
Combination of Cinnamaldehyde with Carvacrol or Thymol Improves the Mechanical Properties of Tibia in Post-Peak Laying Hens
 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Preparation of Essential Oils
2.3. Animal and Study Design
2.4. Sample and Data Collection
2.5. Mechanical Testing
2.6. Tibia Ash Concentration
2.7. Serum Biochemical Analysis
2.8. Intestine and Tibia Histomorphological Analysis
2.9. Cecal Microbiota Composition
2.10. Quantitation of mRNA Related Intestinal Barrier
2.11. Statistical Analysis
3. Results
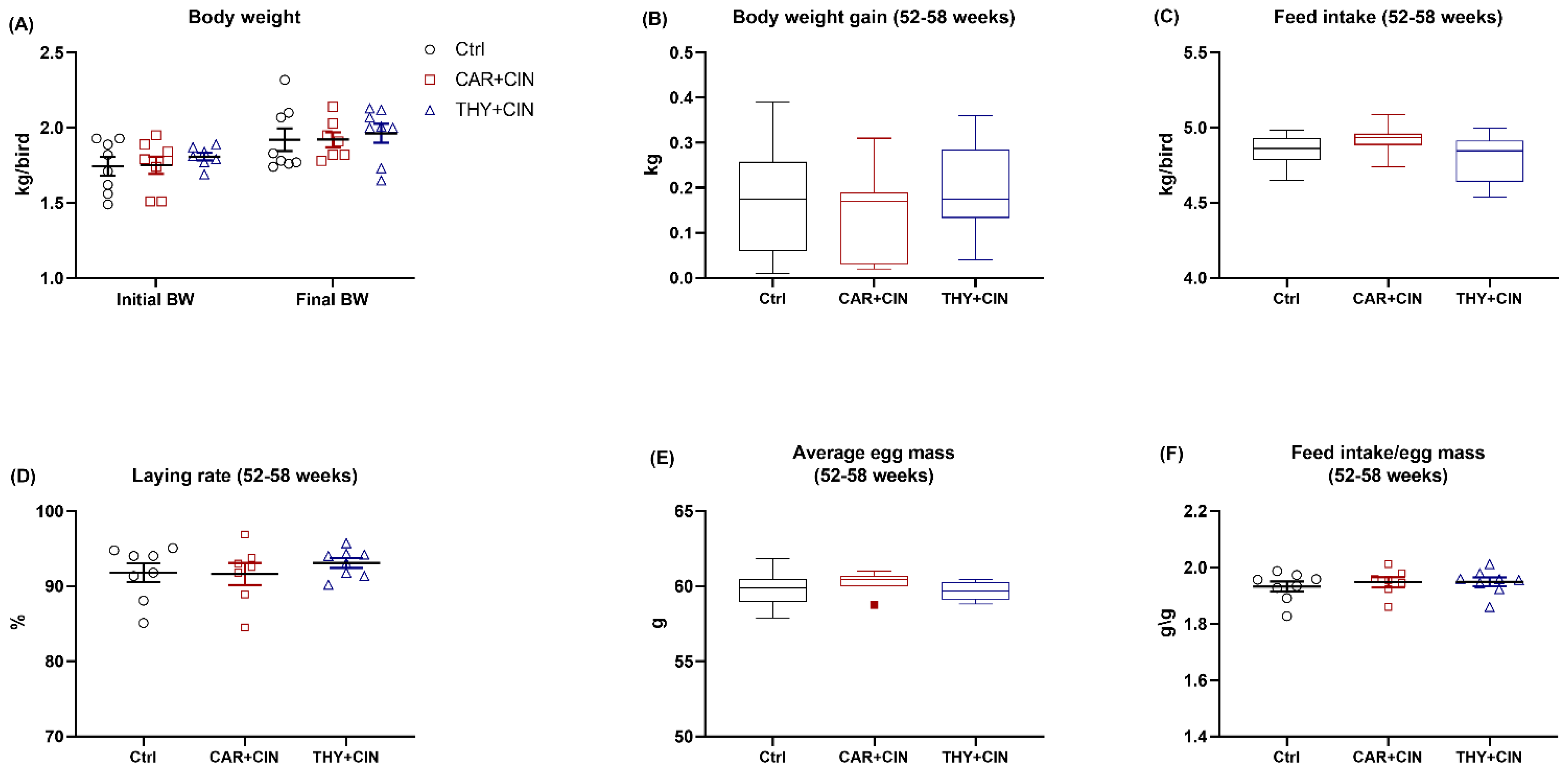
3.1. Dietary Essential Oils Did Not Affect Body Weight, Feed Intake, or Egg Production
3.2. Diet with Cinnamaldehyde and Carvacrol Decreased Tibial Width and Circumference
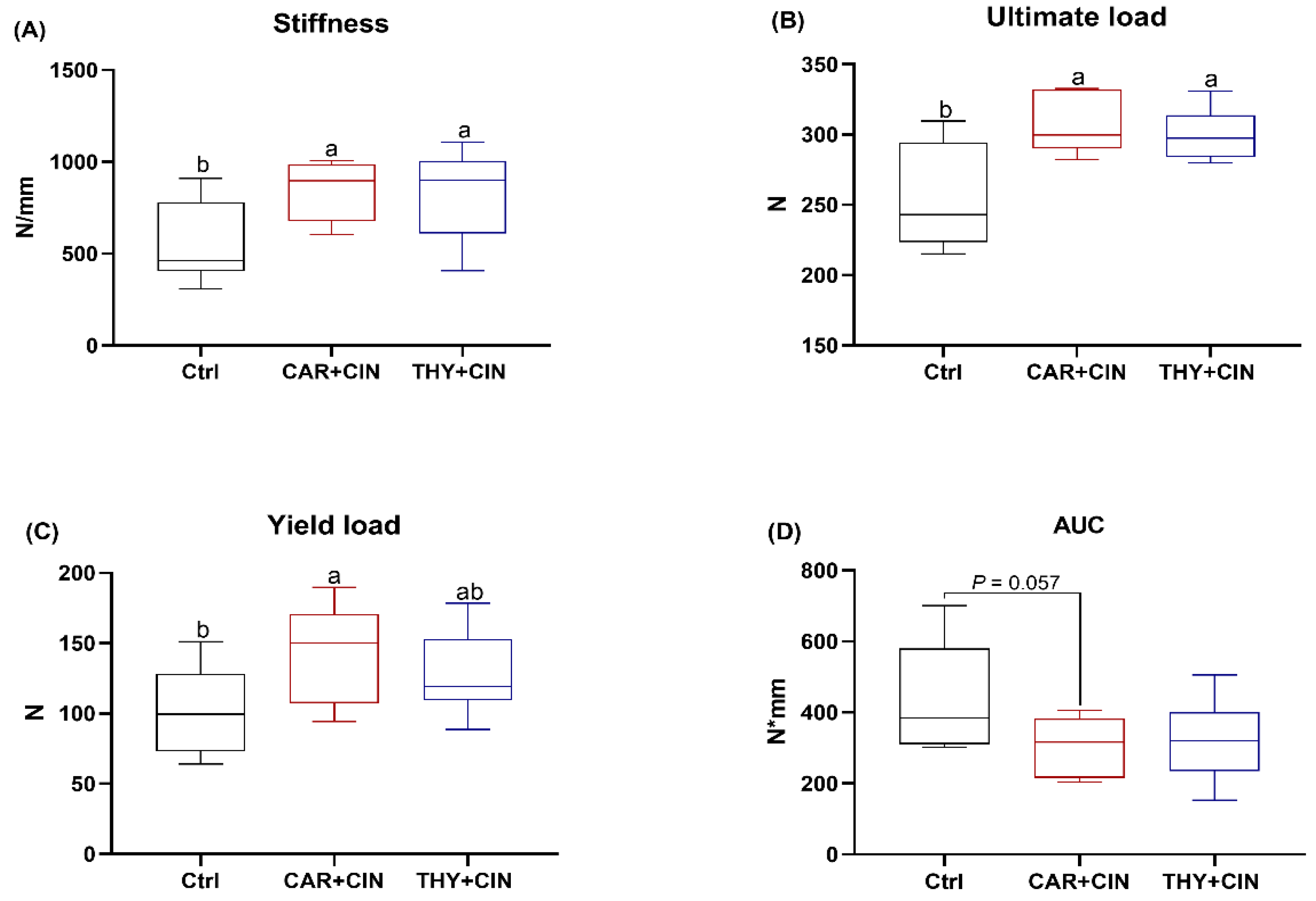
3.3. Mechanical Properties Are Improved by Essential Oils Supplementation
3.4. Histomorphology and Intestinal Barrier of Ileum Affected by Diet with Cinnamaldehyde and Carvacrol
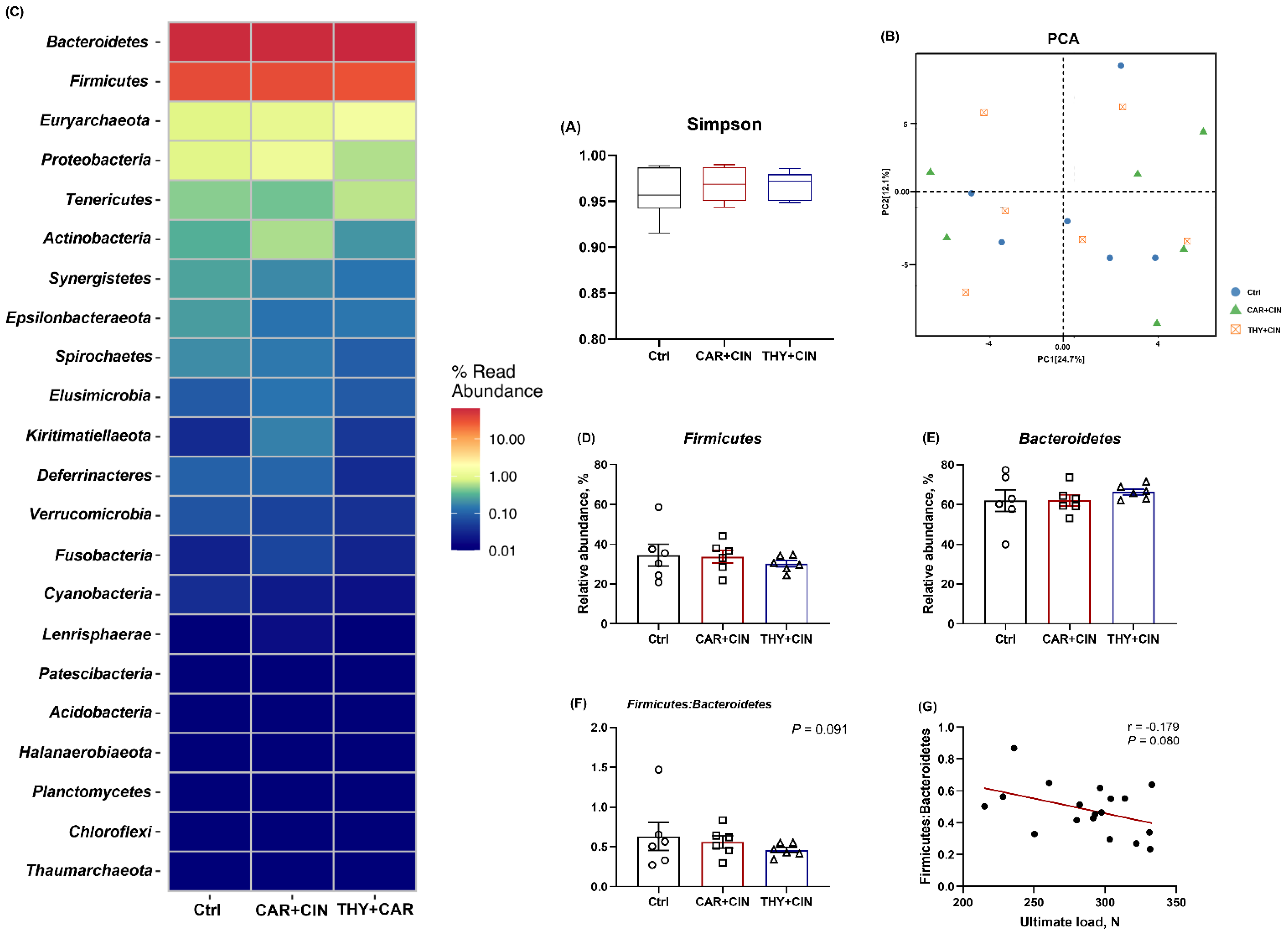
3.5. Cecal Microbiota Composition Was Not Affected by Diet
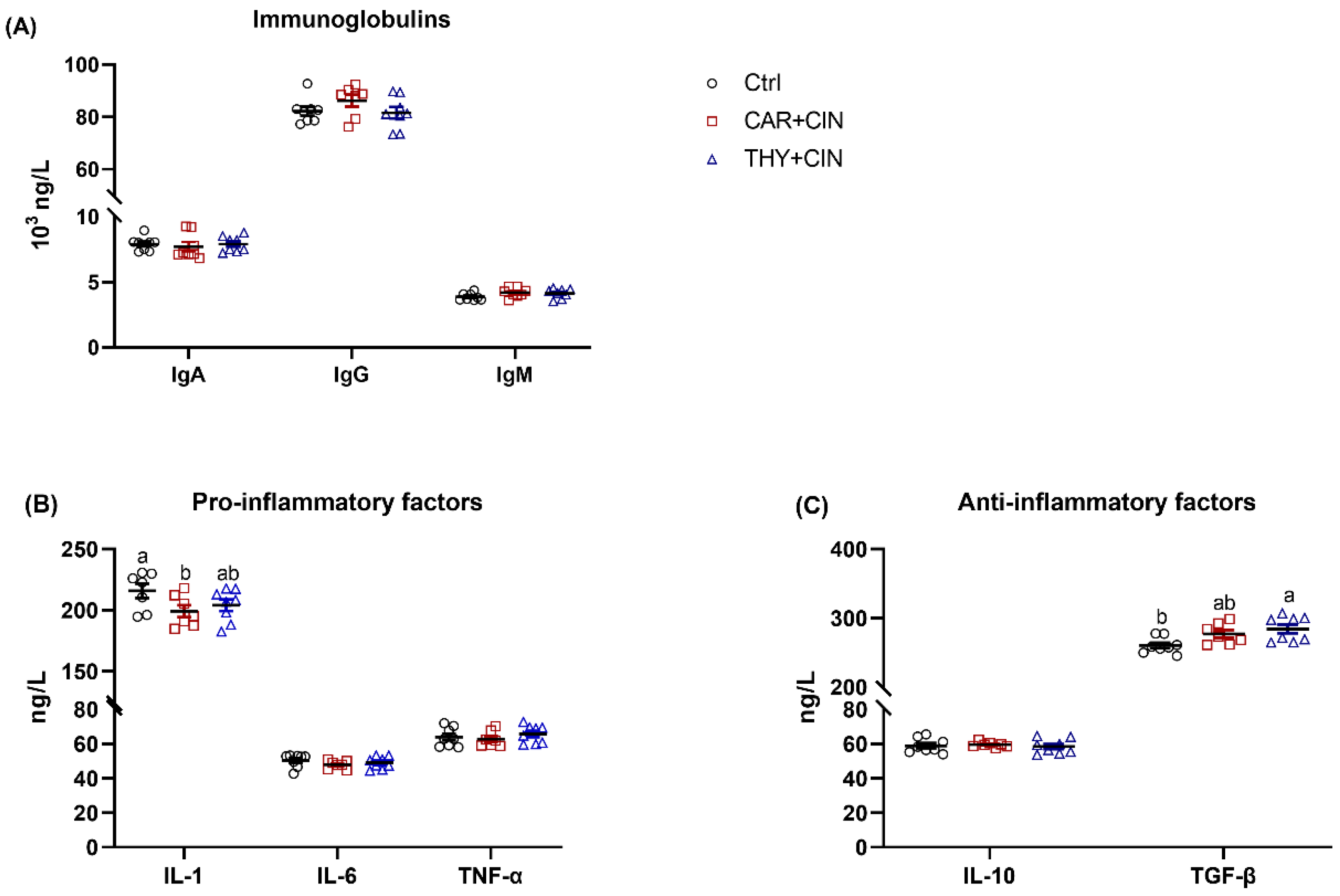
3.6. Dietary Essential Oils Supplementation Decreased Systemic Inflammatory Status
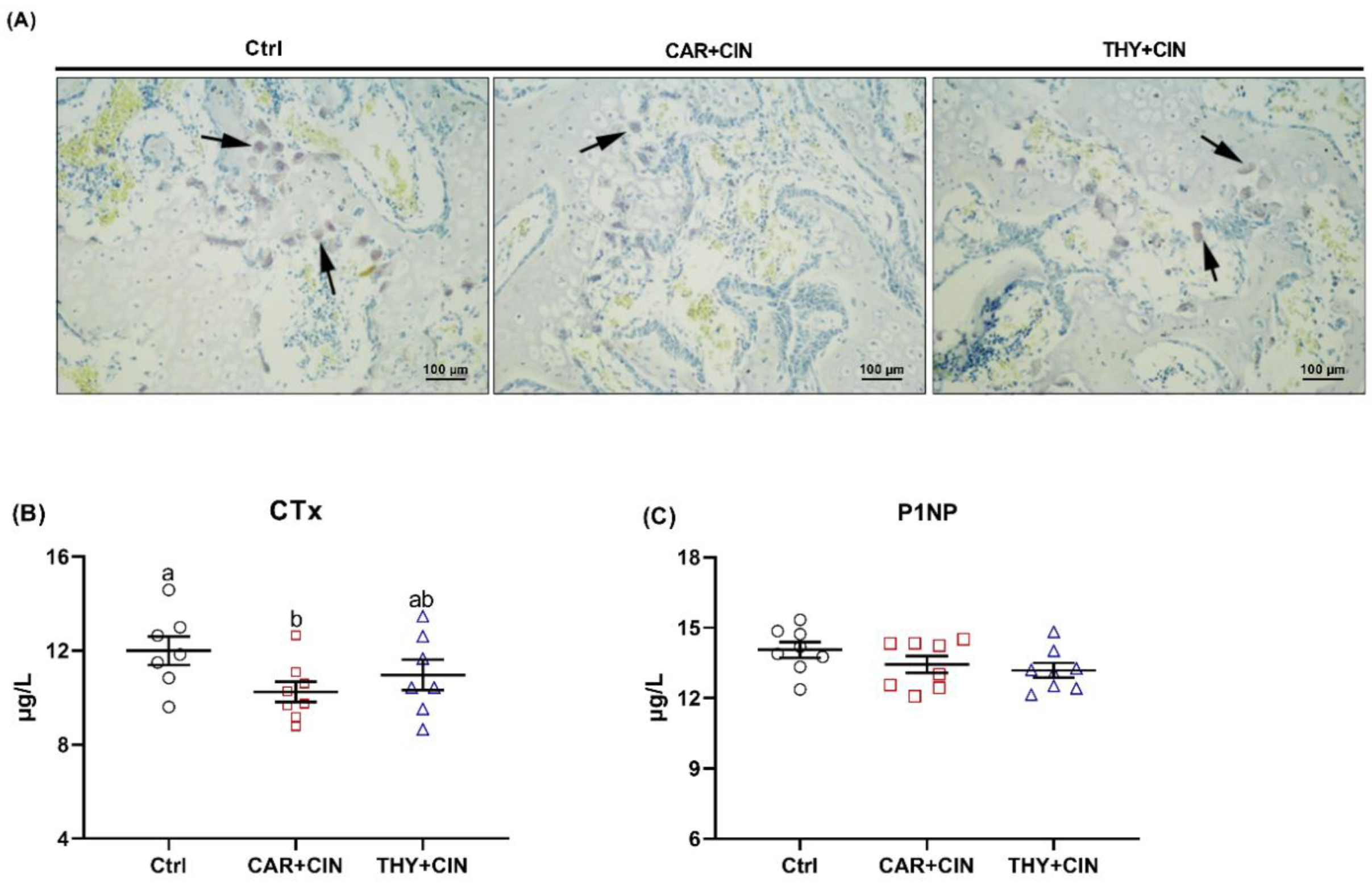
3.7. Diet with Essential Oils Inhibited Bone Resorption
