

Metabolic Hepatic Disorders Caused by Ciguatoxins in Goldfish (Carassius auratus)

 ,
,  , and
, and 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish, Experimental Design, and Animal Welfare
2.2. Hepatosomatic Index (HSI)
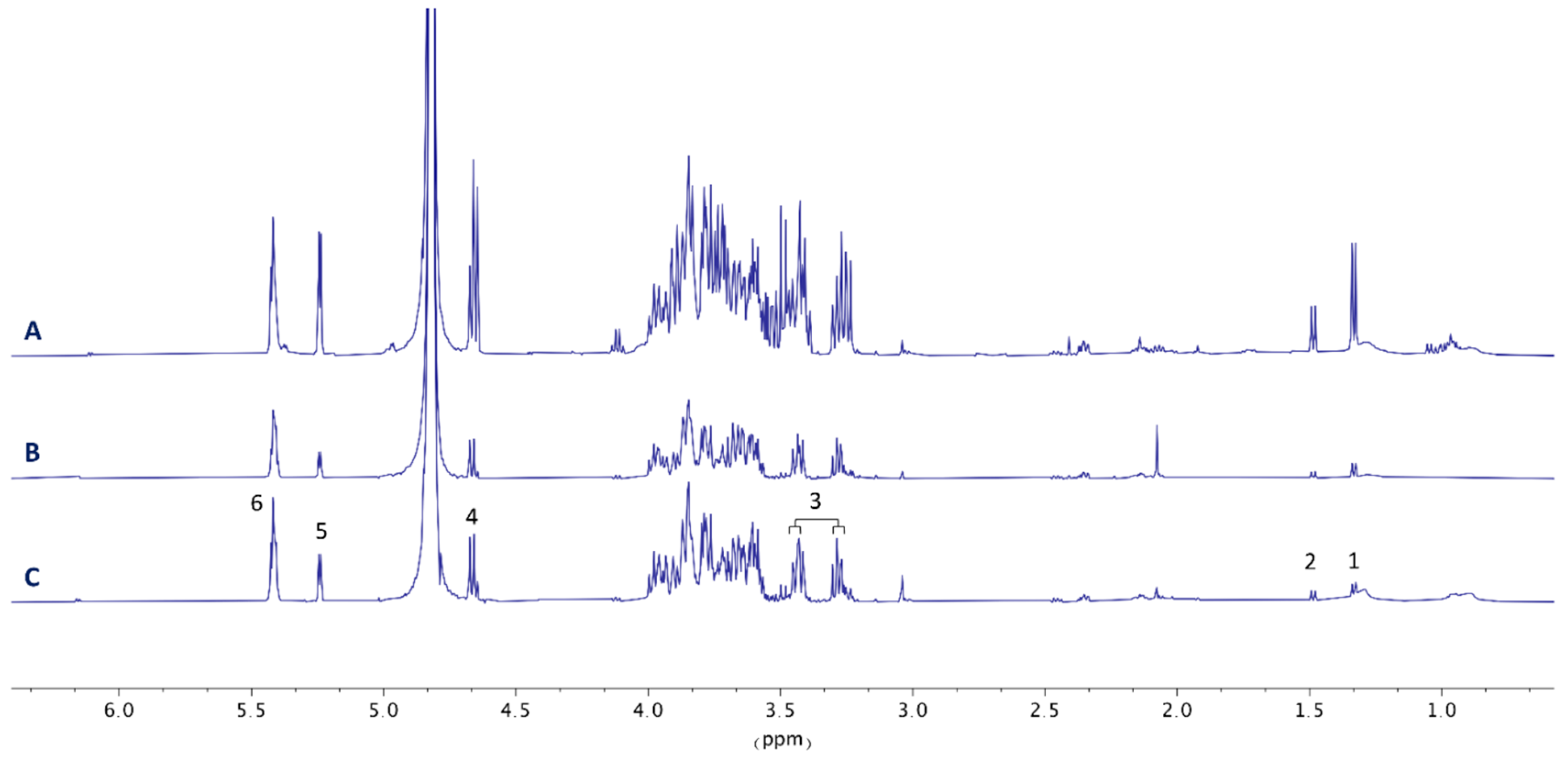
2.3. Sample Preparation and NMR Spectroscopy
2.4. Metabolomics Correlation and Pathway Analysis
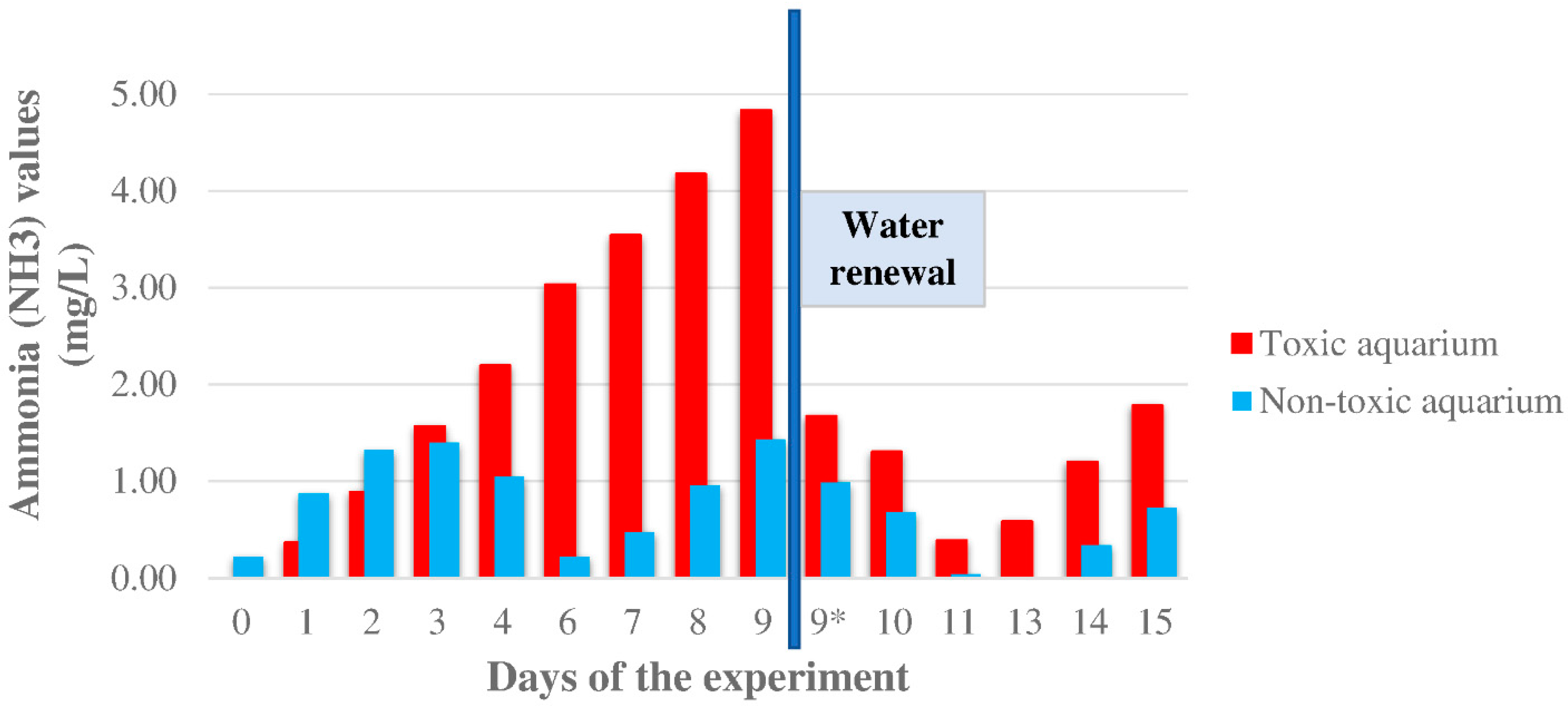
2.5. Determination of the Ammonia Level Variations
2.6. Statistical Analysis
3. Results
3.1. Hepatosomatic Index (HSI) Variations
3.2. Metabolic Profiles of Liver Samples of Carassius auratus
3.3. Ammonia Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chinain, M.; Gatti, C.M.I.; Darius, H.T.; Quod, J.P.; Tester, P.A. Ciguatera poisonings: A global review of occurrences and trends. Harmful Algae 2021, 102, 22. [Google Scholar] [CrossRef] [PubMed]
- FAO; WHO. Food Safety and Quality No.9. In Report of the Expert Meeting on Ciguatera Poisoning: Rome, 19–23 November 2018; World Health Organization: Rome, Italy, 2020. [Google Scholar]
- Sanchez-Henao, J.A.; Garcia-Alvarez, N.; Fernandez, A.; Saavedra, P.; Sergent, F.S.; Padilla, D.; Acosta-Hernandez, B.; Suarez, M.M.; Diogene, J.; Real, F. Predictive score and probability of CTX-like toxicity in fish samples from the official control of ciguatera in the Canary Islands. Sci. Total Environ. 2019, 673, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Henao, A.; Garcia-Alvarez, N.; Sergent, F.S.; Estevez, P.; Gago-Martinez, A.; Martin, F.; Ramos-Sosa, M.; Fernandez, A.; Diogene, J.; Real, F. Presence of CTXs in moray eels and dusky groupers in the marine environment of the Canary Islands. Aquat. Toxicol. 2020, 221, 9. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.R.; Estevez, P.; Solino, L.; Castro, D.; Rodrigues, S.M.; Timoteo, V.; Leao-Martins, J.M.; Santos, C.; Gouveia, N.; Diogene, J.; et al. An Update on Ciguatoxins and CTX-like Toxicity in Fish from Different Trophic Levels of the Selvagens Islands (NE Atlantic, Madeira, Portugal). Toxins 2021, 13, 580. [Google Scholar] [CrossRef] [PubMed]
