
Stranding of Mesopelagic Fishes in the Canary Islands

 , , , ,
, , , ,  ,
,  and
and 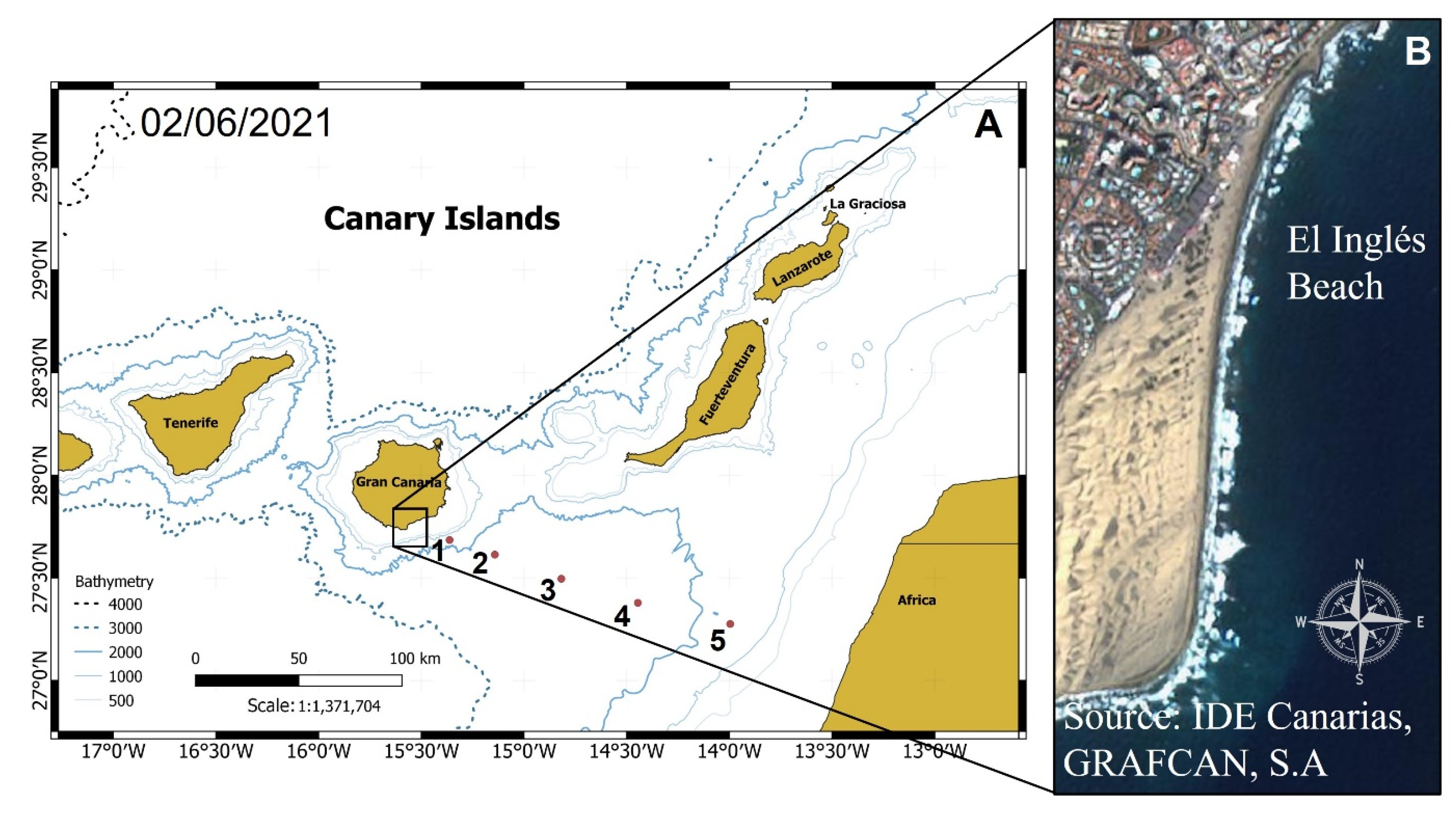{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collecting Samples
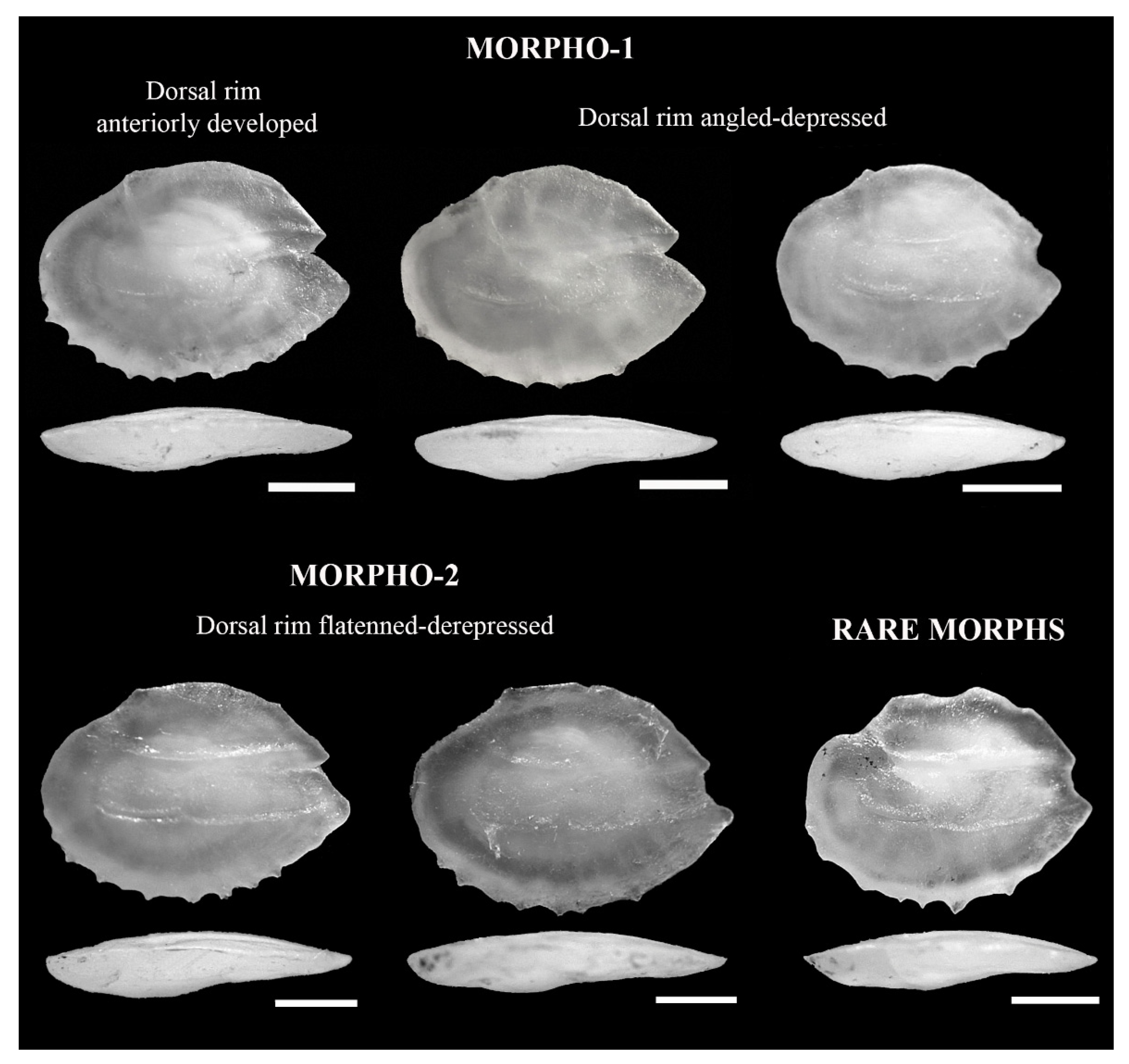
2.2. Classification of Sagittae Otoliths
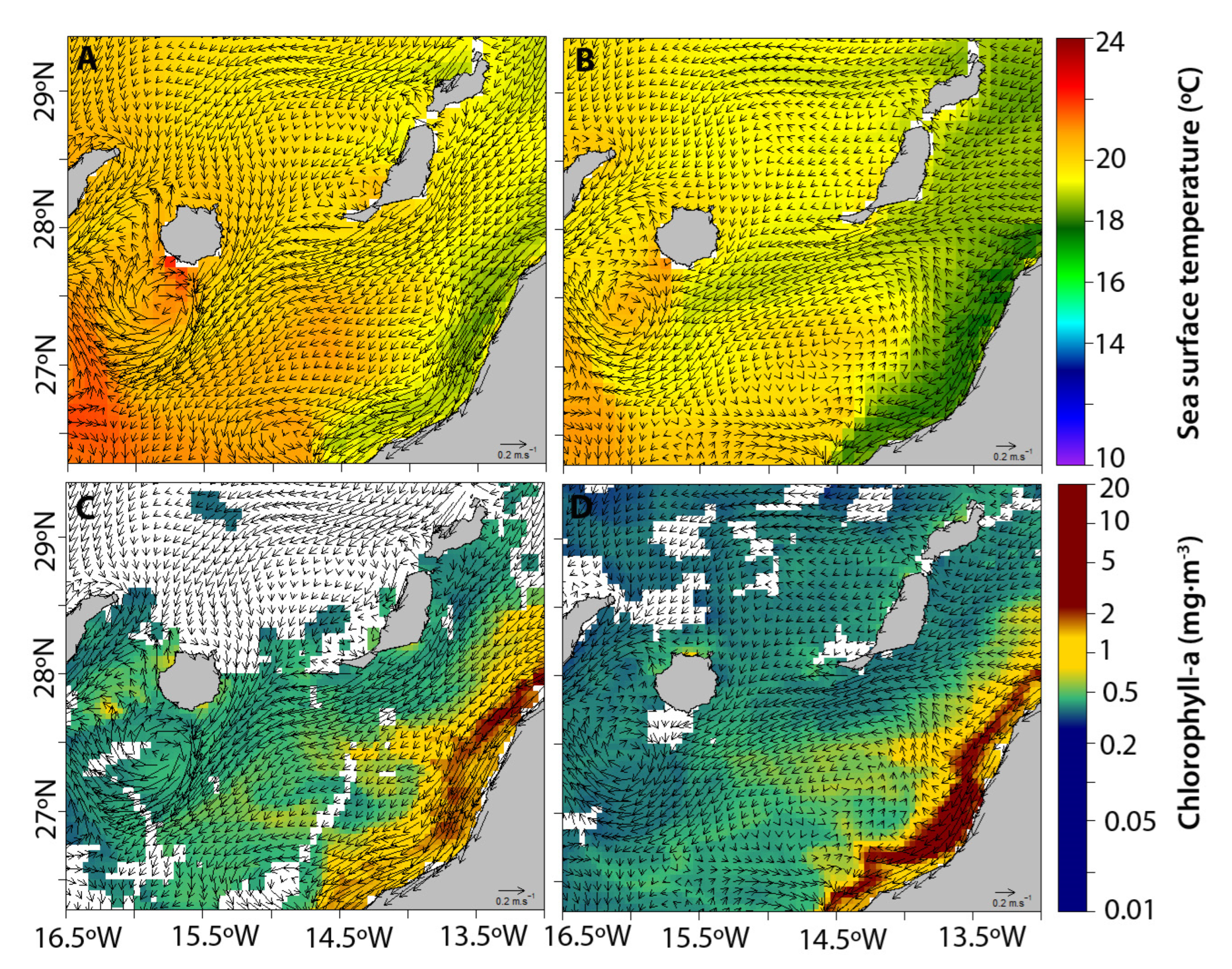
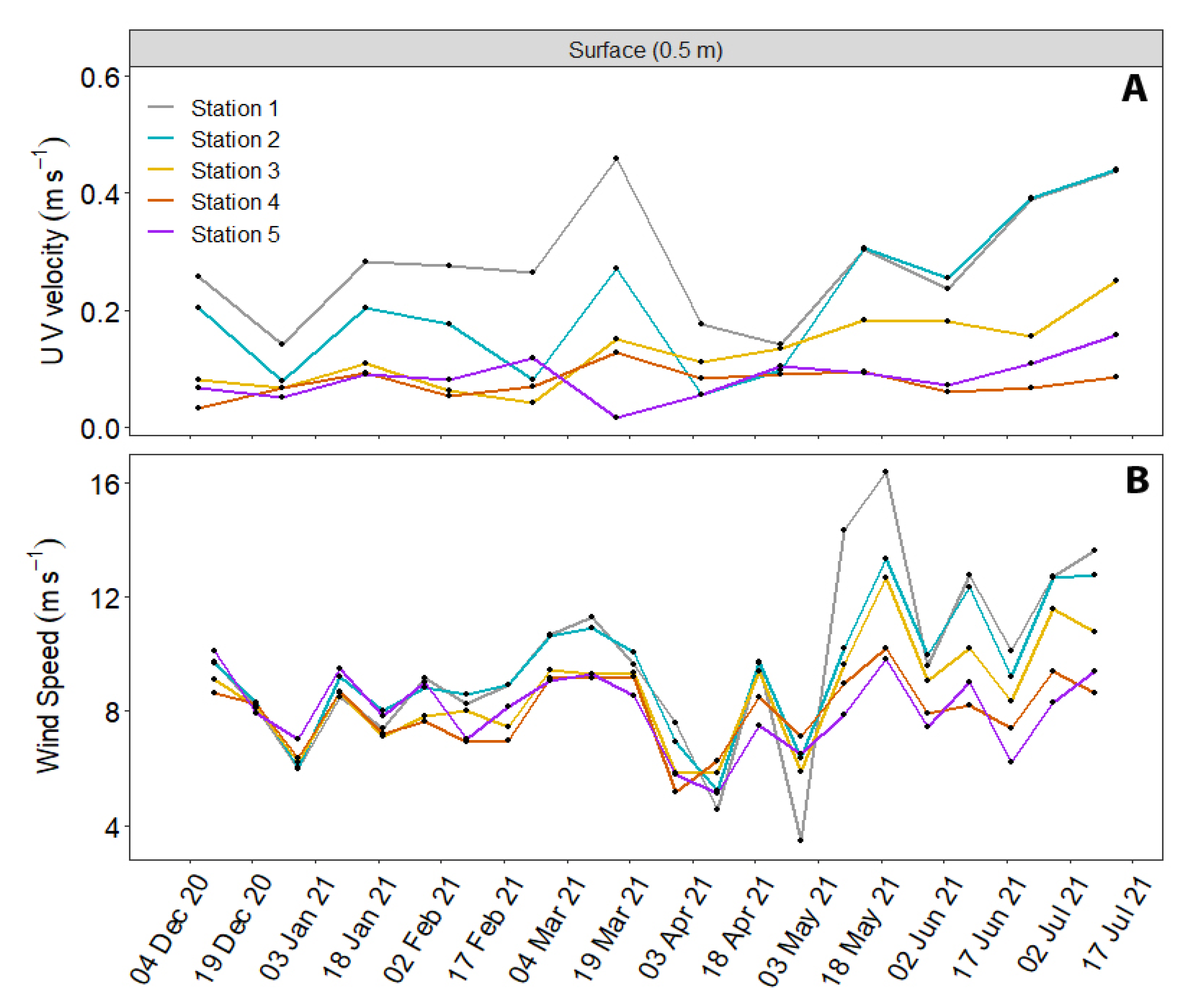
2.3. Oceanographic Characterization
3. Results
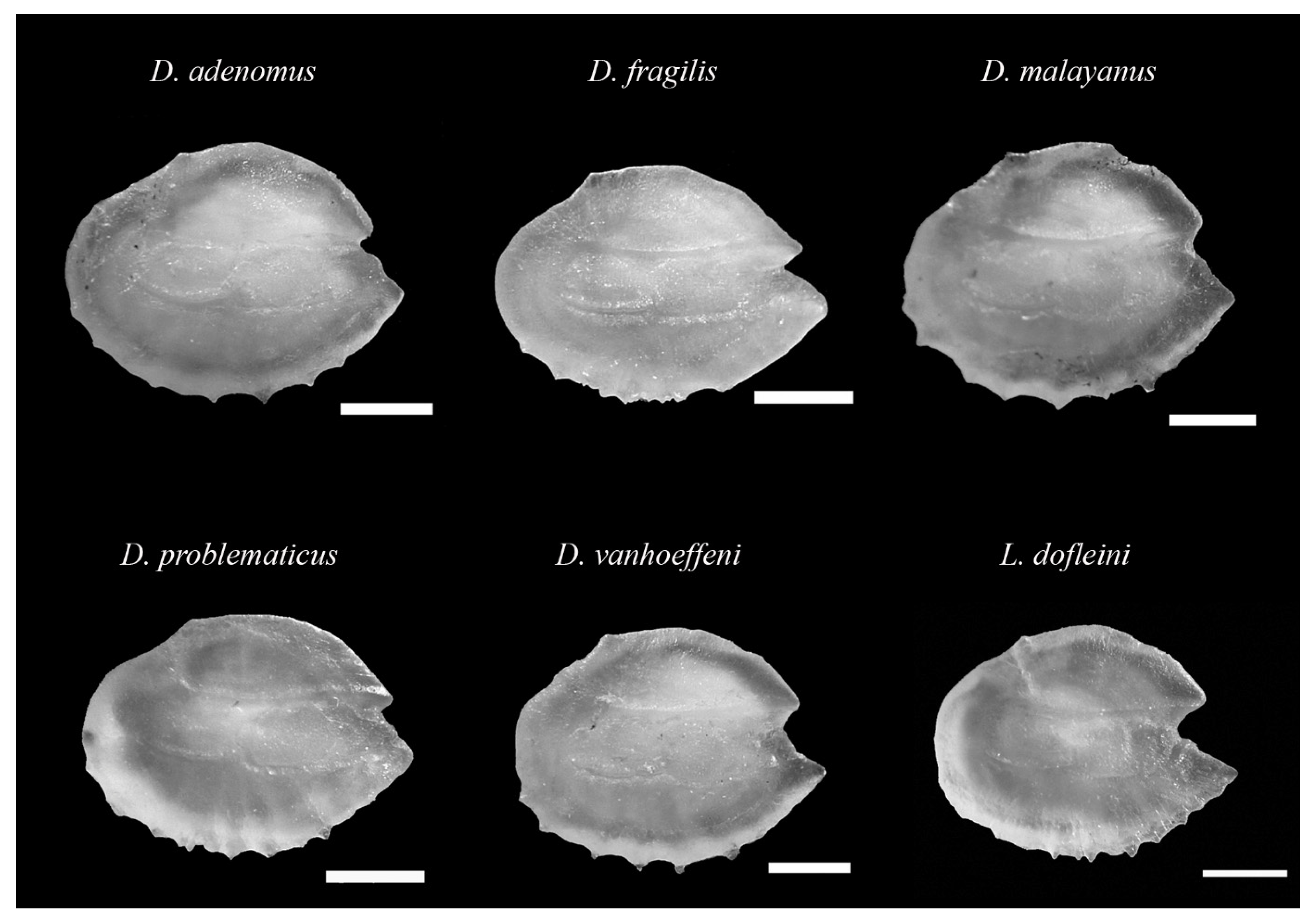
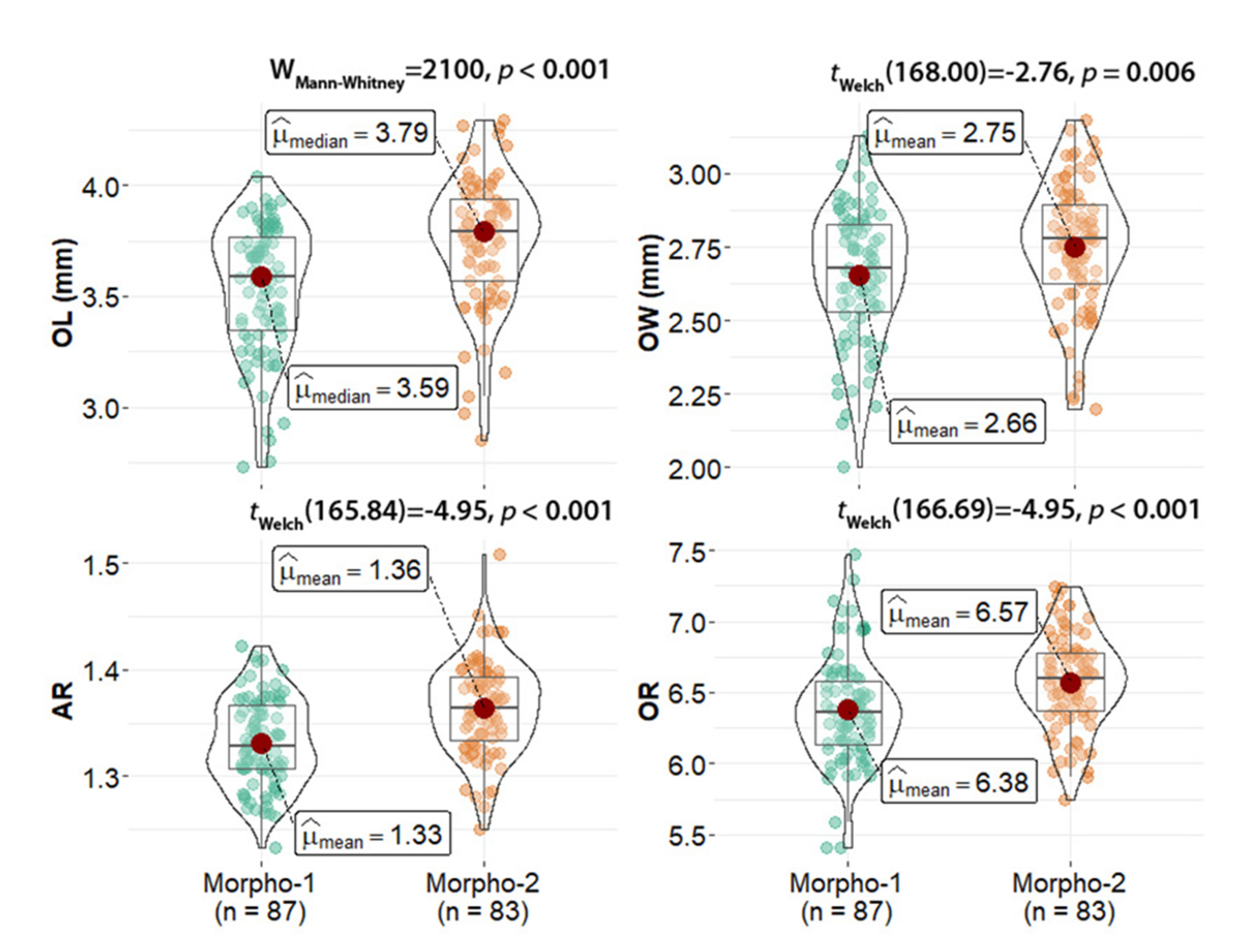
3.1. Taxonomic Classification and Otolith Shape
3.2. Oceanographic Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robinson, C.; Steinberg, D.K.; Anderson, T.R.; Arístegui, J.; Carlson, C.A.; Frost, J.R.; Ghiglione, J.F.; Hernández-León, S.; Jackson, G.A.; Koppelmann, R.; et al. Mesopelagic Zone Ecology and Biogeochemistry—A Synthesis. Deep Sea Res. Part II Top Stud. Oceanogr. 2010, 57, 1504–1518. [Google Scholar] [CrossRef]
- Gjøsaeter, J.; Kawaguchi, K. A Review of the World Resources of Mesopelagic Fish. FAO Fish. Tech. Pap. 1980, 193, 123–134. [Google Scholar] [CrossRef]
- Irigoien, X.; Klevjer, T.A.; Røstad, A.; Martinez, U.; Boyra, G.; Acuña, J.L.; Bode, A.; Echevarria, F.; Gonzalez-Gordillo, J.I.; Hernández-León, S.; et al. Large Mesopelagic Fishes Biomass and Trophic Efficiency in the Open Ocean. Nat. Commun. 2014, 5, 3271. [Google Scholar] [CrossRef] [PubMed]
- Lam, V.; Pauly, D. Mapping the Global Biomass of Mesopelagic Fishes. Sea Around Us Proj. Newsl. 2005, 30, 4. [Google Scholar]
- Proud, R.; Cox, M.J.; Brierley, A.S. Biogeography of the Global Ocean’s Mesopelagic Zone Correspondence. Curr. Biol. 2017, 27, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Proud, R.; Olav Handegard, N.; Kloser, R.J.; Cox, M.J.; Brierley, A.S. From Siphonophores to Deep Scattering Layers: Uncertainty Ranges for the Estimation of Global Mesopelagic Fish Biomass. ICES J. Mar. Sci. 2019, 76, 718–733. [Google Scholar] [CrossRef]
- Drazen, J.C.; Sutton, T.T. Dining in the Deep: The Feeding Ecology of Deep-Sea Fishes. Ann. Rev. Mar. Sci. 2017, 9, 337–366. [Google Scholar] [CrossRef]
