
Serological and Molecular Surveillance for SARS-CoV-2 Infection in Captive Tigers (Panthera tigris), Thailand
 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Study Population and Sample Collection
2.3. Virus and Cell Culture Work
2.4. Plaque Reduction Neutralization Test (PRNT) for SARS-CoV-2 Neutralizing Antibody Detection
2.5. RNA Isolation and SARS-CoV-2 Genome Detection
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 1 October 2022).
- Andersen, K.G.; Rambaut, A.; Lipkin, W.I.; Holmes, E.C.; Garry, R.F. The proximal origin of SARS-CoV-2. Nat. Med. 2020, 26, 450–452. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wu, Q.; Zhang, Z. Probable Pangolin Origin of SARS-CoV-2 Associated with the COVID-19 Outbreak. Curr. Biol. 2020, 30, 1346–1351. [Google Scholar] [CrossRef]
- SARS-CoV-2 in animals situation report. Available online: https://www.woah.org/app/uploads/2022/07/sars-cov-2-situation-report-14.pdf (accessed on 30 September 2022).
- Calvet, G.A.; Pereira, S.A.; Ogrzewalska, M.; Pauvolid-Correa, A.; Resende, P.C.; Tassinari, W.S.; Costa, A.P.; Keidel, L.O.; da Rocha, A.S.B.; da Silva, M.F.B.; et al. Investigation of SARS-CoV-2 infection in dogs and cats of humans diagnosed with COVID-19 in Rio de Janeiro, Brazil. PLoS ONE 2021, 16, e0250853. [Google Scholar] [CrossRef]
- Patterson, E.I.; Elia, G.; Grassi, A.; Giordano, A.; Desario, C.; Medardo, M.; Smith, S.L.; Anderson, E.R.; Prince, T.; Patterson, G.T. Evidence of exposure to SARS-CoV-2 in cats and dogs from households in Italy. Nat. Commun. 2020, 11, 6231. [Google Scholar] [CrossRef] [PubMed]
- Račnik, J.; Kočevar, A.; Slavec, B.; Korva, M.; Rus, K.R.; Zakotnik, S.; Zorec, T.M.; Poljak, M.; Matko, M.; Rojs, O.Z.; et al. Transmission of SARS-CoV-2 from Human to Domestic Ferret. Emerg. Infect. Dis. 2021, 27, 2450–2453. [Google Scholar] [CrossRef] [PubMed]
- Fenollar, F.; Mediannikov, O.; Maurin, M.; Devaux, C.; Colson, P.; Levasseur, A.; Fournier, P.E.; Raoult, D. Mink, SARS-CoV-2, and the Human-Animal Interface. Front. Microbiol. 2021, 12, 663815. [Google Scholar] [CrossRef]
- Hale, V.L.; Dennis, P.M.; McBride, D.S.; Nolting, J.M.; Madden, C.; Huey, D.; Ehrlich, M.; Grieser, J.; Winston, J.; Lombardi, D.; et al. SARS-CoV-2 infection in free-ranging white-tailed deer. Nature 2022, 602, 481–486. [Google Scholar] [CrossRef]
- Tan, C.C.S.; Lam, S.D.; Richard, D.; Owen, C.J.; Berchtold, D.; Orengo, C.; Nair, M.S.; Kuchipudi, S.V.; Kapur, V.; van Dorp, L.; et al. Transmission of SARS-CoV-2 from humans to animals and potential host adaptation. Nat. Commun. 2022, 13, 2988. [Google Scholar] [CrossRef]
- Farag, E.A.; Islam, M.M.; Enan, K.; El-Hussein, A.M.; Bansal, D.; Haroun, M. SARS-CoV-2 at the human-animal interface: A review. Heliyon 2021, 7, e08496. [Google Scholar] [CrossRef]
- Mathavarajah, S.; Dellaire, G. Lions, tigers and kittens too: ACE2 and susceptibility to COVID-19. Evol. Med. Public Health 2020, 2020, 109–113. [Google Scholar] [CrossRef]