- Boada, L.D.; Zumbado, M.; Luzardo, O.R.; Almeida-Gonzalez, M.; Plakas, S.M.; Granade, H.R.; Abraham, A.; Jester, E.L.E.; Dickey, R.W. Ciguatera fish poisoning on the West Africa Coast: An emerging risk in the Canary Islands (Spain). Toxicon 2010, 56, 1516–1519. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.W.; Li, X.M.; Lam, P.K.S.; Cheng, S.H.; Schlenk, D.; de Mitcheson, Y.S.; Li, Y.; Gu, J.D.; Chan, L.L. Proteomic analysis of hepatic tissue of ciguatoxin (CTX) contaminated coral reef fish Cephalopholis argus and moray eel Gymnothorax undulatus. Harmful Algae 2012, 13, 65–71. [Google Scholar] [CrossRef]
- Ramos-Sosa, M.J.; García-Álvarez, N.; Sanchez-Henao, A.; Silva Sergent, F.; Padilla, D.; Estévez, P.; Caballero, M.J.; Martín-Barrasa, J.L.; Gago-Martínez, A.; Diogène, J.; et al. Ciguatoxin Detection in Flesh and Liver of Relevant Fish Species from the Canary Islands. Toxins 2022, 14, 46. [Google Scholar] [CrossRef]
- Morey, J.S.; Ryan, J.C.; Dechraoui, M.Y.B.; Rezvani, A.H.; Levin, E.D.; Gordon, C.J.; Ramsdell, J.S.; Van Dolah, F.M. Liver genomic responses to ciguatoxin: Evidence for activation of phase I and phase II detoxification pathways following an acute hypothermic response in mice. Toxicol. Sci. 2008, 103, 298–310. [Google Scholar] [CrossRef]
- Gwinn, J.K.; Uhlig, S.; Ivanova, L.; Faeste, C.K.; Kryuchkov, F.; Robertson, A. In Vitro Glucuronidation of Caribbean Ciguatoxins in Fish: First Report of Conjugative Ciguatoxin Metabolites. Chem. Res. Toxicol. 2021, 24, 1910–1925. [Google Scholar] [CrossRef]
- Bonvallot, N.; Tremblay-Franco, M.; Chevrier, C.; Canlet, C.; Debrauwer, L.; Cravedi, J.P.; Cordier, S. Potential Input From Metabolomics for Exploring and Understanding the Links Between Environment and Health. J. Toxicol. Env. Health-Pt b-Crit. Rev. 2014, 17, 21–44. [Google Scholar] [CrossRef]
- Bennett, C.T.; Robertson, A. Depuration Kinetics and Growth Dilution of Caribbean Ciguatoxin in the Omnivore Lagodon rhomboides: Implications for Trophic Transfer and Ciguatera Risk. Toxins 2021, 13, 774. [Google Scholar] [CrossRef]
- Leite, I.D.; Sdiri, K.; Taylor, A.; Viallon, J.; Gharbia, H.B.; Junior, L.L.M.; Swarzenski, P.; Oberhaensli, F.; Darius, H.T.; Chinain, M.; et al. Experimental Evidence of Ciguatoxin Accumulation and Depuration in Carnivorous Lionfish. Toxins 2021, 13, 564. [Google Scholar] [CrossRef]
- Emwas, A.H.; Roy, R.; McKay, R.T.; Tenori, L.; Saccenti, E.; Gowda, G.A.N.; Raftery, D.; Alahmari, F.; Jaremko, L.; Jaremko, M.; et al. NMR Spectroscopy for Metabolomics Research. Metabolites 2019, 9, 123. [Google Scholar] [CrossRef]
