
Molecular Characterization and Response of Prolyl Hydroxylase Domain (PHD) Genes to Hypoxia Stress in Hypophthalmichthys molitrix

 ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Fish
2.2. Exposure and Tissue Collection
2.3. Full-Length cDNA Cloning of PHD Genes
2.4. Sequence and Evolutionary Analysis
2.5. Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
2.6. Statistical Analysis
3. Results
3.1. Molecular Characterization of PHD cDNAs in H. molitrix
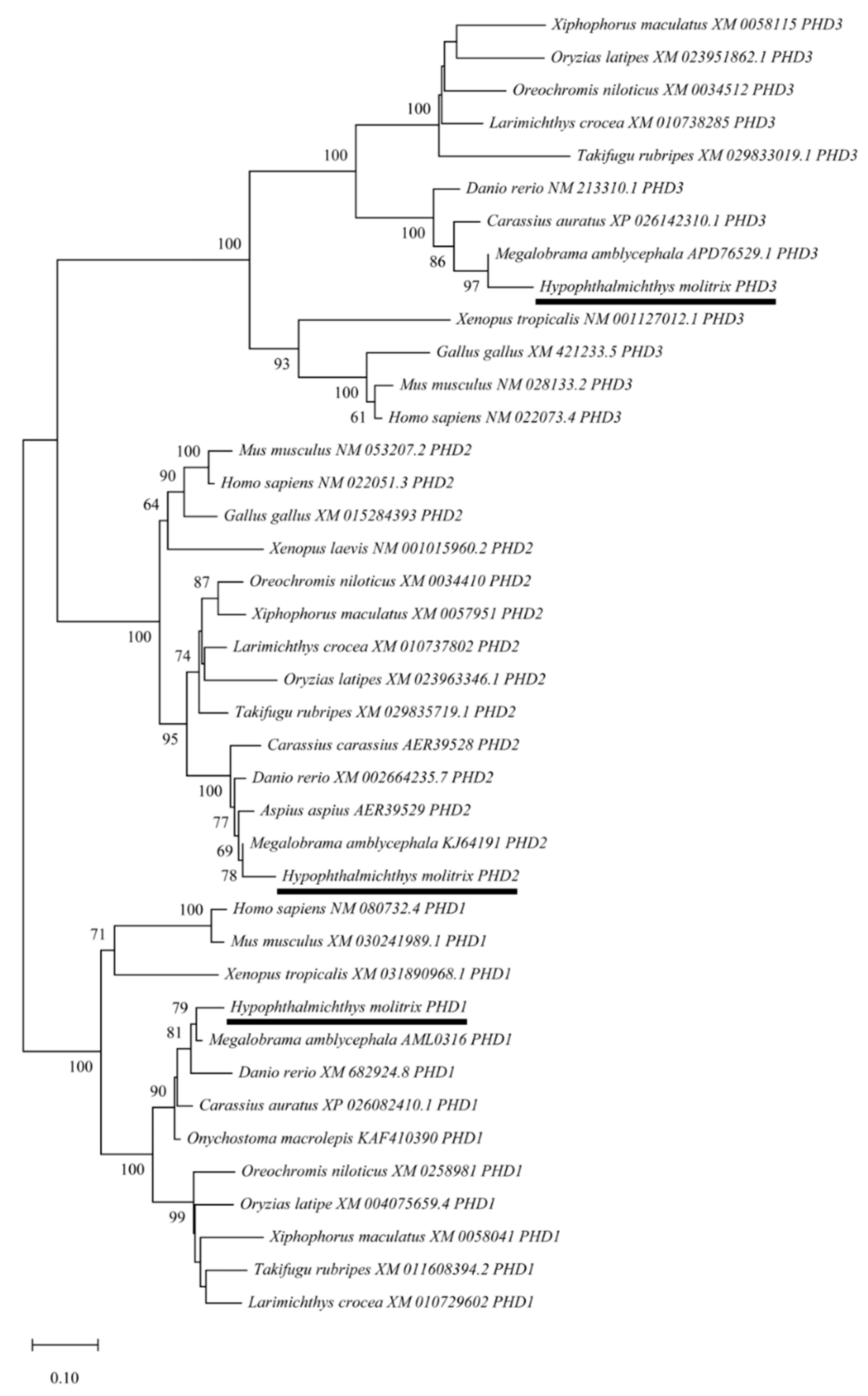
3.2. Evolutionary Relationships of PHD Genes
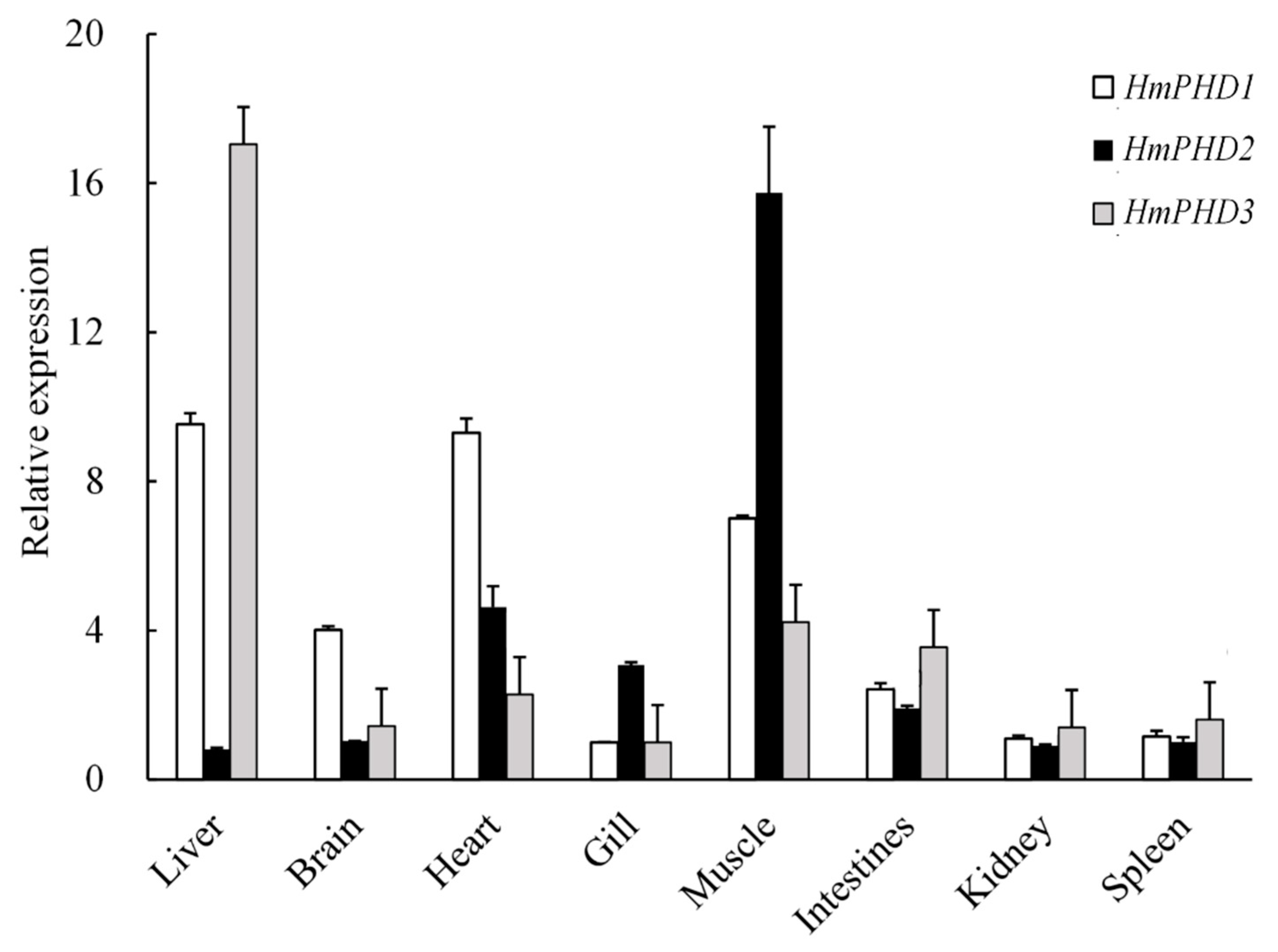
3.3. Transcriptional Expression Patterns of PHD Genes in H. molitrix
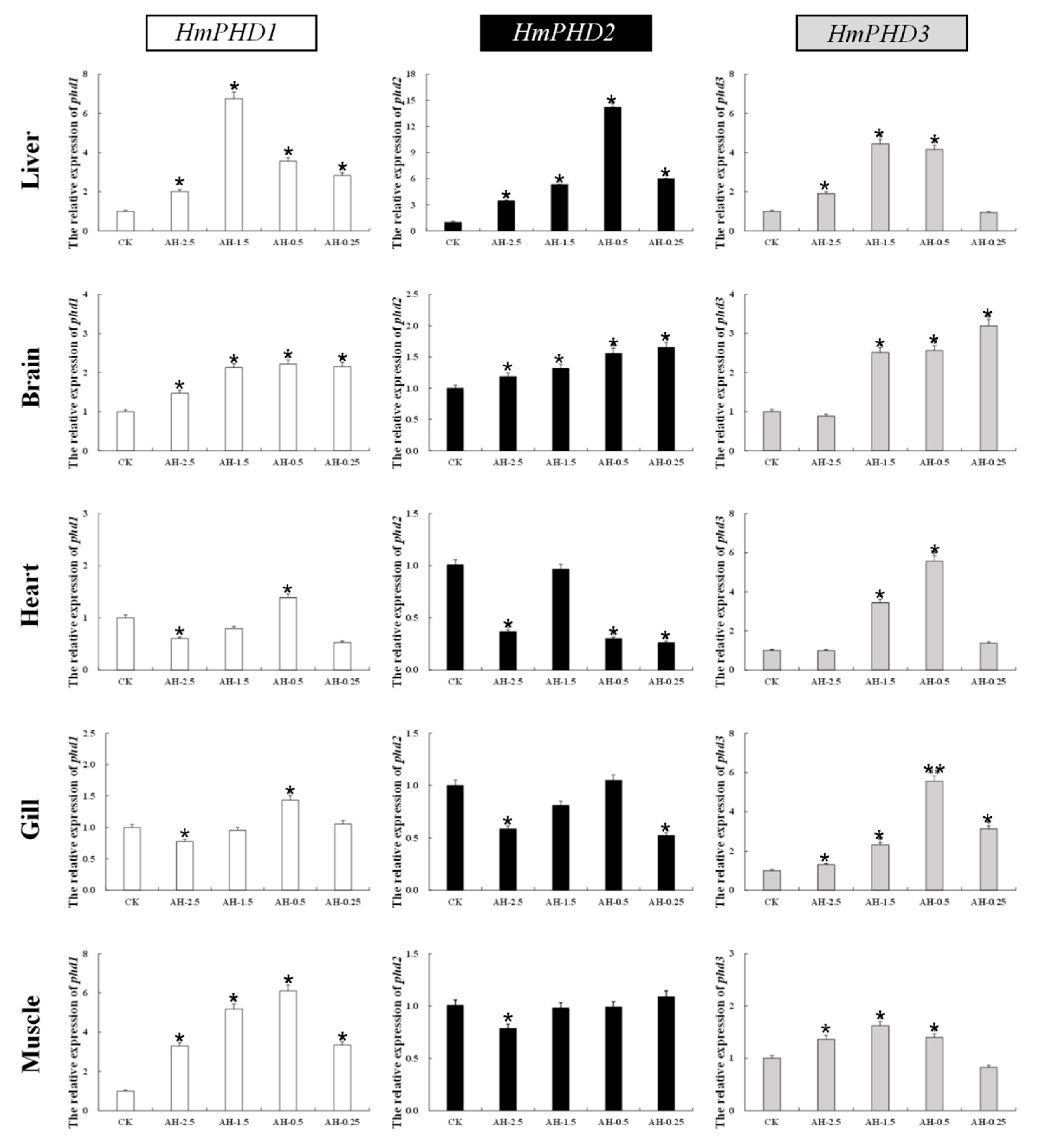
3.4. Transcriptional Response of PHD Genes in H. molitrix to Acute Hypoxia
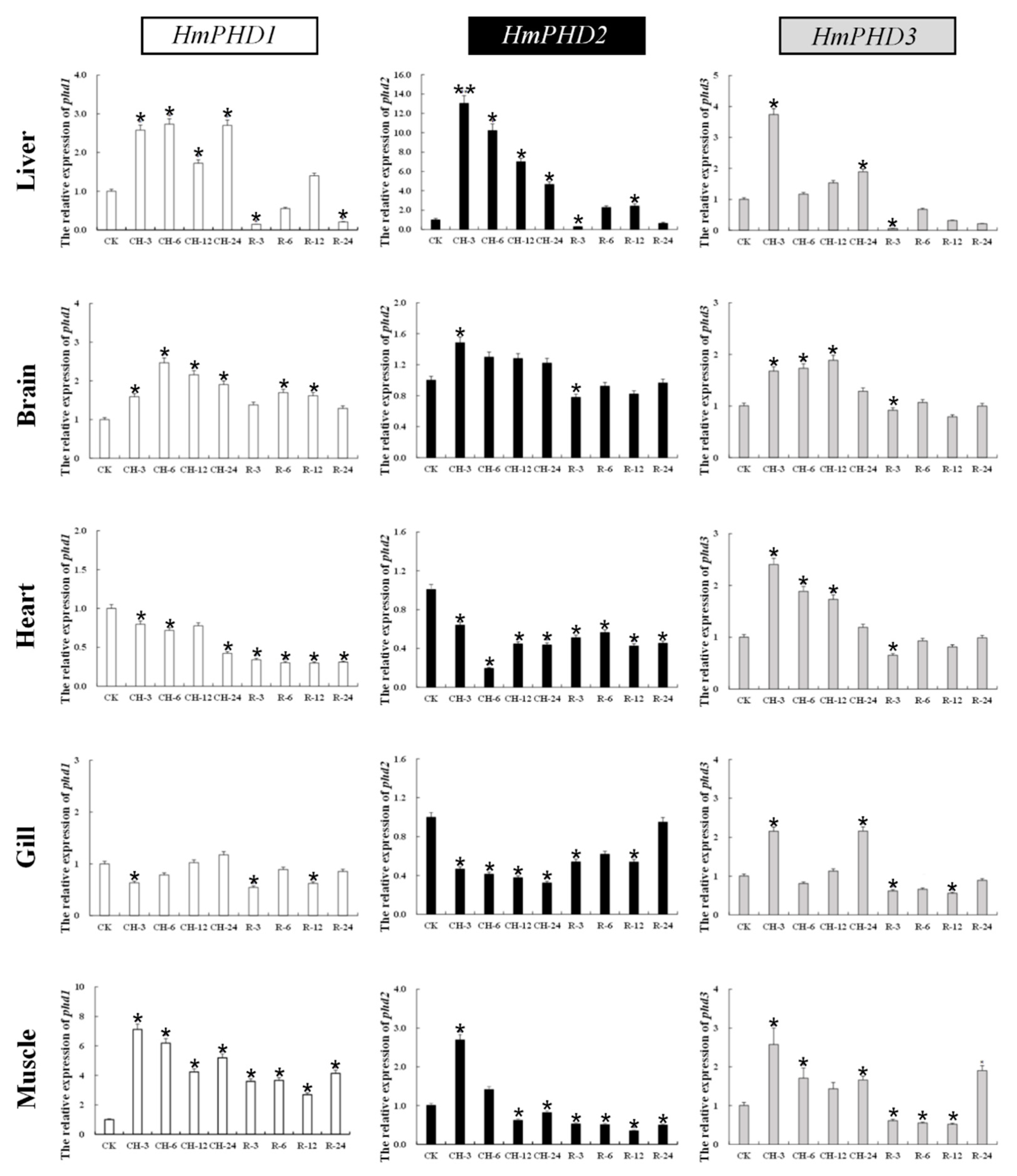
3.5. Transcriptional Response of PHD Genes to Continued Hypoxia and Reoxygenation in H. molitrix
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| bp | Base pair |
| PHD | Prolyl hydroxylase domain |
| HmPHD | PHD genes of H. molitrix |