- Olivar, M.P.; Hulley, P.A.; Castellón, A.; Emelianov, M.; López, C.; Tuset, V.M.; Contreras, T.; Molí, B. Mesopelagic Fishes across the Tropical and Equatorial Atlantic: Biogeographical and Vertical Patterns. Prog. Oceanogr. 2017, 151, 116–137. [Google Scholar] [CrossRef]
- Salvanes, A.G.V.; Kristoffersen, J.B. Mesopelagic Fishes; Academic Press: Cambridge, MA, USA, 2001. [Google Scholar] [CrossRef]
- Eschmeyer, W.N.; Fricke, R.; van der, L.R. Catalog of Fishes, Electronic Version (3 January 2017); California Academy of Sciences: San Francisco, CA, USA, 2018. [Google Scholar]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016. [Google Scholar]
- Sutton, T.T.; Wiebe, P.H.; Madin, L.; Bucklin, A. Diversity and Community Structure of Pelagic Fishes to 5000 m Depth in the Sargasso Sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 2220–2233. [Google Scholar] [CrossRef]
- Brierley, A.S. Diel Vertical Migration. Curr. Biol. 2014, 24, R1074–R1076. [Google Scholar] [CrossRef]
- Bernal, A.; Olivar, M.P.; Maynou, F.; de Puelles, M.L.F. Diet and Feeding Strategies of Mesopelagic Fishes in the Western Mediterranean. Prog. Oceanogr. 2015, 135, 1–17. [Google Scholar] [CrossRef]
- García-Seoane, E.; Wienerroither, R.; Mork, K.A.; Underwood, M.J.; Melle, W. Biogeographical Patterns of Meso- and Bathypelagic Fish along a Northeastern Atlantic Transect. ICES J. Mar. Sci. 2021, 78, 1444–1457. [Google Scholar] [CrossRef]
- Sutton, T.T. Vertical Ecology of the Pelagic Ocean: Classical Patterns and New Perspectives. J. Fish. Biol 2013, 83, 1508–1527. [Google Scholar] [CrossRef] [PubMed]
- Angel, M.V. Vertical Profiles of Pelagic Communities in the Vicinity of the Azores Front and Their Implications to Deep Ocean Ecology. Prog. Oceanogr. 1989, 22, 1–46. [Google Scholar] [CrossRef]
- Catul, V.; Gauns, M.; Karuppasamy, P.K. A Review on Mesopelagic Fishes Belonging to Family Myctophidae. Rev. Fish. Biol. Fish. 2011, 21, 339–354. [Google Scholar] [CrossRef]
- Longhurst, A.R.; Bedo, A.W.; Harrison, W.G.; Head, E.J.H.; Sameoto, D.D. Vertical Flux of Respiratory Carbon by Oceanic Diel Migrant Biota. Deep Sea Res. Part A Oceanogr. Res. Pap. 1990, 37, 685–694. [Google Scholar] [CrossRef]