- Blanco, A.M.; Sundarrajan, L.; Bertucci, J.I.; Unniappan, S. Why goldfish? Merits and challenges in employing goldfish as a model organism in comparative endocrinology research. Gen. Comp. Endocrinol. 2018, 257, 13–28. [Google Scholar] [CrossRef]
- Wilkie, M.P.; Pamenter, M.E.; Duquette, S.; Dhiyebi, H.; Sangha, N.; Skelton, G.; Smith, M.D.; Buck, L.T. The relationship between NMDA receptor function and the high ammonia tolerance of anoxia-tolerant goldfish. J. Exp. Biol. 2011, 214, 4107–4120. [Google Scholar] [CrossRef]
- Li, M.H.; Ruan, L.Y.; Liu, Y.; Xu, H.D.; Chen, T.; Fu, Y.H.; Jiang, L.; Wang, J.S. Insight into biological system responses in goldfish (Carassius auratus) to multiple doses of avermectin exposure by integrated H-1 NMR-based metabolomics. Toxicol. Res. 2015, 4, 1374–1388. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, T.; Li, M.H.; Xu, H.D.; Jia, A.Q.; Zhang, J.F.; Wang, J.S. H-1 NMR based metabolomics approach to study the toxic effects of dichlorvos on goldfish (Carassius auratus). Chemosphere 2015, 138, 537–545. [Google Scholar] [CrossRef]
- Xu, H.D.; Wang, J.S.; Li, M.H.; Liu, Y.; Chen, T.; Jia, A.Q. H-1 NMR based metabolomics approach to study the toxic effects of herbicide butachlor on goldfish (Carassius auratus). Aquat. Toxicol. 2015, 159, 69–80. [Google Scholar] [CrossRef]
- Sanchez-Henao, A.; Garcia-Alvarez, N.; Padilla, D.; Ramos-Sosa, M.; Sergent, F.S.; Fernandez, A.; Estevez, P.; Gago-Martinez, A.; Diogene, J.; Real, F. Accumulation of C-CTX1 in Muscle Tissue of Goldfish (Carassius auratus) by Dietary Experience. Animals 2021, 11, 242. [Google Scholar] [CrossRef]
- Liew, H.J.; Sinha, A.K.; Nawata, C.M.; Blust, R.; Wood, C.M.; De Boeck, G. Differential responses in ammonia excretion, sodium fluxes and gill permeability explain different sensitivities to acute high environmental ammonia in three freshwater teleosts. Aquat. Toxicol. 2013, 126, 63–76. [Google Scholar] [CrossRef]
- Bucking, C. A broader look at ammonia production, excretion, and transport in fish: A review of impacts of feeding and the environment. J. Comp. Physiol. B-Biochem. Syst. Environ. Physiol. 