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Jiang, J.; Xiao, S.; Xu, X.; Ma, H.; Feng, C.; Jia, X. Isomeric flavonoid aglycones derived from Epimedii Folium exerted different intensities in anti-osteoporosis through OPG/RANKL protein targets. Int. Immunopharmacol. 2018, 62, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Reis, J.H.; Gebert, R.R.; Barreta, M.; Baldissera, M.D.; Dos Santos, I.D.; Wagner, R.; Campigotto, G.; Jaguezeski, A.M.; Gris, A.; de Lima, J.L.F.; et al. Effects of phytogenic feed additive based on thymol, carvacrol and cinnamic aldehyde on body weight, blood parameters and environmental bacteria in broilers chickens. Microb. Pathog. 2018, 125, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Bi, Y.; Xin, H.; Pan, L.; Liu, R.; Li, X.; Li, J.; Zhang, R.; Bao, J. Keel fracture changed the behavior and reduced the welfare, production performance, and egg quality in laying hens housed individually in furnished cages. Poult. Sci. 2020, 99, 3334–3342. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Pan, L.; Li, C.; Zhao, P.; Li, J.; Zhang, R.; Bao, J. Dietary soybean oil supplementation affects keel bone characters and daily feed intake but not egg production and quality in laying hens housed in furnished cages. Front. Vet. Sci. 2021, 8, 657585. [Google Scholar] [CrossRef] [PubMed]