- Arístegui, J.; Agustí, S.; Duarte, C.M. Respiration in the Dark Ocean. Geophys. Res. Lett. 2003, 30, 1041. [Google Scholar] [CrossRef]
- Hernández-León, S.; Olivar, M.P.; Fernández de Puelles, M.L.; Bode, A.; Castellón, A.; López-Pérez, C.; Tuset, V.M.; González-Gordillo, J.I. Zooplankton and Micronekton Active Flux Across the Tropical and Subtropical Atlantic Ocean. Front. Mar. Sci. 2019, 6, 535. [Google Scholar] [CrossRef]
- Saba, G.K.; Burd, A.B.; Dunne, J.P.; Hernández-león, S.; Martin, A.H.; Rose, K.A.; Salisbury, J.; Steinberg, D.K.; Trueman, C.N.; Wilson, R.W.; et al. Toward a Better Understanding of Fish-Based Contribution to Ocean Carbon Fl Ux. Limnol. Oceanogr. 2021, 66, 1639–1664. [Google Scholar] [CrossRef]
- Battaglia, P.; Malara, D.; Romeo, T.; Andaloro, F. Relationships between Otolith Size and Fish Size in Some Mesopelagic and Bathypelagic Species from the Mediterranean Sea (Strait of Messina, Italy). Sci. Mar. 2010, 74, 605–612. [Google Scholar] [CrossRef]
- Battaglia, P.; Malara, D.; Ammendolia, G.; Romeo, T. Relationships between Otolith Size and Fish Length in Some Mesopelagic Teleosts (Myctophidae, Paralepididae, Phosichthyidae and Stomiidae). J. Fish. Biol. 2015, 87, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Berdar, A.; Cavaliere, A.; Cavallaro, G.; Giuffrè, G.; Potoschi, A. Lo Studio Degli Organismi Spiaggiati Nello Stretto Di Messina Negli Ultimi Due Secoli. Nat. Sicil SIV 1983, 7, 3–17. [Google Scholar]
- Cavallaro, M.; Ammendolia, G.; Andaloro, F.; Battaglia, P. First Record of the Mesopelagic Fish Diaphus dumerilii (Bleeker, 1856) in the Mediterranean Sea. Mar. Biodivers. 2017, 47, 585–588. [Google Scholar] [CrossRef]
- Battaglia, P.; Sabatino, G.; Ibáñez, A.L.; Andaloro, F. Morphology and Ontogenetic Changes in Otoliths of the Mesopelagic Fishes Ceratoscopelus maderensis (Myctophidae), Vinciguerria attenuata and V. poweriae (Phosichthyidae) from the Strait of Messina (Mediterranean Sea). Acta Zool. 2017, 99, 126–142. [Google Scholar] [CrossRef]