2017, 187, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bingol, K.; Li, D.W.; Bruschweiler-Li, L.; Cabrera, O.A.; Megraw, T.; Zhang, F.L.; Bruschweiler, R. Unified and Isomer-Specific NMR Metabolomics Database for the Accurate Analysis of C-13-H-1 HSQC Spectra. ACS Chem. Biol. 2015, 10, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Stipanuk, M.H.; Londono, M.; Lee, J.I.; Hu, M.; Yu, A.F. Enzymes and metabolites of cysteine metabolism in nonhepatic tissues of rats show little response to changes in dietary protein or sulfur amino acid levels. J. Nutr. 2002, 132, 3369–3378. [Google Scholar] [CrossRef] [PubMed]
- Han, X.B.; Patters, A.B.; Chesney, R.W. Transcriptional repression of taurine transporter gene (TauT) by p53 in renal cells. J. Biol. Chem. 2002, 277, 39266–39273. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Patters, A.B.; Jones, D.P.; Zelikovic, I.; Chesney, R.W. The taurine transporter: Mechanisms of regulation. Acta Physiol. 2006, 187, 61–73. [Google Scholar] [CrossRef]
- Lambert, I.H. Regulation of the cellular content of the organic osmolyte taurine in mammalian cells. Neurochem. Res. 2004, 29, 27–63. [Google Scholar] [CrossRef]
- Miyazaki, T.; Matsuzaki, Y. Taurine and liver diseases: A focus on the heterogeneous protective properties of taurine. Amino Acids 2014, 46, 101–110. [Google Scholar] [CrossRef]
- Li, Y.; Hu, Z.T.; Chen, B.; Bu, Q.; Lu, W.J.; Deng, Y.; Zhu, R.M.; Shao, X.; Hou, J.; Zhao, J.X.; et al. Taurine attenuates methamphetamine-induced autophagy and apoptosis in PC12 cells through mTOR signaling pathway. Toxicol. Lett. 2012, 215, 1–7. [Google Scholar] [CrossRef]
- Holecek, M.; Sispera, L. Effects of Arginine Supplementation on Amino Acid Profiles in Blood and Tissues in Fed and Overnight-Fasted Rats. Nutrients 2016, 8, 206. [Google Scholar] [CrossRef]
- Li, Y.H.; Li, F.N.; Wu, L.; Wei, H.K.; Liu, Y.Y.; Li, T.J.; Tan, B.; Kong, X.F.; Yao, K.; Chen, S.; et al. Effects of dietary protein restriction on muscle fiber characteristics and mTORC1 pathway in the skeletal muscle of growing-finishing pigs. J. Anim. Sci. Biotechnol. 2016, 7, 12. [Google Scholar] [CrossRef]
- Holecek, M.; Kovarik, M. Alterations in protein metabolism and amino acid concentrations in rats fed by a high-protein (casein-enriched) diet—Effect of starvation. Food Chem. Toxicol. 2011, 49, 3336–3342. [Google Scholar] [CrossRef]
- Baliou, S.; Kyriakopoulos, A.M.; Goulielmaki, M.; Panayiotidis, M.I.; Spandidos, D.A.; Zoumpourlis, V. Significance of taurine transporter (TauT) in homeostasis and its layers of regulation. Mol. Med. Rep. 2020, 22, 2163–2173. [Google Scholar] [CrossRef]