- Rodenburg, T.; Tuyttens, F.; De Reu, K.; Herman, L.; Zoons, J.; Sonck, B. Welfare assessment of laying hens in furnished cages and non-cage systems: An on-farm comparison. Anim. Welf. 2008, 17, 363–373. [Google Scholar]
- Xing, L.; Schwarz, E.M.; Boyce, B.F. Osteoclast precursors, RANKL/RANK, and immunology. Immunol. Rev. 2005, 208, 19–29. [Google Scholar] [CrossRef]
- Chatziravdeli, V.; Katsaras, G.N.; Lambrou, G.I. Gene expression in osteoblasts and osteoclasts under microgravity conditions: A systematic review. Curr. Genom. 2019, 20, 184–198. [Google Scholar] [CrossRef]
- Li, P.; Schwarz, E.M.; O’Keefe, R.J.; Ma, L.; Looney, R.J.; Ritchlin, C.T.; Boyce, B.F.; Xing, L. Systemic tumor necrosis factor alpha mediates an increase in peripheral CD11bhigh osteoclast precursors in tumor necrosis factor alpha-transgenic mice. Arthritis Rheum. 2004, 50, 265–276. [Google Scholar] [CrossRef]
- Poli, V.; Balena, R.; Fattori, E.; Markatos, A.; Yamamoto, M.; Tanaka, H.; Ciliberto, G.; Rodan, G.A.; Costantini, F. Interleukin-6 deficient mice are protected from bone loss caused by estrogen depletion. EMBO J. 1994, 13, 1189–1196. [Google Scholar] [CrossRef]
- D’Amelio, P.; Sassi, F. Gut Microbiota, immune system, and bone. Calcif. Tissue Int. 2018, 102, 415–425. [Google Scholar] [CrossRef]