- McAloose, D.; Laverack, M.; Wang, L.; Killian, M.L.; Caserta, L.C.; Yuan, F.; Mitchell, P.K.; Queen, K.; Mauldin, M.R.; Cronk, B.D. From people to Panthera: Natural SARS-CoV-2 infection in tigers and lions at the Bronx Zoo. mBio 2020, 11, e02220-20. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, S.L.; Diel, D.; Wang, L.; Zec, S.; Laverack, M.; Martins, M.; Caserta, L.C.; Killian, M.L.; Terio, K.; Olmstead, C.; et al. SARS-CoV-2 infection and longitudinal fecal screening in Malayan tigers (Panthera Tigris Jacksoni), Amur tigers (Panthera Tigris Altaica), and African lions (Panthera Leo Krugeri) at the Bronx Zoo, New York, USA. J. Zoo Wildl. Med. 2021, 51, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Karikalan, M.; Chander, V.; Mahajan, S.; Deol, P.; Agrawal, R.-K.; Nandi, S.; Rai, S.-K.; Mathur, A.; Pawde, A.; Singh, K.-P. Natural infection of Delta mutant of SARS-CoV-2 in Asiatic lions of India. Transbound. Emerg. Dis. 2021, 69, 3047–3055. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Kumar, N.; Bhatia, S.; Aasdev, A.; Kanniappan, S.; Sekhar, A.T.; Gopinadhan, A.; Silambarasan, R.; Sreekumar, C.; Dubey, C.K. SARS-CoV-2 Delta variant among Asiatic lions, India. Emerg. Infect. Dis. 2021, 27, 2723–2725. [Google Scholar] [CrossRef]
- Fernandez-Bellon, H.; Rodon, J.; Fernandez-Bastit, L.; Almagro, V.; Padilla-Sole, P.; Lorca-Oro, C.; Valle, R.; Roca, N.; Grazioli, S.; Trogu, T.; et al. Monitoring Natural SARS-CoV-2 Infection in Lions (Panthera leo) at the Barcelona Zoo: Viral Dynamics and Host Responses. Viruses 2021, 13, 1683. [Google Scholar] [CrossRef]
- Koeppel, K.N.; Mendes, A.; Strydom, A.; Rotherham, L.; Mulumba, M.; Venter, M. SARS-CoV-2 Reverse zoonoses to pumas and lions, South Africa. Viruses 2022, 14, 120. [Google Scholar] [CrossRef]
- WHO Thailand Weekly Situation Update No. 247. Available online: https://cdn.who.int/media/docs/default-source/searo/thailand/2022_09_14_tha-sitrep-247-covid-19.pdf?sfvrsn=d1b0bfca_1 (accessed on 1 October 2022).
- Jairak, W.; Charoenkul, K.; Chamsai, E.; Udom, K.; Chaiyawong, S.; Bunpapong, N.; Boonyapisitsopa, S.; Tantilertcharoen, R.; Techakriengkrai, N.; Surachetpong, S.; et al. First cases of SARS-CoV-2 infection in dogs and cats in Thailand. Transbound. Emerg. Dis. 2021, 69, e979–e991. [Google Scholar] [CrossRef]
- Udom, K.; Jairak, W.; Chamsai, E.; Charoenkul, K.; Boonyapisitsopa, S.; Bunpapong, N.; Techakriengkrai, N.; Amonsin, A. Serological survey of antibodies against SARS-CoV-2 in dogs and cats, Thailand. Transbound. Emerg. Dis. 2021, 69, 2140–2147. [Google Scholar] [CrossRef]
- Jairak, W.; Charoenkul, K.; Chamsai, E.; Udom, K.; Chaiyawong, S.; Hangsawek, A.; Waenkaew, S.; Mungaomklang, A.; Tangwangvivat, R.; Amonsin, A. Survey of SARS-CoV-2 in dogs and cats in high-risk areas during the second wave of COVID-19 outbreak, Thailand. Zoonoses Public Health 2022, 69, 737–745. [Google Scholar] [CrossRef]
- Jairak, W.; Chamsai, E.; Udom, K.; Charoenkul, K.; Chaiyawong, S.; Techakriengkrai, N.; Tangwangvivat, R.; Suwannakarn, K.; Amonsin, A. SARS-CoV-2 delta variant infection in domestic dogs and cats, Thailand. Sci. Rep. 2022, 12, 8403. [Google Scholar] [CrossRef]