- Carlo, B.S.; Costanzo, G.; Fresil, E.; Guglielmo, L.; Ianoral, A. Feeding Ecology and Stranding Mechanisms in Two Lanternfishes, Hygophum benoiti and Myctophum punctatum. Mar. Ecol Prog Ser. 1982, 9, 13–24. [Google Scholar] [CrossRef]
- Vittoria, M.M.; Sergio, S.; Andrea, T.; Fabrizio, V.; Marco, O.; Capua, D. The Strait of Messina Where Deep-Sea Fishes Land on the Beach. Deep Sea Life 2017, 9, 12–14. [Google Scholar]
- Battaglia, P.; Pedà, C.; Malara, D.; Milisenda, G.; MacKenzie, B.R.; Esposito, V.; Consoli, P.; Vicchio, T.M.; Stipa, M.G.; Pagano, L.; et al. Importance of the Lunar Cycle on Mesopelagic Foraging by Atlantic Bluefin Tuna in the Upwelling Area of the Strait of Messina (Central Mediterranean Sea). Animals 2022, 12, 2261. [Google Scholar] [CrossRef]
- Badcock, J. The Vertical Distribution of Mesopelagic Fishes Collected on the SOND Cruise. J. Mar. Biol. Assoc. 1970, 50, 1001–1044. [Google Scholar] [CrossRef]
- Foxton, P. SOND Cruise 1965 Biological Sampling Methods and Procedures. J. Mar. Biol. Assoc. 1969, 49, 603–620. [Google Scholar] [CrossRef]
- Bordes, F.; Wienerroither, R.; Uiblein, F.; Moreno, T.; Bordes, I.; Hernández-García, V.; Caballero, C. Catálogo de Especies Meso y Batipelágicas. Peces, Moluscos y Crustáceos. Colectadas Con Arrastre En Las Islas Canarias, Durante Las Campañas Realizadas a Bordo de B/E “La Bocaina”; Instituto Canarios de Ciencias Marinas (ICCM), Agencia Canaria de Investigación: Las Palmas de GC, Spain, 2009; p. 326. [Google Scholar]
- Wienerroither, R.; Uibleina, F.; Bordes, F.; Moreno, T. Composition, Distribution, and Diversity of Pelagic Fishes around the Canary Islands, Eastern Central Atlantic. Mar. Biol. Res. 2009, 5, 328–344. [Google Scholar] [CrossRef]
- Ariza, A.; Landeira, J.M.; Escánez, A.; Wienerroither, R.; Aguilar de Soto, N.; Røstad, A.; Kaartvedt, S.; Hernández-León, S. Vertical Distribution, Composition and Migratory Patterns of Acoustic Scattering Layers in the Canary Islands. J. Mar. Syst. 2016, 157, 82–91. [Google Scholar] [CrossRef]
- Foxton, P. The Vertical Distribution of Pelagic Decapods [Crustacea: Natantia] Collected on the Sond Cruise 1965 I. The Caridea. J. Mar. Biol. Assoc. 1970, 50, 939–960. [Google Scholar] [CrossRef]