- Irwin, R.; Raehtz, S.; Parameswaran, N.; McCabe, L.R. Intestinal inflammation without weight loss decreases bone density and growth. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R1149–R1157. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Z.; Luo, P.; Bai, S.; Chen, Y.; Chen, W. Dietary zinc glycine supplementation improves tibia quality of meat ducks by modulating the intestinal barrier and bone resorption. Biol. Trace Elem. Res. 2022, 1–16. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Rudyk, H.; Dobrowolski, P.; Donaldson, J.; Świetlicka, I.; Puzio, I.; Kamiński, D.; Wiącek, D.; Kushnir, V.; Brezvyn, O.; et al. Changes in the intestinal histomorphometry, the expression of intestinal tight junction proteins, and the bone structure and liver of pre-laying hens following oral administration of fumonisins for 21 days. Toxins 2021, 13, 375. [Google Scholar] [CrossRef]
- Chen, Y.C.; Greenbaum, J.; Shen, H.; Deng, H.W. Association between gut microbiota and bone health: Potential mechanisms and prospective. J. Clin. Endocrinol. Metab. 2017, 102, 3635–3646. [Google Scholar] [CrossRef]
- Schwarzer, M.; Makki, K.; Storelli, G.; Machuca-Gayet, I.; Srutkova, D.; Hermanova, P.; Martino, M.E.; Balmand, S.; Hudcovic, T.; Heddi, A.; et al. Lactobacillus plantarum strain maintains growth of infant mice during chronic undernutrition. Science 2016, 351, 854–857. [Google Scholar] [CrossRef]