| HmPHD | PHD proteins of H. molitrix |
| ORFs | Open reading frames |
| DO | Dissolved oxygen |
| HIFs | Hypoxia-inducible factors |
| ODDD | Oxygen dependent degradation domain |
| VHL | Von Hippel-Lindau tumor suppressor protein |
| HRE | Hypoxia response element |
| YFI | Yangtze River Fisheries Research Institute |
| AH | Acute hypoxia |
| CK | Control group |
| RT-PCR | Reverse transcription-polymerase chain reaction |
| RACE | Rapid amplification of cDNA ends |
| aa | amino acid |
References
- Giaccia, A.J.; Simon, M.C.; Johnson, R. The biology of hypoxia: The role of oxygen sensing in development, normal function, and disease. Genes Dev. 2004, 18, 2183–2194. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhao, Q.; Mooney, S.M.; Lee, F.S. Sequence determinants in hypoxia-inducible factor-1α for hydroxylation by the prolyl hydroxylases PHD1, PHD2, and PHD3. J. Biol. Chem. 2002, 277, 39792–39800. [Google Scholar] [CrossRef] [PubMed]
- Rytknen, K.T.; Williams, T.A.; Renshaw, G.M.; Primmer, C.R.; Nikinmaa, M. Molecular Evolution of the Metazoan PHD-HIF Oxygen-Sensing System. Mol. Biol. Evol. 2011, 28, 1913–1926. [Google Scholar] [CrossRef]
