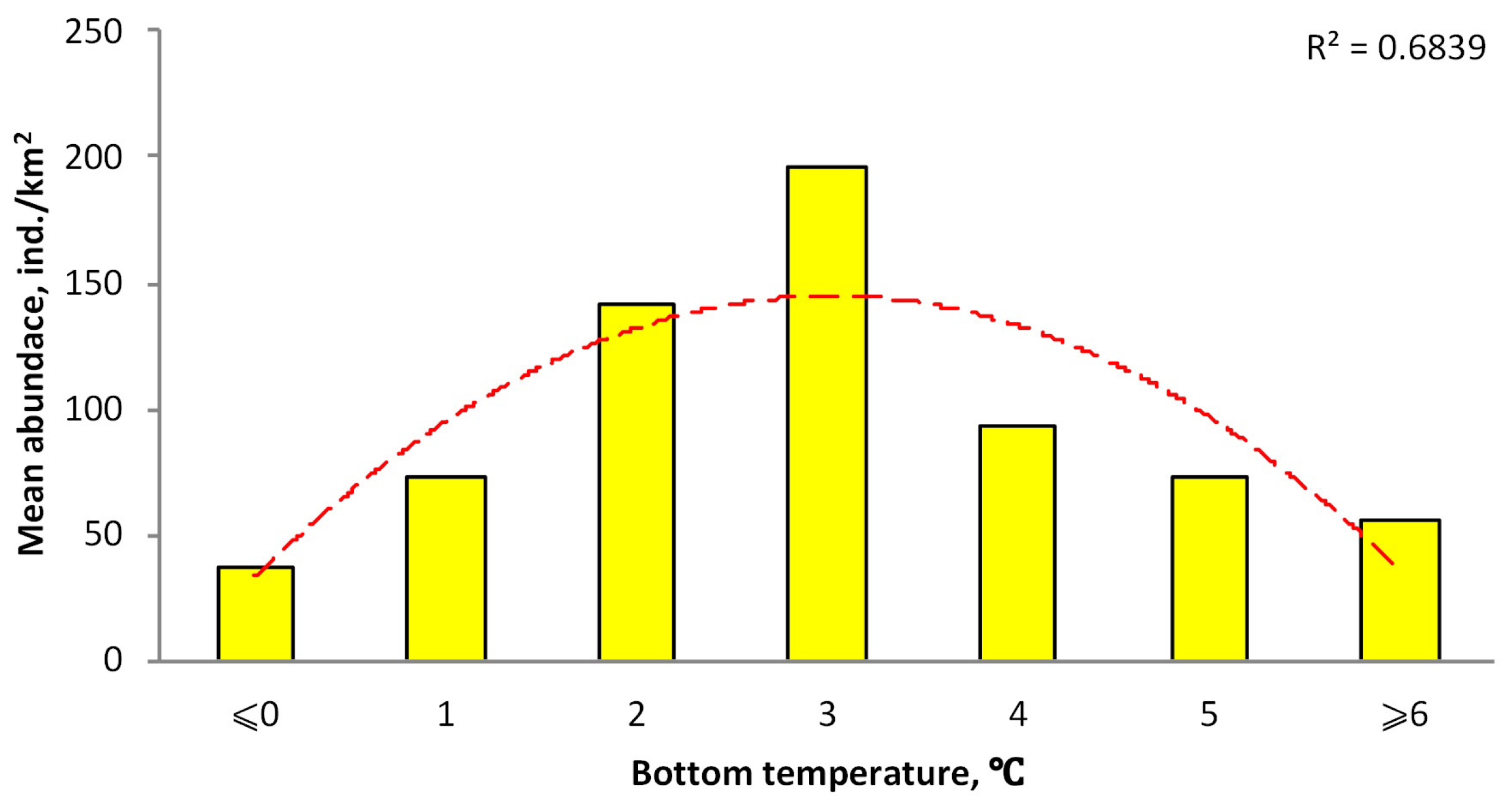
3.2. Distribution Depending on Bottom Temperatures
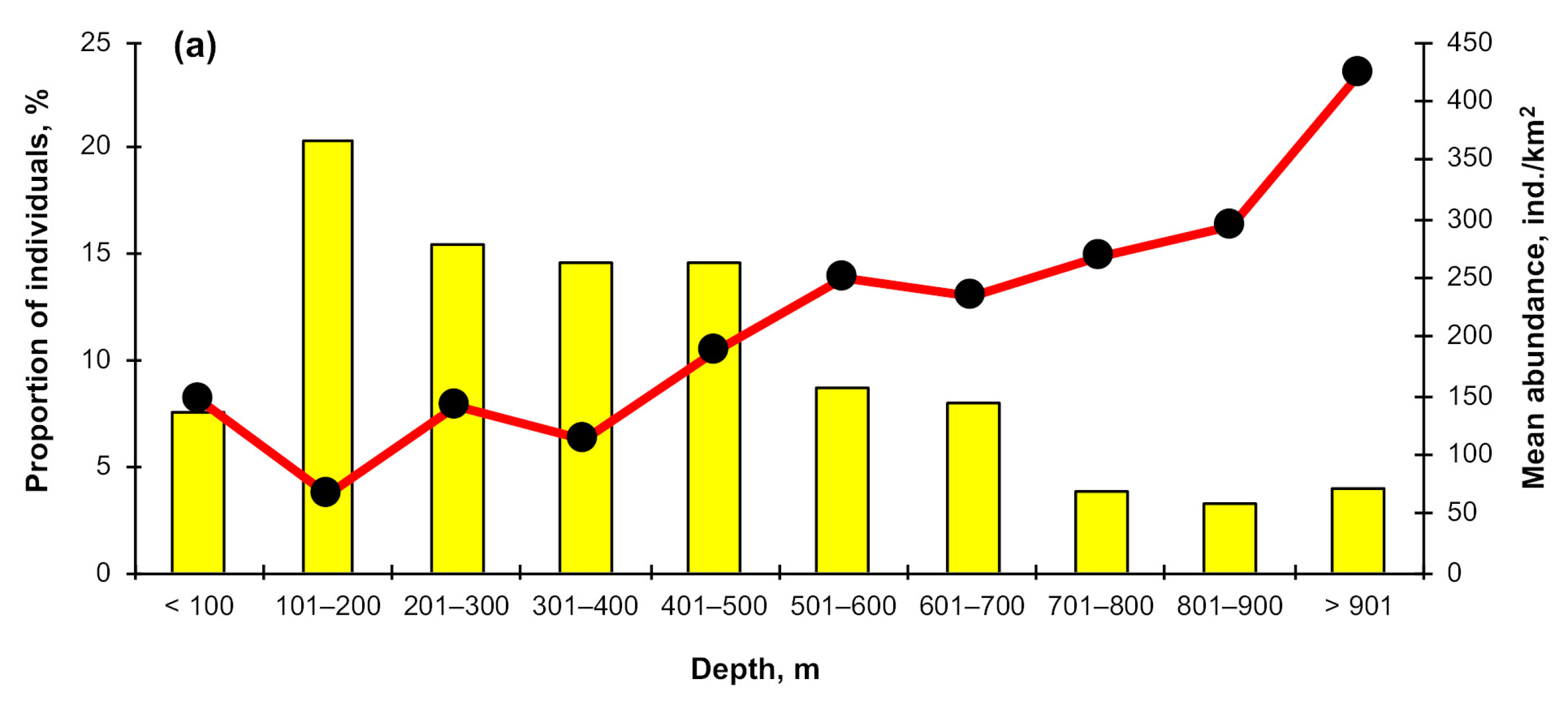
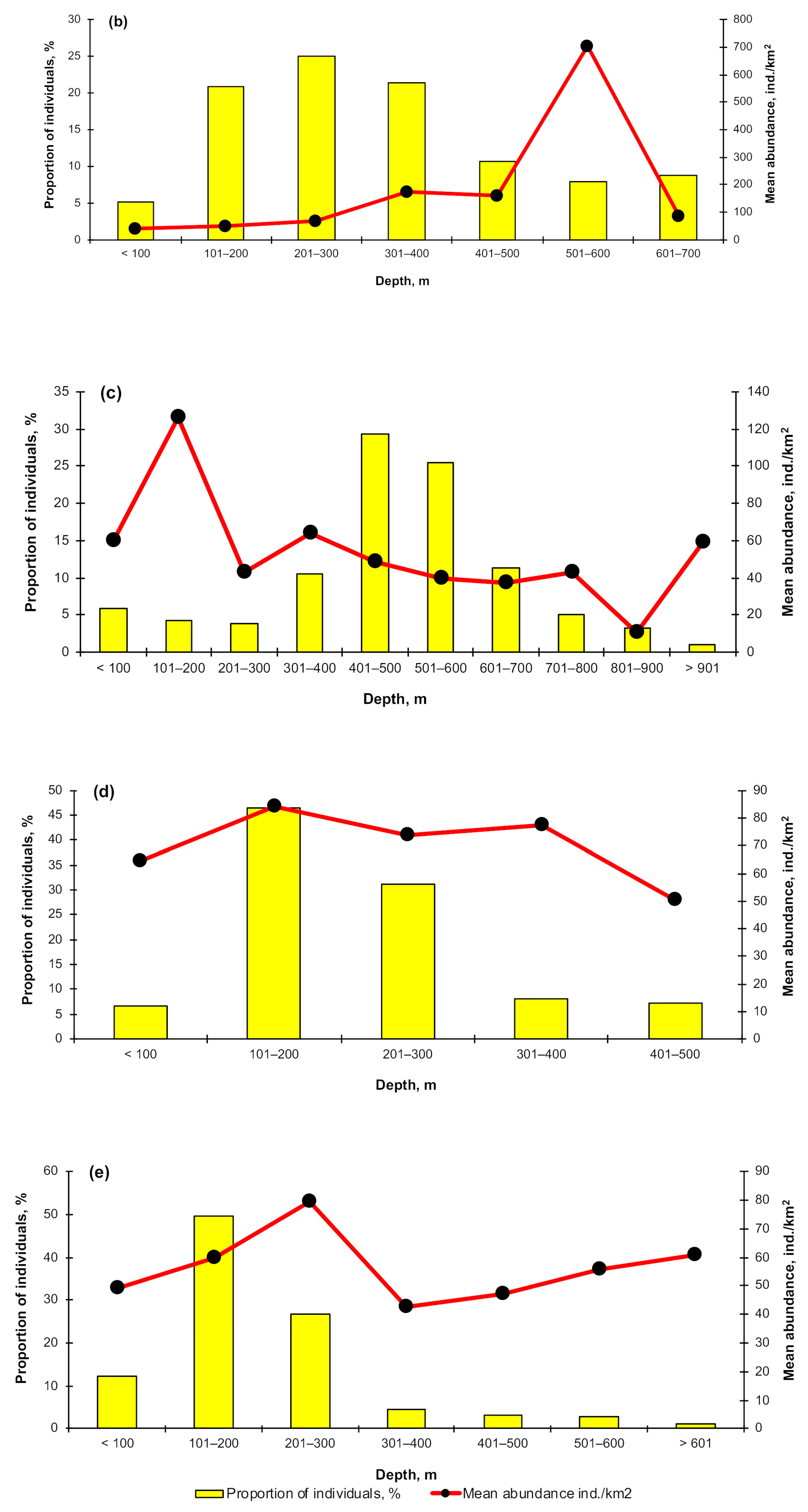
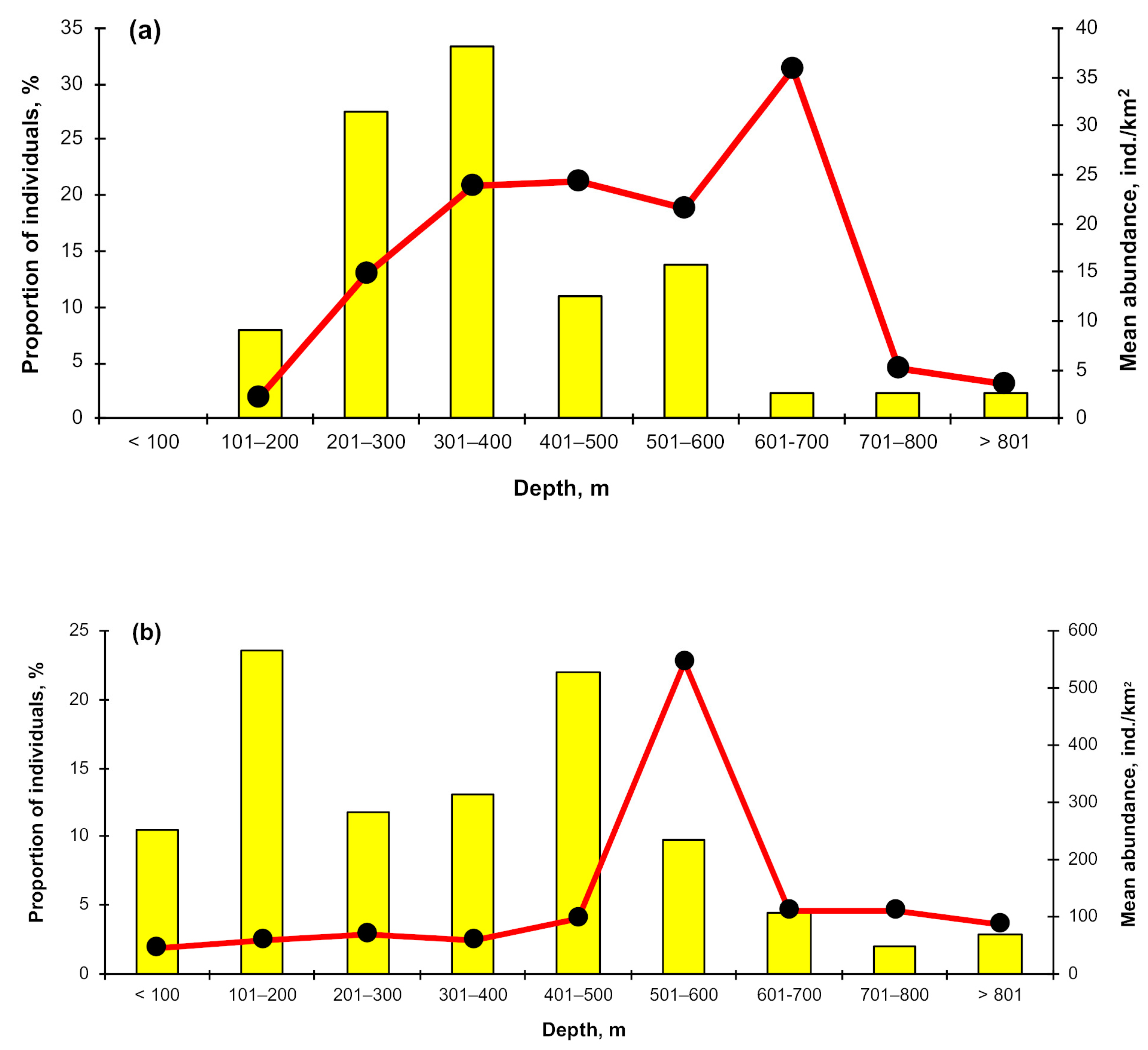
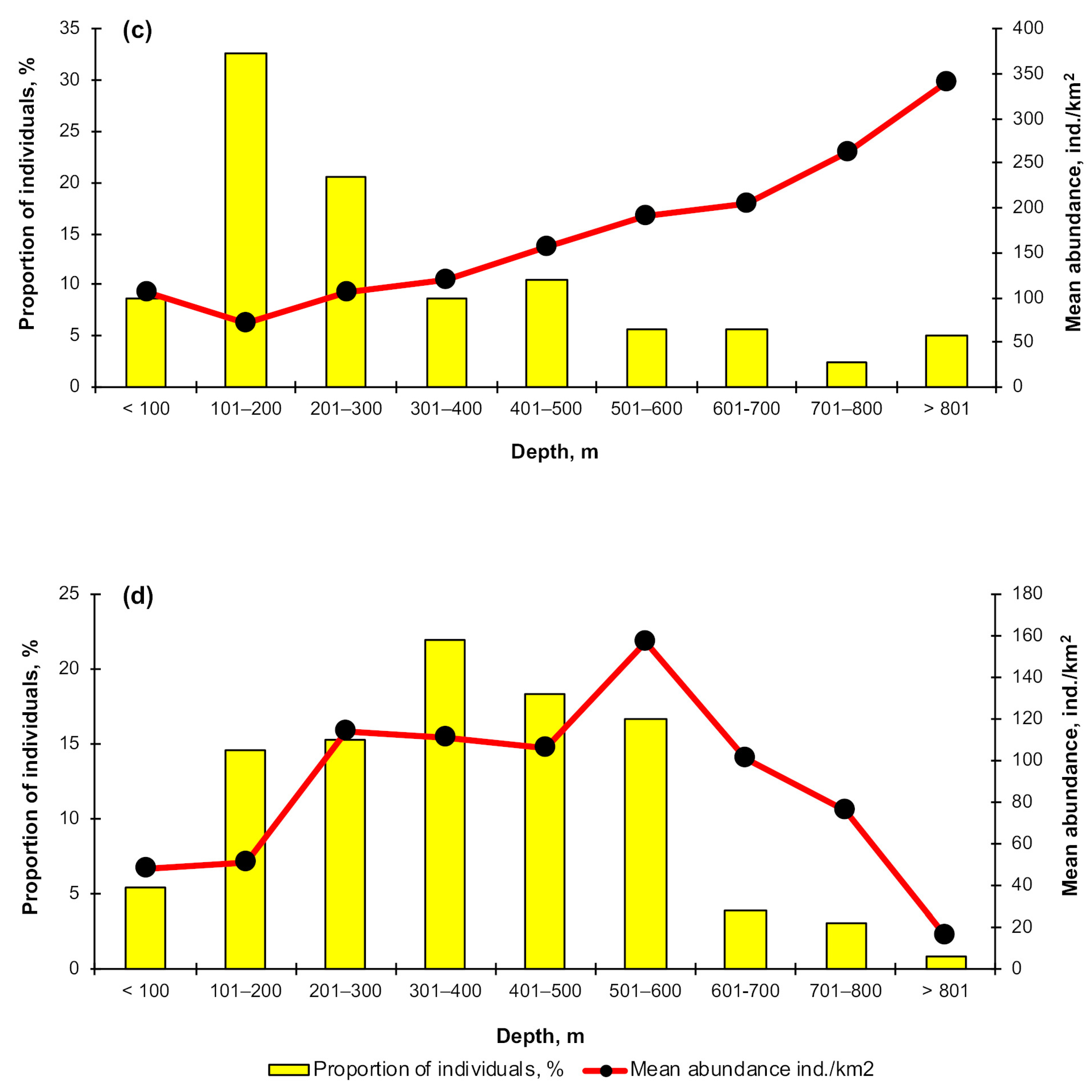
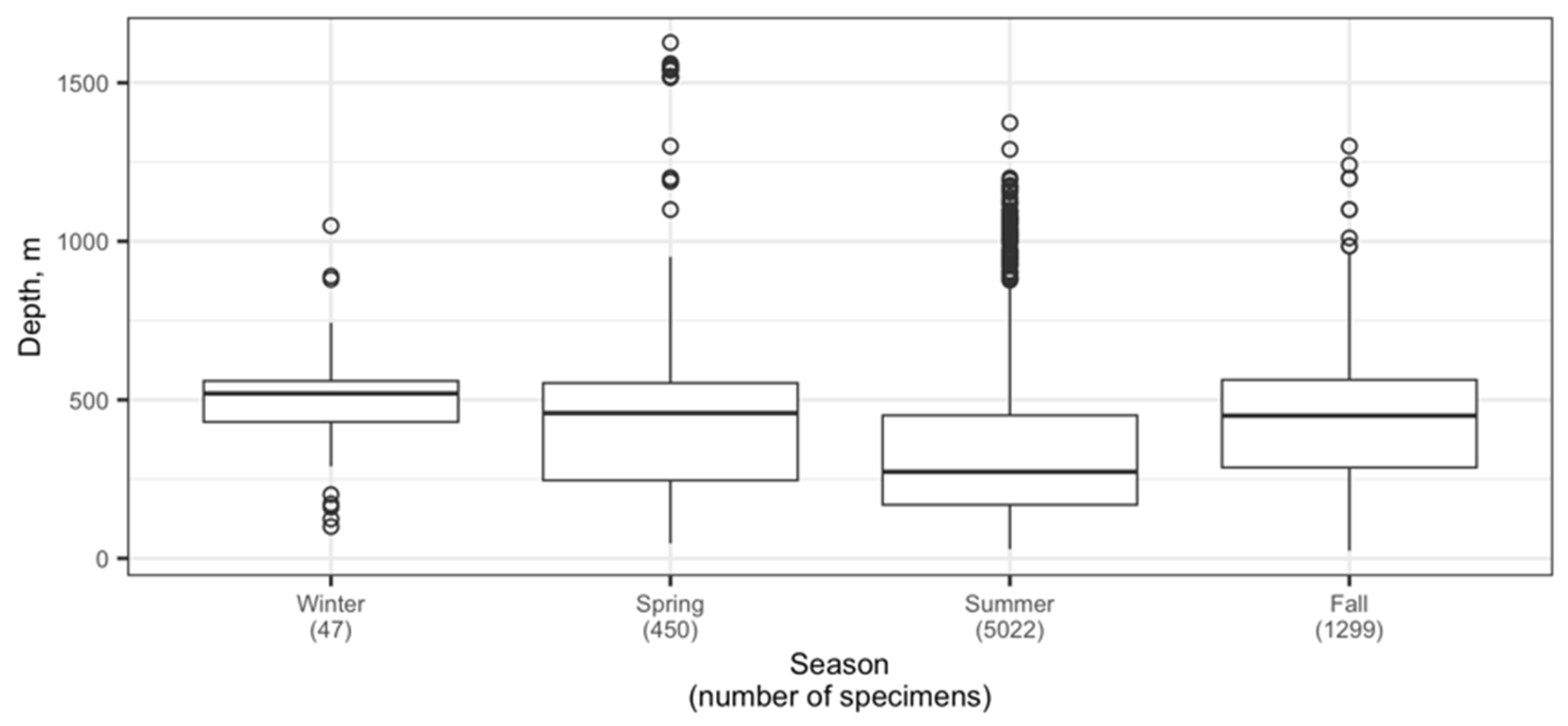
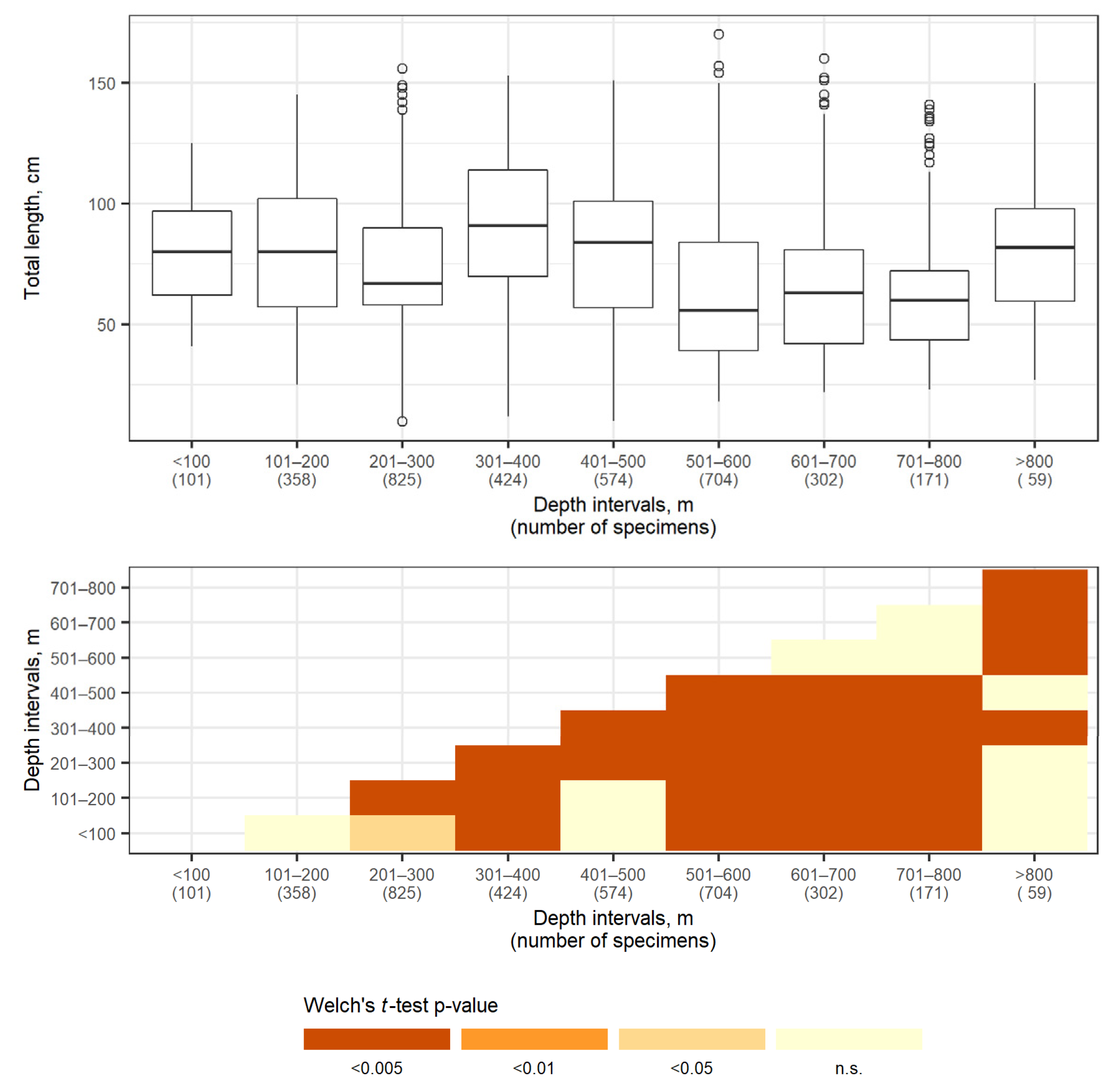
3.3. Depth Distribution
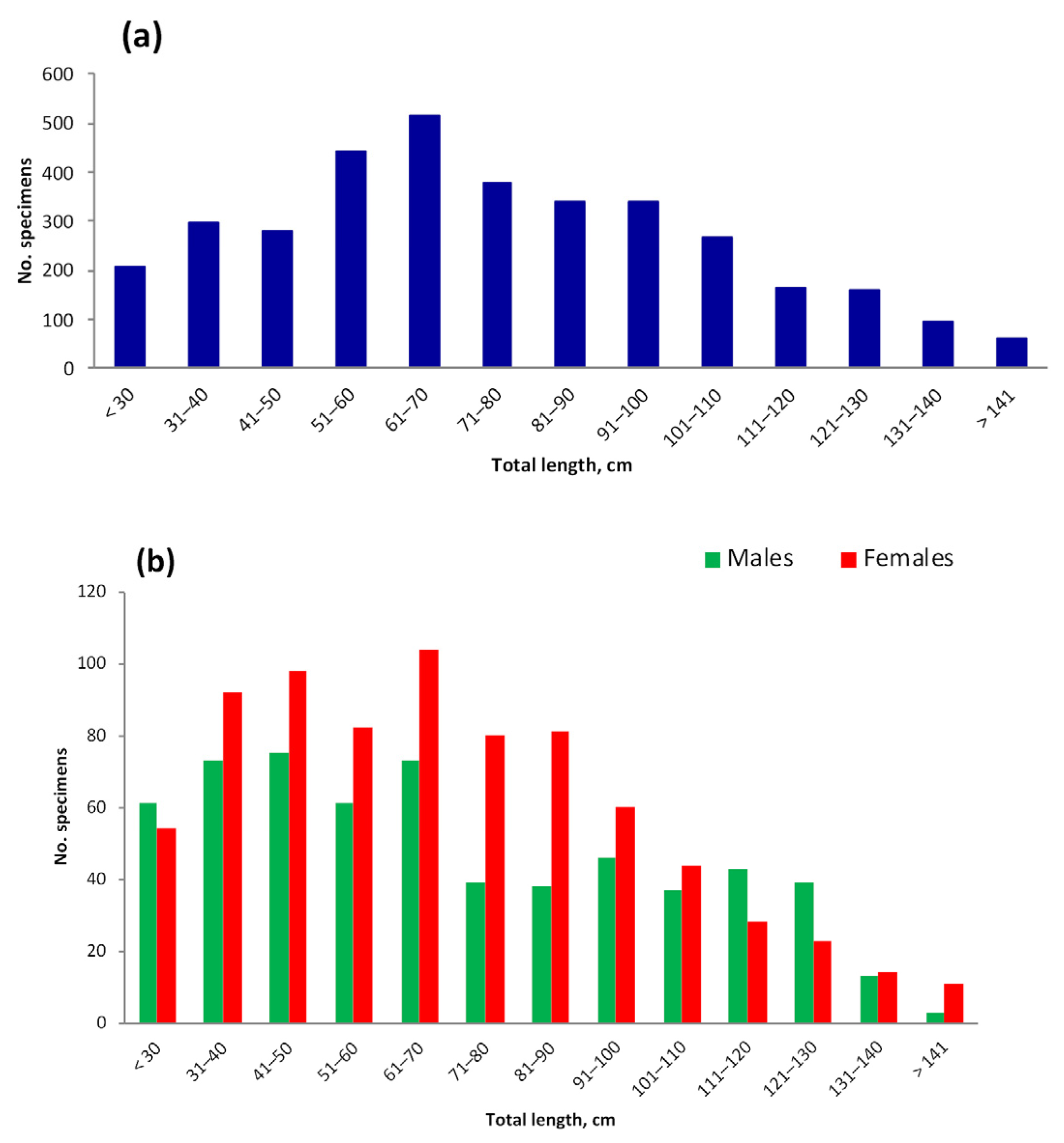
3.4. Length and Weight
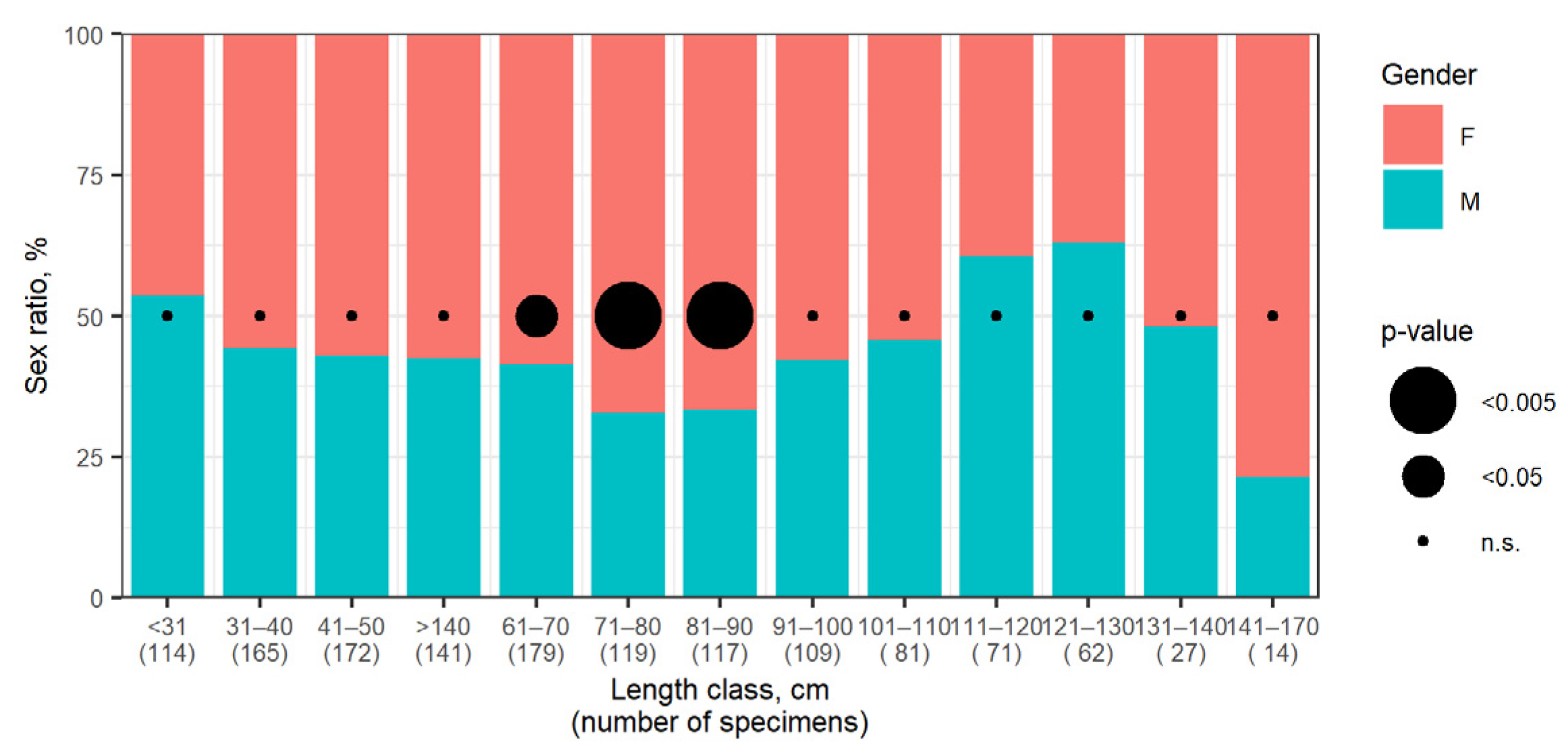
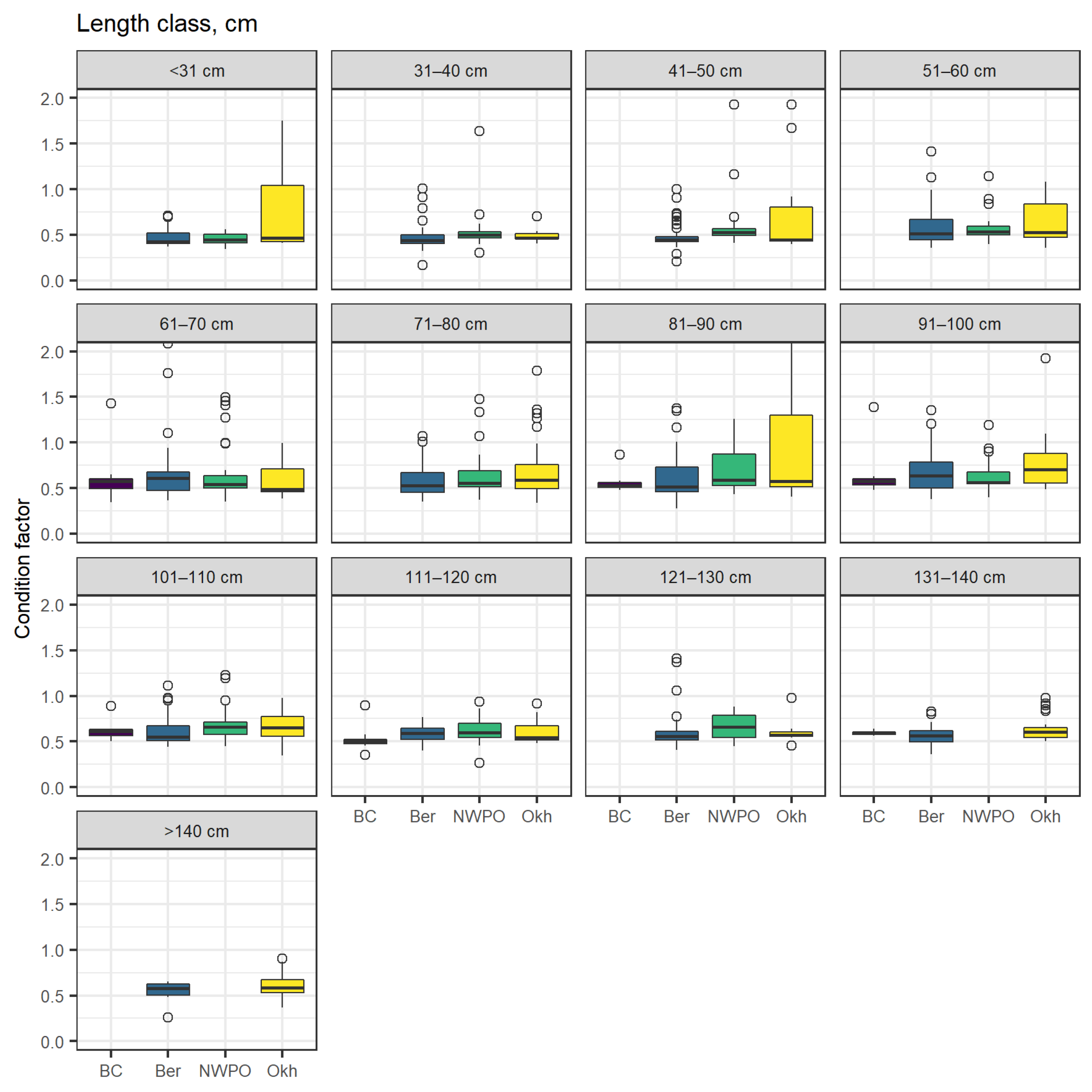
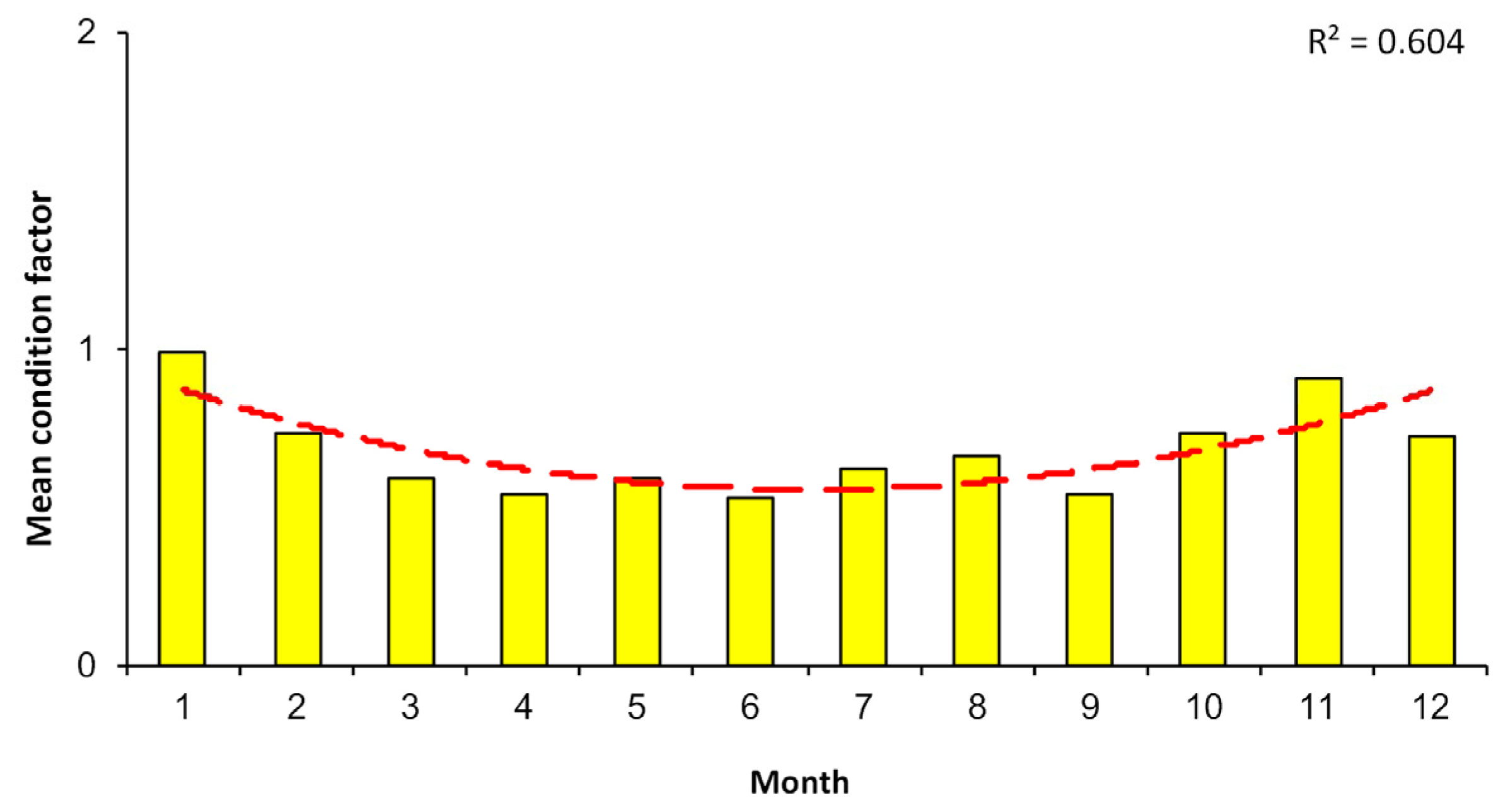
3.5. Sex Ratio and Condition Factor
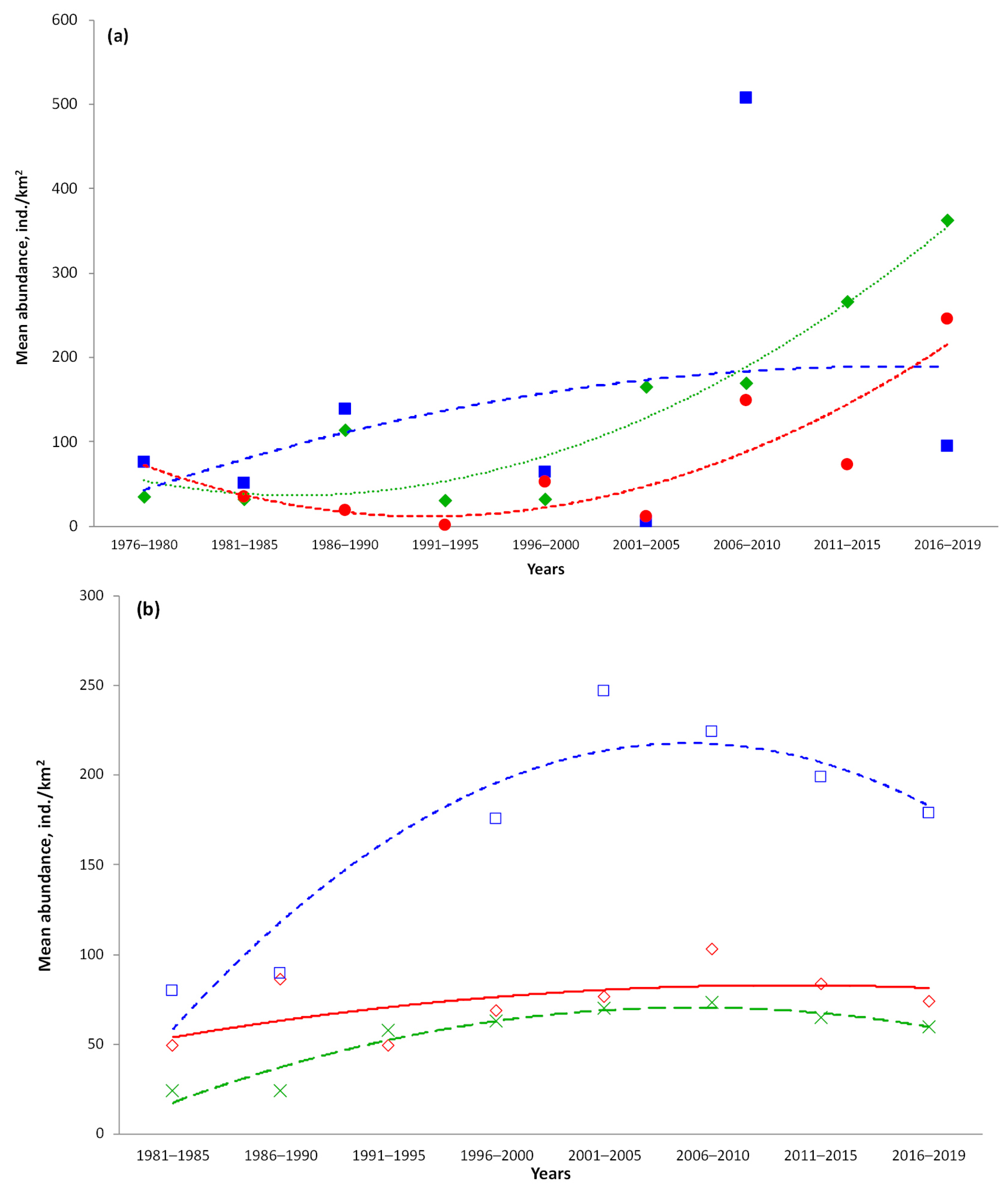
3.6. Catch Dynamics
4. Discussion
4.1. Spatial Distribution
4.2. Depth Distribution
4.3. Distribution Depending on Bottom Temperatures
4.4. Length and Weight
4.5. Sex Ratio and Condition Factor
4.6. Catch Dynamics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mito, K. Food Relationships among Benthic Fish Populations in the Bering Sea on the Theragra chalcogramma Fishing Grounds in October and November of 1972. Master’s Thesis, Hokkaido University Graduate School, Hakodate, Japan, 1974; pp. 1–135. [Google Scholar]
- Brodeur, R.D.; Livingston, P.A. Food habits and diet overlap of various Eastern Bering Sea fishes. U.S. Dep. Commer. NOAA Tech. Memo. NMFS F/NWC 1988, 127, 1–76. [Google Scholar]
- Livingston, P.A.; de Reynier, Y. Groundfish food habits and predation on commercially important prey species in the Eastern Bering Sea from 1990 to 1992. U.S. Dep. Commer. NOAA/NMFS Alaska Fish. Sci. Cent. Proc. Rep. 1996, 96, 1–214. [Google Scholar]