- Foxton, P. The Vertical Distribution of Pelagic Decapods [Crustacea: Natantia] Collected on the Sond Cruise 1965 II. The Penaidea and General Discussion. J. Mar. Biol. Assoc. 1970, 50, 960–1000. [Google Scholar] [CrossRef]
- Clarke, M.R. Cephalopoda Collected on the SOND Cruise. J. Mar. Biol. Assoc. 1969, 49, 961–976. [Google Scholar] [CrossRef]
- Baker, A.D.C. The Vertical Distribution of Euphausiids Near Fuerteventura, Canary Islands (‘Discovery’ Sond Cruise, 1965). J. Mar. Biol. Assoc. 1970, 50, 301–342. [Google Scholar] [CrossRef]
- Sarmiento-Lezcano, A.N.; Triay-Portella, R.; Castro, J.J.; Rubio-Rodríguez, U.; Pajuelo, J.G. Age-Based Life-History Parameters of the Mesopelagic Fish Notoscopelus resplendens (Richardson, 1845) in the Central Eastern Atlantic. Fish. Res. 2018, 204, 412–423. [Google Scholar] [CrossRef]
- Sarmiento-Lezcano, A.; Triay-Portella, R.; Guerra-Marrero, A.; Jiménez-Alvarado, D.; Rubio-Rodríguez, U.; Núñez-González, R.; Bordes, F.; Castro, J.J. Contribution to the Reproductive Ecology of Notoscopelus resplendens (Richardson, in the Central—Eastern Atlantic. Sci. Rep. 2020, 10, 15821. [Google Scholar] [CrossRef]
- Guerra-Marrero, A.; Hernández-García, V.; Sarmiento-Lezcano, A.; Jiménez-Alvarado, D.; Pino, A.S.; Castro, J.J. Migratory Patterns, Vertical Distributions and Diets of Abralia veranyi and Abraliopsis morisii (Cephalopoda: Enoploteuthidae) in the Eastern North Atlantic. J. Molluscan Stud. 2019, 86, 27–43. [Google Scholar] [CrossRef]
- Bordes, F.; Uiblein, F.; Castillo, R.; Barrera-Luján, A.; Castro, J.J.; Coca, J.; Gómez, J.; Hansen, K.; Hernández-García, V.; Merrett, N.; et al. Epi- and Mesopelagic FIshes, Acoustic Data, and SST Images Collected off Lanzarote, Fuerteventura and Gran Canaria, Canary Islands, during Cruise ‘La Bocaina 04-97’. In Informes Técnicos del Instituto Canario de Ciencias Marinas; Agencia Canaria de Investigación: Las Palmas de GC, Spain, 1999; pp. 1–45. [Google Scholar]
- Hulley, P.A. Results of the Research Cruises of FRV” Walther Herwig” to South America. LVIII. Family Myctophidae (Osteichthyes, Myctophiformes). Arch. Für Fisch. 1981, 31, 1–300. [Google Scholar]