- Schulz, C.; Martina, B.; Mirolo, M.; Müller, E.; Klein, R.; Volk, H.; Egberink, H.; Gonzalez-Hernandez, M.; Kaiser, F.; von Köckritz-Blickwede, M. SARS-CoV-2-specific antibodies in domestic cats during first COVID-19 wave, Europe. Emerg. Infect. Dis. 2021, 27, 3115–3118. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.L.; Anderson, E.R.; Cansado-Utrilla, C.; Prince, T.; Farrell, S.; Brant, B.; Smyth, S.; Noble, P.M.; Pinchbeck, G.L.; Marshall, N. SARS-CoV-2 neutralising antibodies in dogs and cats in the United Kingdom. CRVIRO 2021, 2, 100011. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.; Yip, C.C.; To, K.K.; Tang, T.H.; Wong, S.C.; Leung, K.H.; Fung, A.Y.; Ng, A.C.; Zou, Z.; Tsoi, H.W.; et al. Improved molecular diagnosis of COVID-19 by the novel, highly sensitive and specific COVID-19-RdRp/Hel real-time reverse transcription-polymerase chain reaction assay validated in vitro and with clinical specimens. J. Clin. Microbiol. 2020, 58, e00310-20. [Google Scholar] [CrossRef]
- Zhai, S.L.; Li, C.L.; Sun, M.F.; Zhang, J.F.; Zheng, C.; Liao, M. Natural infections of SARS-CoV-2 increased in animals: How should humans interact with animals? J. Med. Virol. 2022, 94, 3503–3505. [Google Scholar] [CrossRef]
- Wibmer, C.K.; Ayres, F.; Hermanus, T.; Madzivhandila, M.; Kgagudi, P.; Oosthuysen, B.; Lambson, B.E.; de Oliveira, T.; Vermeulen, M.; van der Berg, K.; et al. SARS-CoV-2 501Y.V2 escapes neutralization by South African COVID-19 donor plasma. Nat. Med. 2021, 27, 622–625. [Google Scholar] [CrossRef] [PubMed]
- Tada, T.; Zhou, H.; Dcosta, B.M.; Samanovic, M.I.; Chivukula, V.; Herati, R.S.; Hubbard, S.R.; Mulligan, M.J.; Landau, N.R. Increased resistance of SARS-CoV-2 Omicron variant to neutralization by vaccine-elicited and therapeutic antibodies. EBioMedicine 2022, 78, 103944. [Google Scholar] [CrossRef]
- Hachmann, N.P.; Miller, J.; Collier, A.-R.Y.; Ventura, J.D.; Yu, J.; Rowe, M.; Bondzie, E.A.; Powers, O.; Surve, N.; Hall, K.; et al. Neutralization Escape by SARS-CoV-2 Omicron Subvariants BA.2.12.1, BA.4, and BA.5. N. Engl. J. Med. 2022, 387, 86–88. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.P.; Zeng, C.; Qu, P.; Faraone, J.; Zheng, Y.M.; Carlin, C.; Bednash, J.S.; Zhou, T.; Lozanski, G.; Mallampalli, R.; et al. Neutralization of SARS-CoV-2 Omicron sub-lineages BA.1, BA.1.1, and BA.2. Cell Host Microbe 2022, 30, 1093–1102.e3. [Google Scholar] [CrossRef]
- Yu, J.; Collier, A.Y.; Rowe, M.; Mardas, F.; Ventura, J.D.; Wan, H.; Miller, J.; Powers, O.; Chung, B.; Siamatu, M.; et al. Neutralization of the SARS-CoV-2 Omicron BA.1 and BA.2 Variants. N. Engl. J. Med. 2022, 386, 1579–1580. [Google Scholar] [CrossRef]
- Lu, L.; Mok, B.W.Y.; Chen, L.L.; Chan, J.M.C.; Tsang, O.T.Y.; Lam, B.H.S.; Chuang, V.W.M.; Chu, A.W.H.; Chan, W.M.; Ip, J.D.; et al. Neutralization of Severe Acute Respiratory Syndrome Coronavirus 2 Omicron Variant by Sera From BNT162b2 or CoronaVac Vaccine Recipients. Clin. Infect. Dis. 2022, 75, e822–e826. [Google Scholar] [CrossRef]
- Bashor, L.; Gagne, R.B.; Bosco-Lauth, A.M.; Bowen, R.A.; Stenglein, M.; VandeWoude, S. SARS-CoV-2 evolution in animals suggests mechanisms for rapid variant selection. Proc. Natl. Acad. Sci. USA 2021, 118, e2105253118. [Google Scholar] [CrossRef] [PubMed]
- Delta Variant Responsible for over 98 Percent of New Infections. Available online: https://www.nationthailand.com/in-focus/40008034 (accessed on 22 July 2022).