- Orlov, A.M. The diets and feeding habits of some deep-water benthic skates (Rajidae) in the Pacific waters off the northern Kuril Islands and southeastern Kamchatka. Alaska Fish. Res. Bull. 1998, 5, 1–17. [Google Scholar]
- Orlov, A.M. Feeding habits of some deep-benthic skates (Rajidae) in the western Bering Sea. In Proceedings of the MTS/IEEE Oceans 2001. An Ocean Odyssey, Honolulu, HI, USA, 5–8 November 2001; Volume 2, pp. 842–855. [Google Scholar] [CrossRef]
- Orlov, A.M. Diets, feeding habits, and trophic relations of six deep-benthic skates (Rajidae) in the western Bering Sea. Aqua. J. Ichthyol. Aquat. Biol. 2003, 7, 45–60. [Google Scholar]
- Chuchukalo, V.I.; Lapko, V.V.; Kuznetsova, N.A.; Slabinsky, A.M.; Napazakov, V.V.; Nadtochy, V.A.; Koblikov, V.N.; Pushchina, O.I. Feeding of groundfishes on the shelf and continental slope of the northern Sea of Okhotsk. Izv. TINRO 1999, 126, 24–57. [Google Scholar]
- Chuchukalo, V.I.; Napazakov, V.V. Feeding and trophic state of the mass species of skates (Rajidae) from the western part of the Bering Sea. Izv. TINRO 2002, 130, 422–428. [Google Scholar]
- Ishihara, H. The skates and rays of the western North Pacific: An overview of their fisheries, utilization, and classification. U.S. Dep. Commer. NOAA Tech. Rep. NMFS 1990, 90, 485–497. [Google Scholar]
- Orlov, A.M. Bycatch reduction of multispecies bottom trawl fisheries: Some approaches to solve the problem in the Northwest Pacific. Am. Fish. Soc. Symp. 2007, 49, 241–258. [Google Scholar]
- Orlov, A.M. Some approaches to reducing non-commercial bycatch of bottom trawl fisheries in the western Bering Sea. Asian Fish. Sci. 2011, 24, 397–412. [Google Scholar] [CrossRef]