- Epstein, A.C.; Gleadle, J.; McNeill, L.; Hewitson, K.; O’rourke, J.; Mole, D.; Mukherji, M.; Metzen, E.; Wilson, M.; Dhanda, A.; et al. C. elegans EGL-9 and Mammalian Homologs Define a Family of Dioxygenases that Regulate HIF by Prolyl Hydroxylation. Cell 2001, 107, 43–54. [Google Scholar] [CrossRef]
- Bruick, R.K. A conserved family of prolyl-4-hydroxylases that modify HIF. Science 2001, 294, 1337–1340. [Google Scholar] [CrossRef]
- Taylor, C.T.; Pouyssegur, J. Oxygen, Hypoxia, and Stress. Ann. N. Y. Acad. Sci. 2007, 1113, 87–94. [Google Scholar] [CrossRef]
- Hochachka, P.W.; Lutz, P.L. Mechanism, origin, and evolution of anoxia tolerance in animals. Comp. Biochem. Physiol. Part B 2001, 130, 435–459. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Monier, M.N.; Hoseinifar, S.H.; Faggio, C. Fish response to hypoxia stress: Growth, physiological, and immunological biomarkers. Fish Physiol. Biochem. 2019, 45, 997–1013. [Google Scholar] [CrossRef] [PubMed]