- How Dangerous Is the Delta Variant (B.1.617.2)? Available online: https://asm.org/Articles/2021/July/How-Dangerous-is-the-Delta-Variant-B-1-617-2 (accessed on 1 October 2022).
- WHO Thailand Weekly Situation Update No. 209. Available online: https://cdn.who.int/media/docs/default-source/searo/thailand/2021_11_11_eng-sitrep-209-covid-19.pdf?sfvrsn=92f99090_3 (accessed on 22 July 2022).
- Grome, H.N.; Meyer, B.; Read, E.; Buchanan, M.; Cushing, A.; Sawatzki, K.; Levinson, K.J.; Thomas, L.S.; Perry, Z.; Uehara, A.; et al. SARS-CoV-2 Outbreak among Malayan Tigers and Humans, Tennessee, USA, 2020. Emerg. Infect. Dis. 2022, 28, 833–836. [Google Scholar] [CrossRef] [PubMed]
- Meekins, D.A.; Gaudreault, N.N.; Richt, J.A. Natural and Experimental SARS-CoV-2 Infection in Domestic and Wild Animals. Viruses 2021, 13, 1993. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Sample Collection | Place | No. of Tigers | No. of Samples | |||
|---|---|---|---|---|---|---|
| Year | Date | Female | Male | Serum | Swab | |
| 2020 | 29 May | WBC No.1 | 25 | 10 | 35 | NA |
| 21–22 October | WBC No.2 | 8 | 1 | 9 | NA | |
| 2021 | 26 April | WBC No.2 | 8 | - | 8 | NA |
| 19 August | WBC No.1 | - | 1 | 1 | NA | |
| 9 November | Zoo | 4 | 5 | 9 | 9 N, 9 R | |
| Total | 45 | 17 | 62 | 18 | ||
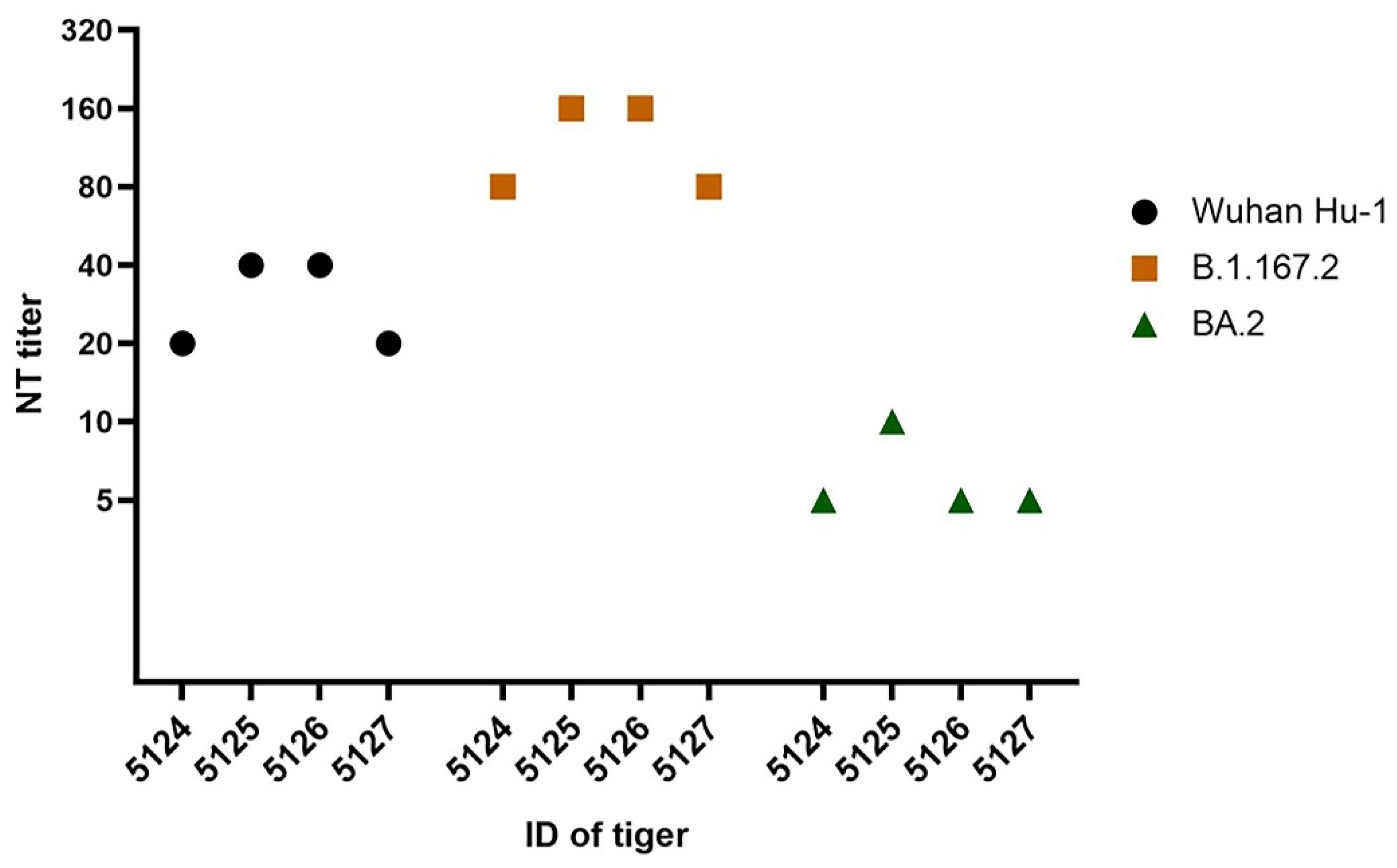
| Seropositive Tigers | Neutralizing Antibodies Titer a | |||
|---|---|---|---|---|
| ID | Sex | Wuhan Hu-1 | Delta (B.1.617.2) | Omicron (BA.2) |
| 5124 | female | 20 | 80 | <10 |
| 5125 | female | 40 | 160 | <10 |
| 5126 | female | 40 | 160 | 10 |
| 5127 | female | 20 | 80 | <10 |
| NC | female | <10 | <10 | <10 |