- Dolganov, V.N. Reserves of the skates in Far Eastern seas of Russia and their prospective fishery. Izv. TINRO 1999, 126, 650–652. [Google Scholar]
- Orlov, A.M.; Volvenko, I.V. Long-term Changes of Distribution and Abundance of Nine Deep-water Skates (Arhynchobatidae: Rajiformes: Chondrichthyes) in the Northwestern Pacific. Animals 2022, 12, 3485. [Google Scholar] [CrossRef]
- Nasedkina, E.A. Characteristics of flesh of skates. Rybn. Khoz. 1969, 6, 59–60. [Google Scholar]
- Baidalova, G.F.; Dubnitskaya, G.M.; Nechaev, A.P.; Severinenko, S.M. Chemical composition and properties of thorny skate liver oil depending on the fishing season. Rybn. Khoz. 1984, 10, 60–62. [Google Scholar]
- Orlov, A.M. Groundfish resources of the northern North Pacific continental slope: From science to sustainable fishery. In Proceedings of the Seventh North Pacific Rim Fisheries Conference, Busan, Republic of Korea, 18–20 May 2004; Alaska Pacific University: Anchorage, AK, USA, 2005; pp. 139–150. [Google Scholar]
- Dvinin, Y.F.; Konstantinova, L.L.; Kuz’mina, V.I. Techno-chemical characteristics of some skates and chimaeras. Tr. PINRO 1981, 47, 29–51. [Google Scholar]
- Orlov, A.M. Substantiation of commercial size of Far Eastern skates (fam. Rajidae) by example of widespread species from the Bering Sea. Tr. VNIRO 2006, 146, 252–264. [Google Scholar]
- Ormseth, O.A. Assessment of the skate stock complex in the Bering Sea and Aleutian Islands. In Stock Assessment and Fishery Evaluation Report for the Groundfish Resources of the Bering Sea/Aleutian Islands Regions; North Pacific Fishery Management Council: Anchorage, AK, USA, 2012; pp. 1647–1734. [Google Scholar]