- Takeuchi, K.; Toyohara, H.; Sakaguchi, M. A hyperosmotic stress-induced mRNA of carp cell encodes Na+- and Cl--dependent high affinity taurine transporter. Biochim. Biophys. Acta-Biomembr. 2000, 1464, 219–230. [Google Scholar] [CrossRef]
- Voss, J.W.; Pedersen, S.F.; Christensen, S.T.; Lambert, I.H. Regulation of the expression and subcellular localization of the taurine transporter TauT in mouse NIH3T3 fibroblasts. Eur. J. Biochem. 2004, 271, 4646–4658. [Google Scholar] [CrossRef]
- Mollerup, J.; Lambert, I.H. Calyculin A modulates the kinetic constants for the Na+-coupled taurine transport in Ehrlich ascites tumour cells. Biochim. Biophys. Acta-Biomembr. 1998, 1371, 335–344. [Google Scholar] [CrossRef]
- Sakai, S.; Tosaka, T.; Tasaka, J.; Hashiguchi, T.; Yoshihama, I. Taurine uptake by glial-cells in the bullfrog sympathetic-ganglia. Neurochem. Int. 1989, 14, 193–198. [Google Scholar] [CrossRef]
- Nicholson, G.M.; Lewis, R.J. Ciguatoxins: Cyclic polyether modulators of voltage-gated Iion channel function. Marine Drugs 2006, 4, 82–118. [Google Scholar] [CrossRef]
- Bañuelos-Vargas, I.; Lopez, L.M.; Perez-Jimenez, A.; Peres, H. Effect of fishmeal replacement by soy protein concentrate with taurine supplementation on hepatic intermediary metabolism and antioxidant status of totoaba juveniles (Totoaba macdonaldi). Comp. Biochem. Physiol. B-Biochem. Mol. Biol. 2014, 170, 18–25. [Google Scholar] [CrossRef]
- de Moura, L.B.; Diogenes, A.F.; Campelo, D.A.V.; de Almeida, F.L.A.; Pousao-Ferreira, P.M.; Furuya, W.M.; Oliva-Teles, A.; Peres, H. Taurine and methionine supplementation as a nutritional strategy for growth promotion of meagre (Argyrosomus regius) fed high plant protein diets. Aquaculture 2018, 497, 389–395. [Google Scholar] [CrossRef]
- Moon, T.W. Glucose intolerance in teleost fish: Fact or fiction? Comp. Biochem. Physiol. B-Biochem. Mol. Biol. 2001, 129, 243–249. [Google Scholar] [CrossRef]
- Enes, P.; Panserat, S.; Kaushik, S.; Oliva-Teles, A. Dietary Carbohydrate Utilization by European Sea Bass (Dicentrarchus labrax L.) and Gilthead Sea Bream (Sparus aurata L.) Juveniles. Rev. Fish. Sci. 2011, 19, 201–215. [Google Scholar] [CrossRef]