| PC | female | 160 | 20 | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sangkachai, N.; Chaiwattanarungruengpaisan, S.; Thongdee, M.; Suksai, P.; Tangsudjai, S.; Wongluechai, P.; Suwanpakdee, S.; Wiriyarat, W.; Buddhirongawatr, R.; Prasittichai, L.; et al. Serological and Molecular Surveillance for SARS-CoV-2 Infection in Captive Tigers (Panthera tigris), Thailand. Animals 2022, 12, 3350. https://doi.org/10.3390/ani12233350
Sangkachai N, Chaiwattanarungruengpaisan S, Thongdee M, Suksai P, Tangsudjai S, Wongluechai P, Suwanpakdee S, Wiriyarat W, Buddhirongawatr R, Prasittichai L, et al. Serological and Molecular Surveillance for SARS-CoV-2 Infection in Captive Tigers (Panthera tigris), Thailand. Animals. 2022; 12(23):3350. https://doi.org/10.3390/ani12233350
Chicago/Turabian StyleSangkachai, Nareerat, Somjit Chaiwattanarungruengpaisan, Metawee Thongdee, Parut Suksai, Siriporn Tangsudjai, Peerawat Wongluechai, Sarin Suwanpakdee, Witthawat Wiriyarat, Ruangrat Buddhirongawatr, Luxsana Prasittichai, and et al. 2022. "Serological and Molecular Surveillance for SARS-CoV-2 Infection in Captive Tigers (Panthera tigris), Thailand" Animals 12, no. 23: 3350. https://doi.org/10.3390/ani12233350
APA StyleSangkachai, N., Chaiwattanarungruengpaisan, S., Thongdee, M., Suksai, P., Tangsudjai, S., Wongluechai, P., Suwanpakdee, S., Wiriyarat, W., Buddhirongawatr, R., Prasittichai, L., Skulpong, A., Okada, P. A., Puthavathana, P., & Paungpin, W. (2022). Serological and Molecular Surveillance for SARS-CoV-2 Infection in Captive Tigers (Panthera tigris), Thailand. Animals, 12(23), 3350. https://doi.org/10.3390/ani12233350