- Ohlsson, C.; Nigro, G.; Boneca, I.G.; Backhed, F.; Sansonetti, P.; Sjogren, K. Regulation of bone mass by the gut microbiota is dependent on NOD1 and NOD2 signaling. Cell Immunol. 2017, 317, 55–58. [Google Scholar] [CrossRef]
- Sjogren, K.; Engdahl, C.; Henning, P.; Lerner, U.H.; Tremaroli, V.; Lagerquist, M.K.; Backhed, F.; Ohlsson, C. The gut microbiota regulates bone mass in mice. J. Bone Min. Res. 2012, 27, 1357–1367. [Google Scholar] [CrossRef]
- Zhang, H.; Majdeddin, M.; Gaublomme, D.; Taminiau, B.; Boone, M.; Elewaut, D.; Daube, G.; Josipovic, I.; Zhang, K.; Michiels, J. 25-hydroxycholecalciferol reverses heat induced alterations in bone quality in finisher broilers associated with effects on intestinal integrity and inflammation. J. Anim. Sci. Biotechnol. 2021, 12, 104. [Google Scholar] [CrossRef]
- Zhang, H.; Guo, Y.; Wang, Z.; Wang, Y.; Chen, B.; Du, P.; Zhang, X.; Huang, Y.; Li, P.; Michiels, J.; et al. Acidification of drinking water improvement tibia mass of broilers associated with the alterations in intestinal barrier and microbiota. Anim Biosci. 2022, 35, 902–915. [Google Scholar] [CrossRef]
- Palaska, I.; Papathanasiou, E.; Theoharides, T.C. Use of polyphenols in periodontal inflammation. Eur. J. Pharm. 2013, 720, 77–83. [Google Scholar] [CrossRef]