- Polakof, S.; Mommsen, T.P.; Soengas, J.L. Glucosensing and glucose homeostasis: From fish to mammals. Comp. Biochem. Physiol. B-Biochem. Mol. Biol. 2011, 160, 123–149. [Google Scholar] [CrossRef] [PubMed]
- Sampath, W.; Rathnayake, R.; Yang, M.X.; Zhang, W.B.; Mai, K.S. Roles of dietary taurine in fish nutrition. Mar. Life Sci. Technol. 2020, 2, 360–375. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, Z.H.; Liu, G.X.; Deng, K.Y.; Yang, M.X.; Pan, M.Z.; Gu, Z.X.; Liu, D.; Zhang, W.B.; Mai, K.S. Synergistic effects of dietary carbohydrate and taurine on growth performance, digestive enzyme activities and glucose metabolism in juvenile turbot Scophthalmus maximus L. Aquaculture 2019, 499, 32–41. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Monier, M.N. Stimulatory effect of dietary taurine on growth performance, digestive enzymes activity, antioxidant capacity, and tolerance of common carp, Cyprinus carpio L., fry to salinity stress. Fish Physiol. Biochem. 2018, 44, 639–649. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Hu, Y.; Ai, Q.H.; Mao, P.; Tian, Q.Q.; Zhong, L.; Xiao, T.Y.; Chu, W.Y. Effect of dietary taurine supplementation on growth performance, digestive enzyme activities and antioxidant status of juvenile black carp (Mylopharyngodon piceus) fed with low fish meal diet. Aquac. Res. 2018, 49, 3187–3195. [Google Scholar] [CrossRef]
- Magalhaes, R.; Martins, N.; Martins, S.; Lopes, T.; Diaz-Rosales, P.; Pousao-Ferreira, P.; Oliva-Teles, A.; Peres, H. Is dietary taurine required for white seabream (Diplodus sargus) juveniles? Aquaculture 2019, 502, 296–302. [Google Scholar] [CrossRef]
- Dashty, M. A quick look at biochemistry: Carbohydrate metabolism. Clin. Biochem. 2013, 46, 1339–1352. [Google Scholar] [CrossRef]
- Chellappa, S.; Huntingford, F.A.; Strang, R.H.C.; Thomson, R.Y. Condition factor and hepatosomatic index as estimates of energy status in male 3-spined stickleback. J. Fish Biol. 1995, 47, 775–787. [Google Scholar] [CrossRef]
- Saleh, H. Fish liver as indicator of aquatic environmental pollution. Bull. Inst. Oceanogr. Fish. 1982, 8, 69–79. [Google Scholar]
- Biagiantirisbourg, S.; Bastide, J. Hepatic perturbations induced by a herbicide (atrazine) in juvenile grey mullet liza-ramada (mugilidae, teleostei)—An ultrastructural-study. Aquat. Toxicol. 1995, 31, 217–229. [Google Scholar] [CrossRef]