- Hulley, P.A. Myctophidae Paris. In Fishes of the North-Eastern Atlantic and the Mediterranean; Whitehead, P., Bauchot, M., Hureau, J., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, French, 1984; Volume 1, pp. 429–483. [Google Scholar]
- Hulley, P.A.; Paxton, J.R. Myctophidae. Bony Fishes, Part 1 (Elopiformes-Scorpaeniformes), The Living Marine Resources of the Eastern Central Atlantic 3; Carpenter, K., de Angelis, N., Eds.; FAO: Rome, Italy, 2016; Volume 3, pp. 1860–1928. [Google Scholar]
- Lombarte, A.; Chic, O.; Parisi-Baradad, V.; Olivella, R.; Piera, J.; Garca-Ladona, E. A Web-Based Environment from Shape Analysis of Fish Otoliths. The AFORO Database. Sci. Mar. 2006, 70, 147–152. [Google Scholar] [CrossRef]
- Parisi-Baradad, V.; Lombarte, A.; Garcia-Ladona, E.; Cabestany, J.; Piera, J.; Chic, O. Otolith Shape Contour Analysis Using Affine Transformation Invariant Wavelet Transforms and Curvature Scale Space Representation. Mar. Freshw. Res. 2005, 56, 795. [Google Scholar] [CrossRef]
- Parisi-Baradad, V.; Manjabacas, A.; Lombarte, A.; Olivella, R.; Chic, Ò.; Piera, J.; García-Ladona, E. Automated Taxon Identification of Teleost Fishes Using an Otolith Online Database—AFORO. Fish. Res. 2010, 105, 13–20. [Google Scholar] [CrossRef]
- Schwarzhans, W. A Comparative Morphological Study of the Recent Otoliths of the Genera Diaphus, Idiolychnus and Lobianchia (Myctophidae). Palaeo. Ichthyol. 2013, 13, 41–82. [Google Scholar]
- Lombarte, A.; Cruz, A. Otolith Size Trends in Marine Fish Communities from Different Depth Strata. J. Fish. Biol. 2007, 71, 53–76. [Google Scholar] [CrossRef]
- Sarmiento-Lezcano, A.N.; Couret, M.; Lombarte, A.; Olivar, M.P.; Landeira, J.M.; Hernández-León, S.; Tuset, V.M. Otolith Morphological Measures of Stranding Mesopelagic Fishes in the Canary Islands during June 2021. PANGAEA. 2022. Available online: https://doi.pangaea.de/10.1594/PANGAEA.951480 (accessed on 12 August 2022).
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation 2022. Available online: https://www.qgis.org/es/site/ (accessed on 20 July 2022).