- Wei, H.K.; Xue, H.X.; Zhou, Z.X.; Peng, J. A carvacrol-thymol blend decreased intestinal oxidative stress and influenced selected microbes without changing the messenger RNA levels of tight junction proteins in jejunal mucosa of weaning piglets. Animal 2017, 11, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Michiels, J.; Missotten, J.; Van Hoorick, A.; Ovyn, A.; Fremaut, D.; De Smet, S.; Dierick, N. Effects of dose and formulation of carvacrol and thymol on bacteria and some functional traits of the gut in piglets after weaning. Arch. Anim. Nutr. 2010, 64, 136–154. [Google Scholar] [CrossRef] [PubMed]
- Deepak, V.; Kasonga, A.; Kruger, M.C.; Coetzee, M. Carvacrol inhibits osteoclastogenesis and negatively regulates the survival of mature osteoclasts. Biol. Pharm. Bull. 2016, 39, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.X.; Zhong, S.; Meng, X.B.; Zheng, N.Y.; Zhang, P.; Wang, Y.; Qin, L.; Wang, X.L. Cinnamaldehyde inhibits inflammation of human synoviocyte cells through regulation of Jak/Stat pathway and ameliorates collagen-induced arthritis in rats. J. Pharm. Exp. Ther. 2020, 373, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, M.; Li, L.; Kim, S.W.; Soh, Y. Thymol inhibits RANKL-induced osteoclastogenesis in RAW264.7 and BMM cells and LPS-induced bone loss in mice. Food Chem. Toxicol. 2018, 120, 418–429. [Google Scholar] [CrossRef] [PubMed]
- NY/T 33-2004; Nutrient Requirement of Meat-Type Chickens. China Agricultural Industry Standards: Beijing, China, 2004.