- Dolganov, V.N. Guide to the Diagnostic Characters of Cartilaginous Fishes from the Far East Seas of the U.S.S.R. and Neighboring Waters; TINRO Vladivostok: Vladivostok, Russia, 1983; pp. 1–92. [Google Scholar]
- Mecklenburg, C.; Mecklenburg, T.; Thorsteinson, L. Fishes of Alaska; American Fisheries Society: Bethesda, MA, USA, 2002; pp. 1–1037. [Google Scholar]
- Hoff, G.R. New records of the Aleutian skate, Bathyraja aleutica, from northern California. Calif. Fish Game 2002, 88, 145–148. [Google Scholar]
- Fedorov, V.V.; Chereshnev, I.A.; Nazarkin, M.V.; Shestakov, A.V.; Volobuev, V.V. Catalogue of Marine and Freshwater Fishes in Northern Sea of Okhotsk; Dal’nauka: Vladivostok, Russia, 2003; pp. 1–204. [Google Scholar]
- Shinohara, G.; Narimatsu, Y.; Hattori, T.; Ito, M.; Takata, Y.; Matsuura, K. Annotated checklist of deep-sea fishes from the Pacific coast off Tohoku District, Japan. Natl. Mus. Nat. Sci. Monogr. 2009, 39, 683–735. [Google Scholar]
- Lynghammar, A.; Christiansen, J.S.; Mecklenburg, C.W.; Karamushko, O.V.; Møller, P.R.; Gallucci, V.F. Species richness and distribution of chondrichthyan fishes in the Arctic Ocean and adjacent seas. Biodiversity 2013, 14, 57–66. [Google Scholar] [CrossRef][Green Version]
- Grigorov, I.V.; Orlov, A.M. Species diversity and conservation status of cartilaginous fishes (Chondrichthyes) of Russian waters. J. Ichthyol. 2013, 53, 923–936. [Google Scholar] [CrossRef]
- Parin, N.V.; Evseenko, S.A.; Vasil’eva, E.D. Fishes of Russian seas: Annotated Catalogue; KMK Scientific Press: Moscow, Russia, 2014; pp. 1–733. [Google Scholar]