- Battaglia, P.; Ammendolia, G.; Romeo, T.; Andaloro, F. Finding of Eretmophorus kleinenbergi (Moridae) in the Central Mediterranean Sea 93 Years after Its Previous Record, with a First Description of Sagittae and Fresh Coloration. Mar. Biodiv Rec. 2012, 5, 1–3. [Google Scholar] [CrossRef]
- Cavallaro, M.; Battaglia, P.; Laurà, R.; Guerrera, M.; Abbate, F.; Germanà, A. The Morphology Ofphotophores in the Garrick, Cyclothone braueri (Family: Gonostomatidae): An Ultrastructure Study. Acta Zool. 2015, 96, 296–300. [Google Scholar] [CrossRef]
- Cavallaro, M.; Mammola, C.L.; Verdiglione, R. Structural and Ultrastructural Comparison of Photophores of Two Species of Deep-Sea Fishes: Argyropelecus Hemigymnus and Maurolicus muelleri. J. Fish. Biol 2004, 64, 1552–1567. [Google Scholar] [CrossRef]
- Battaglia, P.; Ammendolia, G.; Cavallaro, M.; Consoli, P.; Esposito, V.; Malara, D.; Rao, I.; Romeo, T.; Andaloro, F. Influence of Lunar Phases, Winds and Seasonality on the Stranding of Mesopelagic Fish in the Strait of Messina (Central Mediterranean Sea). Mar. Ecol. 2017, 38, e12459. [Google Scholar] [CrossRef]
- Battaglia, P.; Canese, S.; Ammendolia, G.; Romeo, T.; Sandulli, R.; Tunesi, L.; Andaloro, F. New Records and Underwater Observation Ofthe Rare Fish Scorpaenodes arenai (Osteichthyes: Scorpaenidae) from the Central and Western Mediterranean. Sea. It J. Zool 2015, 82, 454–458. [Google Scholar] [CrossRef]
- Tuset, V.M.; Olivar, M.P.; Otero-Ferrer, J.L.; López-Pérez, C.; Hulley, P.A.; Lombarte, A. Morpho-Functional Diversity in Diaphus sp. (Pisces: Myctophidae) from the Central Atlantic Ocean: Ecological and Evolutionary Implications. Deep Sea Res. Part I Oceanogr. Res. Pap. 2018, 138, 46–59. [Google Scholar] [CrossRef]
- Brzobohaty, R.; Nolf, D. Diaphus Otoliths from the European Neogene (Myctophidae, Teleostei). Bull. L’institut R. Sci. Nat. Belg. Sci. Terre 2000, 70, 185–206. [Google Scholar]
- Winlker, N.S.; Paz-Goicoechea, M.; Lamb, R.W.; Pérez-Matus, A. Diet Reveals Linkesbetween Morphology Ad Foraging in a Cryptic Temperate Reeffish. Ecol. Evol. 2017, 7, 1124–1134. [Google Scholar]
- Webb, P.W. Body Form, Locomotion and Foraging in Aquatic Vertebrates. Am. Zool. 1984, 24, 107–120. [Google Scholar] [CrossRef]
- Czudaj, S.; Möllmann, C.; Fock, H.O. Length–Weight Relationships of 55 Mesopelagic Fishes from the Eastern Tropical. North Atlantic: Across- and Within-species Variation (Body Shape, Growth Stanza, Condition Factor). J. Fish. Biol. 2022, 101, 26–41. [Google Scholar] [CrossRef]
- Arístegui, J.; Sangrá, P.; Hernández-León, S.; Cantón, M.; Hernández-Guerra, A.; Kerling, J.L. Island-Induced Eddies in the Canary Islands. Deep Sea Res. Part I Oceanogr. Res. Pap. 1994, 41, 1509–1525. [Google Scholar] [CrossRef]
- Tejera, A.; García-Weil, L.; Heywood, K.; Cantón-Garbín, M. Observations of Oceanic Mesoscale Features and Variability in the Canary Islands Area from ERS-1 Altimeter Data, Satellite Infrared Imagery and Hydrographic Measurements. Int. J. Remote Sens. 2002, 23, 4897–4916. [Google Scholar] [CrossRef]
- Sangrà, P.; Pascual, A.; Rodríguez-Santana, Á.; Machín, F.; Mason, E.; McWilliams, J.C.; Pelegrí, J.L.; Dong, C.; Rubio, A.; Arístegui, J.; et al. The Canary Eddy Corridor: A Major Pathway for Long-Lived Eddies in the Subtropical North Atlantic. Deep Sea Res. Part I Oceanogr. Res. Pap. 2009, 56, 2100–2114. [Google Scholar] [CrossRef]
- Basterretxea, G.; Arístegui, J. Mesoscale Variability in Phytoplankton Biomass Distribution and Photosynthetic Parameters in the Canary-NW African Coastal Transition Zone. Mar. Ecol Prog Ser. 2000, 197, 27–40. [Google Scholar] [CrossRef]
- Arístegui, J.; Tett, P.; Hernández-Guerra, A.; Basterretxea, G.; Montero, M.F.; Wild, K.; Sangrá, P.; Hernández-León, S.; Cantón, M.; García-Braun, J.A. The Influence of Island-Generated Eddies on Chlorophyll Distribution: A Study of Mesoscale Variation around Gran Canaria. Deep Sea Res. Part I Oceanogr. Res. Pap. 1997, 44, 71–96. [Google Scholar] [CrossRef]
- van Camp, L.; Nykjaer, L.; Mittelstaedt, E.; Schlittenhardt, P. Upwelling and Boundary Circulation off Northwest Africa as Depicted by Infrared and Visible Satellite Observations. Prog. Oceanogr. 1991, 26, 357–402. [Google Scholar] [CrossRef]
- Hernández-Guerra, A.; Arıstegui, J.; Cantón, M.; Nykjaer, L. Phytoplankton Pigment Patterns in the Canary Islands Area as Determined Using Coastal Zone Colour Scanner Data. Int. J. Remote Sens. 1993, 14, 1431–1437. [Google Scholar] [CrossRef]
- Barton, E.D.; Arístegui, J.; Tett, P.; Cantón, M.; García-Braun, J.; Hernández-León, S.; Nykjaer, J.; Almeida, C.; Almunia, J.; Ballesteros, S.; et al. The Transition Zone of the Canary Current Upwelling Region. Prog. Oceanogr. 1998, 41, 455–504. [Google Scholar] [CrossRef]
- Rodrıguez, J.M.; Barton, E.D.; Eve, L.; Hernández-León, S. Mesozooplankton and Ichthyoplankton Distribution around Gran Canaria, an Oceanic Island in the NE Atlantic. Deep Sea Res. Part I Oceanogr. Res. Pap. 2001, 48, 2161–2183. [Google Scholar] [CrossRef]
- Landeira, J.M.; Lozano-Soldevilla, F.; Hernández-León, S.; Barton, E.D. Spatial Variability of Planktonic Invertebrate Larvae in the Canary Islands Area. J. Mar. Biol. Assoc. 2010, 90, 1217–1225. [Google Scholar] [CrossRef]