- Jepsen, K.J.; Silva, M.J.; Vashishth, D.; Guo, X.E.; van der Meulen, M.C. Establishing biomechanical mechanisms in mouse models: Practical guidelines for systematically evaluating phenotypic changes in the diaphyses of long bones. J. Bone Miner. Res. 2015, 30, 951–966. [Google Scholar] [CrossRef]
- Marume, U.; Mokagane, J.M.; Shole, C.O.; Hugo, A. Citrullus lanatus essential oils inclusion in diets elicit nutraceutical effects on egg production, egg quality, and physiological characteristics in layer hens. Poult. Sci. 2020, 99, 3038–3046. [Google Scholar] [CrossRef]
- Ruff, J.; Tellez, G., Jr.; Forga, A.J.; Senas-Cuesta, R.; Vuong, C.N.; Greene, E.S.; Hernandez-Velasco, X.; Uribe, A.J.; Martinez, B.C.; Angel-Isaza, J.A.; et al. Evaluation of three formulations of essential oils in broiler chickens under cyclic heat stress. Animals 2021, 11, 1084. [Google Scholar] [CrossRef]
- Currey, J.D. Bones: Structure and Mechanics; Princeton University Press: Princeton, NJ, USA, 2006. [Google Scholar]
- Zhang, H.; Zeng, Q.; Bai, S.; Wang, J.; Ding, X.; Xuan, Y.; Su, Z.; Zhang, K. Effect of graded calcium supplementation in low-nutrient density feed on tibia composition and bone turnover in meat ducks. Brit. J. Nutr. 2018, 120, 1217–1229. [Google Scholar] [CrossRef]
- Oftadeh, R.; Perez-Viloria, M.; Villa-Camacho, J.C.; Vaziri, A.; Nazarian, A. Biomechanics and mechanobiology of trabecular bone: A review. J. Biomech Eng.-T Asme 2015, 137, 0108021–01080215. [Google Scholar] [CrossRef]
- Tatara, M.; Majcher, P.; Krupski, W.; Studziński, T. Volumetric bone density, morphological and mechanical properties of femur and tibia in farm turkeys with leg deformities. B Vet. I Pulawy 2004, 48, 169–172. [Google Scholar]
- Wu, Z.; Weng, S.; Yan, D.; Xie, Z.; Zhou, Q.; Li, H.; Bai, B.; Boodhun, V.; Shen, Z.; Tang, J.; et al. Administration of cinnamaldehyde promotes osteogenesis in ovariectomized rats and differentiation of osteoblast in vitro. J. Pharm. Sci. 2018, 138, 63–70. [Google Scholar] [CrossRef]
- Cho, S.; Choi, Y.; Park, S.; Park, T. Carvacrol prevents diet-induced obesity by modulating gene expressions involved in adipogenesis and inflammation in mice fed with high-fat diet. J. Nutr. Biochem. 2012, 23, 192–201. [Google Scholar] [CrossRef]
- Hashemipour, H.; Khaksar, V.; Rubio, L.A.; Veldkamp, T.; van Krimpen, M.M. Effect of feed supplementation with a thymol plus carvacrol mixture, in combination or not with an NSP-degrading enzyme, on productive and physiological parameters of broilers fed on wheat-based diets. Anim. Feed Sci. Tech. 2016, 211, 117–131. [Google Scholar] [CrossRef]
- Pirgozliev, V.; Mansbridge, S.C.; Rose, S.P.; Lillehoj, H.S.; Bravo, D. Immune modulation, growth performance, and nutrient retention in broiler chickens fed a blend of phytogenic feed additives. Poult. Sci. 2019, 98, 3443–3449. [Google Scholar] [CrossRef]
- Schepper, J.D.; Collins, F.L.; Rios-Arce, N.D.; Raehtz, S.; Schaefer, L.; Gardinier, J.D.; Britton, R.A.; Parameswaran, N.; McCabe, L.R. Probiotic Lactobacillus reuteri prevents postantibiotic bone loss by reducing intestinal dysbiosis and preventing barrier disruption. J. Bone Miner. Res. 2019, 34, 681–698. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Proportion (%) | Calculated Analysis | Proportion (%) |