- Tan, Q.; Xie, S.; Zhu, X.; Lei, W.; Yang, Y. Effect of dietary carbohydrate sources on growth performance and utilization for gibel carp (Carassius auratus gibelio) and Chinese longsnout catfish (Leiocassis longirostris Gunther). Aquac. Nutr. 2006, 12, 61–70. [Google Scholar] [CrossRef]
- Bacchetta, C.; Rossi, A.; Ale, A.; Campana, M.; Parma, M.J.; Cazenave, J. Combined toxicological effects of pesticides: A fish multi-biomarker approach. Ecol. Indic. 2014, 36, 532–538. [Google Scholar] [CrossRef]
- Diniz, M.S.; Peres, I.; Castro, L.; Freitas, A.C.; Rocha-Santos, T.A.P.; Pereira, R.; Duarte, A.C. Impact of a secondary treated bleached Kraft pulp mill effluent in both sexes of goldfish (Carassius auratus L.). J. Environ. Sci. Health Part A-Toxic/Hazard. Subst. Environ. Eng. 2010, 45, 1858–1865. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hepatosomatic Index (HSI) | |||
|---|---|---|---|
| Diet group | Non-toxic n = 10 | Toxic n = 11 | Detox n = 2 |
| Mean ± SD | 2.605 ± 0.789 | 2.899 ± 0.572 | 4.331 ± 0.599 |
| Median | 2.388 | 2.827 | 4.331 |
| Minimum | 1.627 | 1.985 | 3.907 |
| Maximum | 4.061 | 3.931 | 4.755 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos-Sosa, M.J.; Hernández López, V.; Sanchez-Henao, A.; García-Álvarez, N.; Díaz-Marrero, A.R.; Rosario, I.; Real, F.; Fernández, J.J. Metabolic Hepatic Disorders Caused by Ciguatoxins in Goldfish (Carassius auratus). Animals 2022, 12, 3500. https://doi.org/10.3390/ani12243500
Ramos-Sosa MJ, Hernández López V, Sanchez-Henao A, García-Álvarez N, Díaz-Marrero AR, Rosario I, Real F, Fernández JJ. Metabolic Hepatic Disorders Caused by Ciguatoxins in Goldfish (Carassius auratus). Animals. 2022; 12(24):3500. https://doi.org/10.3390/ani12243500
Chicago/Turabian StyleRamos-Sosa, María José, Víctor Hernández López, Andres Sanchez-Henao, Natalia García-Álvarez, Ana R. Díaz-Marrero, Inmaculada Rosario, Fernando Real, and José J. Fernández. 2022. "Metabolic Hepatic Disorders Caused by Ciguatoxins in Goldfish (Carassius auratus)" Animals 12, no. 24: 3500. https://doi.org/10.3390/ani12243500
APA StyleRamos-Sosa, M. J., Hernández López, V., Sanchez-Henao, A., García-Álvarez, N., Díaz-Marrero, A. R., Rosario, I., Real, F., & Fernández, J. J. (2022). Metabolic Hepatic Disorders Caused by Ciguatoxins in Goldfish (Carassius auratus). Animals, 12(24), 3500. https://doi.org/10.3390/ani12243500