- Tuponogov, V.N.; Kodolov, L.S. Handbook for Identification of the Commercial and Mass Species of Fishes of Far Eastern Seas of Russia; Russkiy Ostrov: Vladivostok, Russia, 2014; pp. 1–336. [Google Scholar]
- Dyldin, Y.V. Annotated checklist of the sharks, batoids and chimaeras (Chondrichthyes: Elasmobranchii, Holocephali) from waters of Russia and adjacent areas. Publ. Seto Mar. Biol. Lab. 2015, 43, 40–91. [Google Scholar] [CrossRef]
- Ivanov, O.A.; Sukhanov, V.V. Species structure of pelagic ichthyocenes in Russian waters of Far Eastern seas and the Pacific Ocean in 1980–2009. J. Ichthyol. 2015, 55, 497–526. [Google Scholar] [CrossRef]
- Orlov, A.M.; Tokranov, A.M. Checklist of deep-sea fishes of the Russian northwestern Pacific Ocean found at depths below 1000 m. Prog. Oceanogr. 2019, 176, 102143. [Google Scholar] [CrossRef]
- Dolganov, V.N. Feeding of skates of the Rajidae family and their role in ecosystems of the Far Eastern seas. Izv. TINRO 1998, 124, 417–424. [Google Scholar]
- Dolganov, V.N. Distribution and migration of skates of the family Rajidae of the Far Eastern seas of Russia. Izv. TINRO 1998, 124, 433–437. [Google Scholar]
- Dolganov, V.N. Abiotic conditions of the habitat of skates of the family Rajidae of the Far Eastern seas of Russia. Izv. TINRO 1998, 124, 429–432. [Google Scholar]
- Dolganov, V.N. Reproduction of skates of the Rajidae family of the Far Eastern seas of Russia. Izv. TINRO 1998, 124, 425–428. [Google Scholar]
- Dolganov, V.N. Geographical and bathymetric distribution of skates of the Rajidae family in the Far Eastern seas of Russia. Vopr. Ikhtiol. 1999, 39, 428–430. [Google Scholar]