|---|---|---|---|
| Corn | 32.95 | AME, MJ/kg | 2620 |
| Wheat | 30.0 | CP | 16.30 |
| Soybean meal (46% CP) | 12.01 | Calcium | 3.60 |
| Wheat bran | 3.7 | Total phosphorus | 0.52 |
| Sprayed corn bran | 2.5 | Available phosphorus | 0.30 |
| Corn gluten | 1.5 | Lysine | 0.88 |
| DDGS a | 3.5 | Methionine + cystine | 0.67 |
| Sodium chloride | 0.24 | Threonine | 0.61 |
| Limestone | 8.6 | ||
| CaHPO4 | 0.43 | ||
| Chicken bone meal | 2.0 | ||
| Soybean oil | 0.57 | ||
| Premix b | 2.0 | ||
| Total | 100.0 |
| Gene ID | Gene | Primer Sequences (5′→3′) | Product Length, bp |
|---|---|---|---|
| XM_046925214.1 | ZO-1 | F: GAAGAGAGCACAGAACGCAG R: CACTTGTGGCAAGCTGAAGT | 123 |
| NM_001013611.2 | Claudin-1 | F: TCTGGTGTTAACGGGTGTGA R: GTCTTTGGTGGCGTGATCTT | 117 |
| NM_205128.1 | Occludin | F: CGTTCTTCACCCACTCCTCC R: CCAGAAGACGCGCAGTAAGA | 107 |
| NM_001039258.3 | CDH1 | F: AGCCAAGGGCCTGGATTATG R: GATAGGGGGCACGAAGACAG | 157 |
| NM_001318434.1 | MUC-2 | F: AGTGGCCATGGTTTCTTGTC R: TGCCAGCCTTTTTATGCTCT | 80 |
| NM_205518.1 | β-actin | F: GTCCACCGCAAATGCTTCTAA R: TGCGCATTTATGGGTTTTGTT | 78 |
| Item | Treatment | p-Value | ||
|---|---|---|---|---|
| Ctrl | CAR+CIN | THY+CIN | ||
| Fresh weight, g | 11.34 ± 0.51 | 10.35 ± 0.82 | 10.84 ± 0.83 | 0.053 |
| Relative weight, % body weight | 0.60 ± 0.06 | 0.54 ± 0.04 | 0.56 ± 0.06 | 0.171 |
| Length, mm | 119.14 ± 3.39 | 117.75 ± 3.68 | 117.82 ± 3.69 | 0.693 |
| Width, mm | 8.27 ± 0.27 a | 7.85 ± 0.34 b | 8.23 ± 0.24 ab | 0.020 |
| Circumference, cm | 2.61 ± 0.11 ab | 2.54 ± 0.05 b | 2.69 ± 0.06 a | 0.010 |
| Ash, % fat-free weight | 58.42 ± 2.36 | 58.40 ± 0.45 | 57.54 ± 2.06 | 0.778 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Wang, Y.; Wang, Y.; Wang, L.; Lv, X.; Cui, G.; Ji, L.; Huang, Y.; Michiels, J.; Chen, W. Combination of Cinnamaldehyde with Carvacrol or Thymol Improves the Mechanical Properties of Tibia in Post-Peak Laying Hens. Animals 2022, 12, 3108. https://doi.org/10.3390/ani12223108
Zhang H, Wang Y, Wang Y, Wang L, Lv X, Cui G, Ji L, Huang Y, Michiels J, Chen W. Combination of Cinnamaldehyde with Carvacrol or Thymol Improves the Mechanical Properties of Tibia in Post-Peak Laying Hens. Animals. 2022; 12(22):3108. https://doi.org/10.3390/ani12223108
Chicago/Turabian StyleZhang, Huaiyong, Yongshuai Wang, Yilu Wang, Leilei Wang, Xiangyun Lv, Guangya Cui, Longxiang Ji, Yanqun Huang, Joris Michiels, and Wen Chen. 2022. "Combination of Cinnamaldehyde with Carvacrol or Thymol Improves the Mechanical Properties of Tibia in Post-Peak Laying Hens" Animals 12, no. 22: 3108. https://doi.org/10.3390/ani12223108
APA StyleZhang, H., Wang, Y., Wang, Y., Wang, L., Lv, X., Cui, G., Ji, L., Huang, Y., Michiels, J., & Chen, W. (2022). Combination of Cinnamaldehyde with Carvacrol or Thymol Improves the Mechanical Properties of Tibia in Post-Peak Laying Hens. Animals, 12(22), 3108. https://doi.org/10.3390/ani12223108