- Kaelin, W.G., Jr.; Ratcliffe, P.J. Oxygen sensing by metazoans: The central role of the HIF hydroxylase pathway. Mol. Cell 2008, 30, 393–402. [Google Scholar] [CrossRef]
- Piontkivska, H.; Chung, J.S.; Ivanina, A.; Sokolov, E.; Techa, S.; Sokolova, I. Molecular characterization and mRNA expression of two key enzymes of hypoxia-sensing pathways in eastern oysters Crassostrea virginica (Gmelin): Hypoxia-inducible factor α (HIF-α) and HIF-prolyl hydroxylase (PHD). Comp. Biochem. Physiol. Part D Genom. Proteom. 2011, 6, 103–114. [Google Scholar] [CrossRef]
- Semenza, G.L. HIF-1, O2, and the 3 PHDs: How animal cells signal hypoxia to the nucleus. Cell 2001, 107, 1–20. [Google Scholar] [CrossRef]
- He, M.; Zhou, C.; Lu, Y.; Mao, L.; Xi, Y.; Mei, X.; Wang, X.; Zhang, L.; Yu, Z.; Zhou, Z. Melatonin antagonizes nickel-induced aerobic glycolysis by blocking ROS-mediated HIF-1α/miR210/ISCU axis activation. Oxidative Med. Cell. Longev. 2020, 2020, 5406284. [Google Scholar] [CrossRef]