- Dolganov, V.N. The size, age and growth of the skates of family Rajidae of the Far Eastern seas of Russia. Izv. TINRO 2005, 143, 84–89. [Google Scholar]
- Dudnik, Y.I.; Dolganov, V.N. Distribution and stocks of fish on the continental slope of the Sea of Okhotsk and the Kuril Islands in the summer of 1989. Vopr. Ikhtiol. 1992, 32, 83–98. [Google Scholar]
- Fatykhov, R.N.; Poltev, Y.N.; Mukhametov, I.N.; Nemchinov, O.Y. Spatial distribution of mass species of skates of the genus Bathyraja in the area of the northern Kuril Islands and southeastern Kamchatka in various seasons of 1996–1997. In Commercial and Biological Studies of Fish in the Pacific Waters of the Kuril Islands and Adjacent Areas of the Okhotsk and Bering Seas in 1992–1998; VNIRO Publishing: Moscow, Russia, 2000; pp. 104–120. [Google Scholar]
- Glubokov, A.I.; Orlov, A.M. Some morphophysiological indicators and feeding characteristics of the Aleutian skate Bathyraja aleutica from the western Bering Sea. Probl. Fish. 2000, 1, 126–149. [Google Scholar]
- Orlov, A.M. Trophic interrelations in predatory fishes of Pacific waters circumambient the northern Kuril Islands and southeastern Kamchatka. Hydrobiol. J. 2004, 40, 1–19. [Google Scholar] [CrossRef]
- Davis, C.D.; Ebert, D.A.; Ishihara, H.; Orlov, A.; Compagno, L.J.V.; Farrugia, T.J.; Tribuzio, C.A. Bathyraja aleutica. In The IUCN Red List of Threatened Species 2015; 2015; p. e.T161661A80674005. Available online: https://www.iucnredlist.org/species/161661/80674005 (accessed on 25 October 2022).