- Hernández-León, S. Accumulation of Mesozooplankton in a Wake Area as a Causative Mechanism of the “Island-Mass Effect”. Mar. Biol. 1991, 109, 141–147. [Google Scholar] [CrossRef]
- Hernández-León, S. Gradients of Mesozooplankton Biomass and ETS Activity in the Wind-Shear Area as Evidence of an Island Mass Effect in the Canary Island Waters. J. Plankton Res. 1988, 10, 1141–1154. [Google Scholar] [CrossRef]
- Moyano, M.; Rodríguez, J.M.; Benítez-Barrios, V.M.; Hernández-León, S. Larval Fish Distribution and Retention in the Canary Current System during the Weak Upwelling Season. Fish. Oceanogr. 2014, 23, 191–209. [Google Scholar] [CrossRef]
- Rodríguez, J.M.; Barton, E.D.; Hernández-León, S.; Arístegui, J. The Influence of Mesoscale Physical Processes on the Larval Fish Community in the Canaries CTZ, in Summer. Prog. Oceanogr. 2004, 62, 171–188. [Google Scholar] [CrossRef]
- Olivar, M.P.; Sabatés, A.; Pastor, M.V.; Pelegrí, J.L. Water Masses and Mesoscale Control on Latitudinal and Cross-Shelf Variations in Larval Fish Assemblages off NW Africa. Deep Sea Res. Part I Oceanogr. Res. Pap. 2016, 117, 120–137. [Google Scholar] [CrossRef]
- Landeira, J.M.; Brochier, T.; Mason, E.; Lozano-Soldevilla, F.; Hernández-León, S.; Barton, E.D. Transport Pathways of Decapod Larvae under Intense Mesoscale Activity in the Canary-African Coastal Transition Zone: Implications for Population Connectivity. Sci. Mar. 2017, 81, 299. [Google Scholar] [CrossRef]
- Mittelstaedt, E. The Ocean Boundary along the Northwest African Coast: Circulation and Oceanographic Properties at the Sea Surface. Prog. Oceanogr. 1991, 26, 307–355. [Google Scholar] [CrossRef]
- Arístegui, J.; Barton, E.D.; Álvarez-Salgado, X.A.; Santos, A.M.P.; Figueiras, F.G.; Kifani, S.; Hernández-León, S.; Mason, E.; Machú, E.; Demarcq, H. Sub-Regional Ecosystem Variability in the Canary Current Upwelling. Prog. Oceanogr. 2009, 83, 33–48. [Google Scholar] [CrossRef]
- Hernández-León, S.; Almeida, C.; Portillo-Hahnefeld, A.; Gómez, M.; Rodríguez, J.M.; Arístegui, J. Zooplankton Biomass and Indices of Feeding and Metabolism in Relation to a Filament off the Northwest African Upwelling Zone. J. Mar. Res. 2002, 60, 327–346. [Google Scholar] [CrossRef]
- Hernández-León, S.; Gómez, M.; Arístegui, J. Mesozooplankton in the Canary Current System: The Coastal–Ocean Transition Zone. Prog. Oceanogr. 2007, 74, 397–421. [Google Scholar] [CrossRef]
- Rodrıguez, J.M.; Hernández-León, S.; Barton, E.D. Mesoscale Distribution of Fish Larvae in Relation to an Upwelling Filament off Northwest Africa. Deep Sea Res. Part I Oceanogr. Res. Pap. 1999, 46, 1969–1984. [Google Scholar] [CrossRef]
- Barton, E.D.; Arístegui, J.; Tett, P.; Navarro-Pérez, E. Variability in the Canary Islands Area of Filament-Eddy Exchanges. Prog. Oceanogr. 2004, 62, 71–94. [Google Scholar] [CrossRef]
- Rodríguez, J.M.; Moyano, M.; Hernández-León, S. The Ichthyoplankton Assemblage of the Canaries–African Coastal Transition Zone: A Review. Prog. Oceanogr. 2009, 83, 314–321. [Google Scholar] [CrossRef]
- Linkowski, T.B. Lunar Rhythms of Vertical Migrations Coded in Otolith Microstructure of North Atlantic Lanternfishes, Genus Hygophum (Myctophidae). Mar. Biol. 1996, 124, 495–508. [Google Scholar] [CrossRef]
- Tarling, G. The Effect of Lunar Eclipse on the Vertical Migration Behaviour of Meganyctiphanes norvegica (Crustacea: Euphausiacea) in the Ligurian Sea. J. Plankton Res. 1999, 21, 1475–1488. [Google Scholar] [CrossRef]
- Hernández-León, S.; Almeida, C.; Yebra, L.; Arístegui, J.; Fernández de Puelles, M.L.; García-Braun, J. Zooplankton Abundance in Subtropical Waters: Is There a Lunar Cycle? Sci. Mar. 2001, 65, 59–64. [Google Scholar] [CrossRef]
- Chiou, W.D.; Cheng, L.Z.; Chen, C.T. Effects of Lunar Phase and Habitat Depth on Vertical Migration patterns of the Sergestid Shrimp Acetes intermedius. Fish. Sci. 2003, 69, 277–287. [Google Scholar] [CrossRef]
- Benoit-Bird, K.J.; Au, W.W.L.; Wisdom, D.W. Nocturnal Light and Lunar Cycle Effects on Diel Migration of Micronekton. Limnol. Oceanogr. 2009, 54, 1789–1800. [Google Scholar] [CrossRef]
- Neumann, D. Tidal and Lunar Rhythms. Handbook of Behavioural Neurobiology; Aschoff, J., Ed.; Plenum: New York, NY, USA, 1981. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarmiento-Lezcano, A.N.; Couret, M.; Lombarte, A.; Olivar, M.P.; Landeira, J.M.; Hernández-León, S.; Tuset, V.M. Stranding of Mesopelagic Fishes in the Canary Islands. Animals 2022, 12, 3465. https://doi.org/10.3390/ani12243465
Sarmiento-Lezcano AN, Couret M, Lombarte A, Olivar MP, Landeira JM, Hernández-León S, Tuset VM. Stranding of Mesopelagic Fishes in the Canary Islands. Animals. 2022; 12(24):3465. https://doi.org/10.3390/ani12243465
Chicago/Turabian StyleSarmiento-Lezcano, Airam N., María Couret, Antoni Lombarte, María Pilar Olivar, José María Landeira, Santiago Hernández-León, and Víctor M. Tuset. 2022. "Stranding of Mesopelagic Fishes in the Canary Islands" Animals 12, no. 24: 3465. https://doi.org/10.3390/ani12243465
APA StyleSarmiento-Lezcano, A. N., Couret, M., Lombarte, A., Olivar, M. P., Landeira, J. M., Hernández-León, S., & Tuset, V. M. (2022). Stranding of Mesopelagic Fishes in the Canary Islands. Animals, 12(24), 3465. https://doi.org/10.3390/ani12243465