- Maxwell, P.H.; Wiesener, M.S.; Chang, G.W.; Clifford, S.C.; Vaux, E.C.; Cockman, M.E.; Wykoff, C.C.; Pugh, C.W.; Maher, E.R.; Ratcliffe, P.J. The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature 1999, 399, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Majmundar, A.J.; Wong, W.J.; Simon, M.C. Hypoxia-inducible factors and the response to hypoxic stress. Mol. Cell 2010, 40, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Kong, P.; Yu, K.N.; Yang, M.; Almahi, W.A.; Nie, L.; Chen, G.; Han, W. Micheliolide Enhances Radiosensitivities of p53-Deficient Non-Small-Cell Lung Cancer via Promoting HIF-1α Degradation. Int. J. Mol. Sci. 2020, 21, 3392. [Google Scholar] [CrossRef] [PubMed]
- Freedman, S.J.; Sun, Z.; Poy, F.; Kung, A.L.; Eck, M.J. Structural basis for recruitment of CBP/p300 by hypoxia-inducible factor-1α. Proc. Natl. Acad. Sci. USA 2002, 99, 5367–5372. [Google Scholar] [CrossRef] [PubMed]
- Wenger, R.; Stiehl, D.; Camenisch, G. Integration of Oxygen Signaling at the Consensus HRE. Sci. STKE 2005, 2005, re12. [Google Scholar] [CrossRef]