- Ebert, D.A. Reproductive biology of skates, Bathyraja (Ishiyama), along the eastern Bering Sea continental slope. J. Fish Biol. 2005, 66, 618–649. [Google Scholar] [CrossRef]
- Tokranov, A.M.; Orlov, A.M.; Sheiko, B.A. Commercial Fishes of Continental Slope of Kamchatka Waters; Kamchatpress: Petropavlovsk-Kamchatsky, Russia, 2005; pp. 1–52. [Google Scholar]
- Orlov, A.; Tokranov, A.; Fatykhov, R. Common deep-benthic skates (Rajidae) of the northwestern Pacific: Basic ecological and biological features. Cybium 2006, 30, 49–65. [Google Scholar]
- Spies, I.B.; Gaichas, S.; Stevenson, D.E.; Orr, J.W.; Canino, M.F. DNA-based identification of Alaska skates (Amblyraja, Bathyraja and Raja: Rajidae) using cytochrome c oxidase subunit I (coI) variation. J. Fish Biol. 2006, 69, 283–292. [Google Scholar] [CrossRef]
- Spies, I.B.; Orr, J.W.; Stevenson, D.E.; Goddard, P.; Hoff, G.; Guthridge, J.; Hollowed, M.; Rooper, C. Skate egg nursery areas support genetic diversity of Alaska and Aleutian skates in the Bering Sea. Mar. Ecol. Progr. Ser. 2021, 669, 121–138. [Google Scholar] [CrossRef]
- Ebert, D.A.; Bizzarro, J.J. Standardized diet compositions and trophic levels of skates (Chondrichthyes: Rajiformes: Rajoidei). Environ. Biol. Fish. 2007, 80, 221–237. [Google Scholar] [CrossRef]
- Yang, M.-S. Food habits and diet overlap of seven skate species in the Aleutian Islands. U.S. Dep. Commer. NOAA Tech. Memo. NMFS-AFSC 2007, 177, 1–46. [Google Scholar]
- Stevenson, D.E.; Orr, J.W.; Hoff, G.R.; McEachran, J.D. Emerging patterns of species richness, diversity, population density, and distribution in the skates (Rajidae) of Alaska. Fish. Bull. 2007, 106, 24–39. [Google Scholar]
- Stevenson, D.E.; Orr, J.W.; Hoff, G.R.; McEachran, J.D. Sharks, Skates and Ratfish of Alaska; University of Alaska Sea Grant: Fairbanks, AK, USA, 2007; pp. 1–85. [Google Scholar]
- Hoff, G.R. Embryo developmental events and the egg case of the Aleutian skate Bathyraja aleutica (Gilbert) and the Alaska skate Bathyraja parmifera (Bean). J. Fish Biol. 2009, 74, 483–501. [Google Scholar] [CrossRef]
- Hoff, G.R. Skate Bathyraja spp. egg predation in the eastern Bering Sea. J. Fish Biol. 2009, 74, 250–269. [Google Scholar] [CrossRef]
- Hoff, G.R. Identification of skate nursery habitat in the eastern Bering Sea. Mar. Ecol. Prog. Ser. 2010, 403, 243–254. [Google Scholar] [CrossRef]
- Hoff, G.R. Identification of multiple nursery habitats of skates in the eastern Bering Sea. J. Fish Biol. 2016, 88, 1746–1757. [Google Scholar] [CrossRef] [PubMed]
- Orlov, A.; Binohlan, C. Length-weight relationships of deep-sea fishes from the western Bering Sea. J. Appl. Ichthyol. 2009, 25, 223–227. [Google Scholar] [CrossRef]
- Brown, S.C.; Bizzarro, J.J.; Cailliet, G.M.; Ebert, D.A. Breaking with tradition: Redefining measures for diet description with a case study of the Aleutian skate Bathyraja aleutica (Gilbert 1896). Environ. Biol. Fish. 2012, 95, 3–20. [Google Scholar] [CrossRef]
- Teshima, K.; Tomonaga, S. Reproduction of Aleutian skate, Bathyraja aleutica, with comments on embryonic development. In Indo-Pacific Fish Biology, Proceedings of the Second International Conference on Indo-Pacific Fishes, Conducted at the Tokyo National Museum, Ueno Park, Tokyo, Japan, 29 July–3 August 1985; Tokyo National Museum: Tokyo, Japan, 1986; pp. 303–309. [Google Scholar]
- Terentyev, D.A.; Zolotov, A.O. Fishing and long-term dynamics of biomass of skates (Bathyraja) off the western coast of Kamchatka. Izv. TINRO 2012, 169, 32–40. [Google Scholar]
- Haas, D.L.; Ebert, D.A.; Cailliet, G.M. Comparative age and growth of the Aleutian skate, Bathyraja aleutica, from the eastern Bering Sea and Gulf of Alaska. Environ. Biol. Fish. 2016, 99, 813–828. [Google Scholar] [CrossRef]
- Vinogradskaya, A.V.; Matveev, A.A.; Ryazanova, T.V.; Terentyev, D.A.; Kurbanov, Y.K. Methods of visualization of annual rings on the vertebrae of some species of rhombic rays (Rajidae Blainville, 1816). Bull. Kamchat. Stat. Tech. Univ. 2019, 49, 89–97. [Google Scholar] [CrossRef]
- Vinogradskaya, A.V.; Matveev, A.A.; Terentyev, D.A. New data on fishing and the state of stocks of skates of the family Arhynchobatidae off the western coast of Kamchatka. Bull. Kamchat. Stat. Tech. Univ. 2022, 59, 49–61. [Google Scholar] [CrossRef]
- Rooper, C.N.; Hoff, G.R.; Stevenson, D.E.; Orr, J.W.; Spies, I.B. Skate egg nursery habitat in the eastern Bering Sea: A predictive model. Mar. Ecol. Prog. Ser. 2019, 609, 163–178. [Google Scholar] [CrossRef]
- Orlov, A.M. Trophic relationships of commercial fishes in the Pacific waters off southeastern Kamchatka and the northern Kuril Islands. In Ecosystem Approaches for Fisheries Management; Alaska Sea Grant College Program: Fairbanks, AK, USA, 1999; pp. 231–263. [Google Scholar]
- Orlov, A.M. Quantitative Distribution of Demersal Nekton in the Pacific Waters of the Northern Kuril Islands and Southeastern Kamchatka; VNIRO Publishing: Moscow, Russia, 2010; pp. 1–335. [Google Scholar]
- Stevenson, D.E. Identification of skates, sculpins, and smelts by observers in north Pacific groundfish fisheries (2002–2003). U.S. Dep. Commer. Tech. Memo. NMFS-AFSC 2004, 142, 1–67. [Google Scholar]
- Dolganov, V.N.; Tuponogov, V.N. Keys to Skates of Genus Bathyraja and Rhinoraja (Fam. Rajidae) of the Far Eastern Seas of Russia. Izv. TINRO 1999, 126, 657–664. [Google Scholar]
- Zotina, R.S.; Zotin, A.I. Quantitative ratio between weight, length, egg sizes, and fecundity of the animals. Zh. Obshch. Biol. 1967, 28, 82–92. [Google Scholar] [PubMed]
- Krivobok, M.N.; Shatunovsky, M.I. Analysis methods of fishes using morphophysiological parameters. In Methods of Morphophysiological and Biochemical Studies of Fishes; VNIRO: Moscow, Russia, 1972; pp. 29–44. [Google Scholar]
- Nikol’sky, G.V. Ecology of Fishes; Vysshaya Shkola: Moscow, Russia, 1974; pp. 1–367. [Google Scholar]
- Orlov, A.M.; Baitalyuk, A.A. Spatial distribution and features of biology of Pacific sleeper shark Somniosus pacificus in the North Pacific. J. Ichthyol. 2014, 54, 526–546. [Google Scholar] [CrossRef]
- Grigorov, I.V.; Orlov, A.M.; Baitalyuk, A.A. Spatial distribution, size composition, feeding habits, and dynamics of abundance of Alaska skate Bathyraja parmifera in the North Pacific. J. Ichthyol. 2015, 55, 644–663. [Google Scholar] [CrossRef]
- Grigorov, I.V.; Baitalyuk, A.A.; Orlov, A.M. Spatial distribution, size composition, and dynamics of catches of the Okhotsk skate Bathyraja violacea in the North Pacific Ocean. J. Ichthyol. 2017, 57, 706–720. [Google Scholar] [CrossRef]
- Ormseth, O.A. Partial Assessment of the Skate Stock Complex in the Bering Sea and Aleutian Islands. In NPFMC Bering Sea and Aleutian Islands SAFE; North Pacific Fisheries Management Council: Anchorage, AK, USA, 2021; pp. 1–9. Available online: https://www.fisheries.noaa.gov/resource/data/2021-assessment-skate-stock-complex-bering-sea-and-aleutian-islands (accessed on 3 October 2022).
- Ormseth, O.A. Assessment of the skate stock complex in the Gulf of Alaska. In NPFMC Gulf of Alaska SAFE; North Pacific Fisheries Management Council: Anchorage, AK, USA, 2021; pp. 1–60. Available online: https://www.fisheries.noaa.gov/resource/data/2021-assessment-skate-stock-complex-gulf-alaska (accessed on 3 October 2022).
- Antonov, N.P.; Klovatch, N.V.; Orlov, A.M.; Datsky, A.V.; Lepskaya, V.A.; Kuznetsov, V.V.; Yarzhombek, A.A.; Abramov, A.A.; Alekseev, D.O.; Moiseev, S.I.; et al. Fishing in the Russian Far East fishery basin in 2013. Tr. VNIRO 2013, 160, 133–211. [Google Scholar]
- Badaev, O.Z. Estimation of bycatch and discards of longline fish in Far East seas. Probl. Fish. 2018, 19, 58–72. [Google Scholar]
- Zolotov, A.O. Modern specialized fishery of sea fish in the western Bering Sea. Izv. TINRO 2021, 201, 76–101. [Google Scholar] [CrossRef]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Type | No. Records | Data Source/ Organization | Link to Data Source |
|---|---|---|---|
| Scientific bottom-trawl surveys | 3356 | Pacific branch of Russian Federal Research Institute of Fisheries and Oceanography (TINRO), Vladivostok, Russia | The data received upon request on 1 October 2022 |
| Scientific bottom-trawl surveys | 3010 | Alaska Fisheries Science Center (AFSC), National Oceanic and Atmospheric Administration (NOAA), Seattle, WA, USA | https://www.fisheries.noaa.gov/alaska/commercial-fishing/alaska-groundfish-bottom-trawl-survey-data accessed on 3 October 2022 |
| Scientific bottom-trawl surveys | 47 | Northwestern Fisheries Science Center National Oceanic and Atmospheric Administration—NWFSC NOAA, Seattle, WA, USA. | https://www.webapps.nwfsc.noaa.gov/data/map accessed on 3 October 2022 |
| Observations on commercial fishing vessels | 8567 | Alaska Fisheries Science Center (AFSC), National Oceanic and Atmospheric Administration (NOAA), Seattle, WA, USA | https://www.fisheries.noaa.gov/resource/map/spatial-data-collected-groundfish-observers-alaska accessed on 3 October 2022 |
| Museum collection | 40 | Fish Collection of Hokkaido University—National Museum of Nature and Science, Hokkaido University, Sapporo, Hokkaido, Japan | https://www.gbif.org/ru/occurrence/search?dataset_key=848ae956-f762-11e1-a439-00145eb45e9a&taxon_key=2420280 accessed via GBIF.org on 3 October 2022 |
| Institution collection | 18 | Ichthyology collection of the California Academy of Sciences (CAS), San Francisco, CA, USA | https://www.gbif.org/ru/occurrence/search?publishing_org=66522820-055c-11d8-b84e-b8a03c50a862&taxon_key=2420280 accessed via GBIF.org on 3 October 2022 |
| Scientific bottom-trawl surveys | 1 | Pacific Biological Station (PBS), Fisheries and Oceans Canada, Nanaimo, BC, Canada | https://www.gbif.org/ru/occurrence/search?dataset_key=310dbb60-e369-4ec2-a6d8-8e4de843b208&publishing_org=c98a8d0d-cab2-4506-8c0f-c91a5e4996b9&taxon_key=2420280 accessed via GBIF.org on 3 October 2022 |
| Scientific bottom-trawl surveys | 128 | Pacific Biological Station (PBS), Fisheries and Oceans Canada, Nanaimo, BC, Canada | https://www.gbif.org/ru/occurrence/search?dataset_key=9fcd8de5-a2b6-4c72-b863-debe63ee3a72&publishing_org=c98a8d0d-cab2-4506-8c0f-c91a5e4996b9&taxon_key=2420280 accessed via GBIF.org on 3 October 2022 |
| Scientific bottom-trawl surveys | 44 | Pacific Biological Station (PBS), Fisheries and Oceans Canada, Nanaimo, BC, Canada | https://www.gbif.org/ru/occurrence/search?dataset_key=48060741-47d3-4093-86e6-2ec484323cc5&publishing_org=c98a8d0d-cab2-4506-8c0f-c91a5e4996b9&taxon_key=2420280 accessed via GBIF.org on 3 October 2022 |
| Scientific bottom-trawl surveys | 46 | Pacific Biological Station (PBS), Fisheries and Oceans Canada, Nanaimo, BC, Canada | https://www.gbif.org/ru/occurrence/search?dataset_key=3d4cac0a-7441-4099-8eed-8a270d522f8f&publishing_org=c98a8d0d-cab2-4506-8c0f-c91a5e4996b9&taxon_key=2420280 accessed via GBIF.org on 3 October 2022 |
| Scientific bottom-trawl surveys | 4 | Pacific Biological Station (PBS), Fisheries and Oceans Canada, Nanaimo, BC, Canada | https://www.gbif.org/ru/occurrence/search?dataset_key=f2189339-9098-4e0e-98d4-9c72e5e5f96d&publishing_org=c98a8d0d-cab2-4506-8c0f-c91a5e4996b9&taxon_key=2420280 accessed via GBIF.org on 3 October 2022 |
| Scientific bottom-longline surveys | 17 | Pacific Biological Station (PBS), Fisheries and Oceans Canada, Nanaimo, BC, Canada | https://www.gbif.org/ru/occurrence/search?dataset_key=1677f287-3666-49b8-90f2-12a63ca58438&publishing_org=c98a8d0d-cab2-4506-8c0f-c91a5e4996b9&taxon_key=2420280 accessed via GBIF.org on 3 October 2022 |
| Scientific bottom-longline surveys | 7 | Pacific Biological Station (PBS), Fisheries and Oceans Canada, Nanaimo, BC, Canada | https://www.gbif.org/ru/occurrence/search?dataset_key=e908605e-fb40-4db1-bff7-de1c241284a9&publishing_org=c98a8d0d-cab2-4506-8c0f-c91a5e4996b9&taxon_key=2420280 accessed via GBIF.org on 3 October 2022 |
| Museum collection | 2 | Vertebrate, zoology, fishes collections of Smithsonian National Museum of Natural History—NMNH, Washington, DC, USA. | https://collections.nmnh.si.edu/search/fishes/ accessed on 3 October 2022 |
| Museum collection | 6 | Fish collection of the Canadian Museum of Nature, Ottawa, ON, Canada | http://ipt.nature.ca/resource?r=cmn_fish&v=1.124 accessed on 3 October 2022 |
| Museum collection | 66 | Ichthyology collection of Burke Museum of Natural History and Culture (BMNHC), Seattle, WA, USA | https://www.burkemuseum.org/collections-and-research/biology/ichthyology/collections-database/results.php?l=100&o=0&f=&g=&s=17h~IymyNd1yIp&d=gpA&w=PhIyNl1XhPYMY0j1yI1Kp0YndhYPhSh|b3NhsPYMY0jld1B0&wo=PhIyNl1XhPYjXhlE1Yj1yI1KpYndhYPhSh|b3NhsPYjXhlE1Yjld1B accessed on 3 October 2022 |
| Museum collection | 2 | Ichthyology collection of Muséum national d’Histoire naturelle (MNHN), Paris, France | https://obis.org/taxon/271506 accessed via OBIS.org on 3 October 2022 |
| Institution collection | 1 | Ichthyology collection University of Kansas Biodiversity Institute & Natural History Museum (KUBI), Lawrence, KS, USA | https://obis.org/taxon/271506 accessed via OBIS.org on 3 October 2022 |
| Museum collection | 1 | Fish collection of the National Museum of Nature and Science (NMNS), Tokyo, Japan | https://obis.org/taxon/271506 accessed via OBIS.org on 3 October 2022 |
| Institution collection | 12 | Texas Cooperative Wildlife Collection (TCWC), Department of Wildlife and Fisheries Sciences, Texas A&M University, College Station, TX, USA | https://www.gbif.org/ru/occurrence/search?publishing_org=2b8e48b0-812d-11de-86fe-b8a03c50a862&taxon_key=2420280 accessed via GBIF.org on 3 October 2022 |
| Institution collection | 1 | Marine Vertebrate Collection of the Scripps Institution of Oceanography (SIO), San Diego, CA, USA | https://sioapps.ucsd.edu/collections/mv/collection/94-205/?q=Bathyraja+aleutica accessed on 3 October 2022 |
| Museum collection | 1 | Fish collection of the Museum of the North, University of Alaska (UAM), Fairbanks, AK, USA | http://arctos.database.museum/guid/UAM:Fish:9455?seid=2666501 accessed on 3 October 2022 |
| Total | 15,377 | 22 sources/15 organizations | |
| Sex | Area | Parameters of LWR | n | TL, cm | Weight, kg | Source | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| a | b | R2 | Min–Max | M ± SE | Min–Max | M ± SE | ||||
| Males | NWPO | 3.17 × 10−6 | 3.12 | 0.99 | 83 | 20–145 | 64.34 ± 2.90 | 0.04–9.10 | 2.13 ± 0.27 | Our data |
| WBS | 4.58 × 10−6 | 3.06 | 0.97 | 213 | 11–135 | 67.30 ± 1.60 | 0.06–13.98 | 4.35 ± 0.26 | Our data | |
| 2.81 × 10−6 | 3.15 | 0.99 | 74 | 22–130 | - | - | - | [56] | ||
| Okh | 4.41 × 10−6 | 3.04 | 0.97 | 50 | 20–146 | 90.51 ± 4.96 | 0.12–18.20 | 5.92 ± 0.79 | Our data | |
| BC | 3.51 × 10−6 | 3.11 | 0.94 | 28 | 59–137 | 88.36 ± 4.06 | 1.02–15.52 | 4.75 ± 0.70 | Our data | |
| Females | NWPO | 2.74 × 10−6 | 3.17 | 0.99 | 145 | 24–115 | 64.83 ± 3.03 | 0.04–17.60 | 2.38 ± 0.26 | Our data |
| WBS | 2.83 × 10−6 | 3.16 | 0.96 | 207 | 13–143 | 66.28 ± 1.25 | 0.07–18.40 | 3.45 ± 0.26 | Our data | |
| 1.55 × 10−6 | 3.28 | 0.99 | 69 | 28–134 | - | - | - | [56] | ||
| Okh | 3.23 × 10−6 | 3.12 | 0.98 | 61 | 27–153 | 85.91 ± 3.78 | 0.08–21.55 | 5.48 ± 0.75 | Our data | |
| BC | 1.31 × 10−5 | 2.82 | 0.90 | 42 | 52–140 | 98.26 ± 3.39 | 0.82–17.15 | 6.35 ± 0.60 | Our data | |
| Both sexes | NWPO | 2.61 × 10−6 | 3.20 | 0.96 | 356 | 20–145 | 74.17 ± 1.54 | 0.04–18.20 | 3.89 ± 0.21 | Our data |
| 3.03 × 10−6 | 3.16 | - | 507 | 20–128 | 82.40 ± 0.93 | 0.06–14.10 | 4.42 ± 0.30 | [45] | ||
| WBS | 2.79 × 10−6 | 3.16 | 0.95 | 783 | 10–148 | 71.92 ± 0.55 | 0.04–31 | 4.16 ± 0.15 | Our data | |
| 2.13 × 10−6 | 3.21 | 0.99 | 143 | 22–134 | - | - | - | [56] | ||
| Okh | 5.51 × 10−6 | 3.03 | 0.93 | 270 | 20–170 | 96.31 ± 1.73 | 0.08–35 | 8.21 ± 0.44 | Our data | |
| BC | 7.07 × 10−6 | 2.95 | 0.92 | 71 | 10–140 | 96.83 ± 2.42 | 0.82–17.15 | 5.81 ± 0.47 | Our data | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grigorov, I.V.; Kivva, K.K.; Orlov, A.M. The Aleutians and Beyond: Distribution, Size Composition, and Catch Dynamics of the Aleutian Skate Bathyraja aleutica across the North Pacific. Animals 2022, 12, 3507. https://doi.org/10.3390/ani12243507
Grigorov IV, Kivva KK, Orlov AM. The Aleutians and Beyond: Distribution, Size Composition, and Catch Dynamics of the Aleutian Skate Bathyraja aleutica across the North Pacific. Animals. 2022; 12(24):3507. https://doi.org/10.3390/ani12243507
Chicago/Turabian StyleGrigorov, Igor V., Kirill K. Kivva, and Alexei M. Orlov. 2022. "The Aleutians and Beyond: Distribution, Size Composition, and Catch Dynamics of the Aleutian Skate Bathyraja aleutica across the North Pacific" Animals 12, no. 24: 3507. https://doi.org/10.3390/ani12243507
APA StyleGrigorov, I. V., Kivva, K. K., & Orlov, A. M. (2022). The Aleutians and Beyond: Distribution, Size Composition, and Catch Dynamics of the Aleutian Skate Bathyraja aleutica across the North Pacific. Animals, 12(24), 3507. https://doi.org/10.3390/ani12243507