- He, Y.; Munday, J.S.; Perrott, M.; Wang, G.; Liu, X. Association of age with the expression of hypoxia-inducible factors HIF-1α, HIF-2α, HIF-3α and VEGF in lung and heart of tibetan sheep. Animals 2019, 9, 673. [Google Scholar] [CrossRef]
- Aragonés, J.; Fraisl, P.; Baes, M.; Ca Rmeliet, P. Oxygen sensors at the crossroad of metabolism. Cell Metab. 2009, 9, 11–22. [Google Scholar] [CrossRef]
- McMahon, S.; Charbonneau, M.; Grandmont, S.; Richard, D.; Dubois, C. Transforming growth factor beta1 induces hypoxia-inducible factor-1 stabilization through selective inhibition of PHD2 expression. J. Biol. Chem. 2006, 281, 24171–24181. [Google Scholar] [CrossRef]
- Wang, H.; Huang, C.; Chen, N.; Zhu, K.; Chen, B.; Wang, W.; Wang, H. Molecular characterization and mRNA expression of HIF-prolyl hydroxylase-2 (phd2) in hypoxia-sensing pathways from Megalobrama amblycephala. Comp. Biochem. Physiol. Part B 2015, 186, 28–35. [Google Scholar] [CrossRef]
- Lin, X.H.; Ye, M.H.; Seatan, W.; Pan, Y.Y.; Chen, F.Y.; Huang, Y.; Zhu, C.H.; Li, G.L.; Tian, C.X. Sequence Characteristics and mRNA Expression Analysis of the phds Gene Family in Silver Sillago (Sillago sihama) under Hypoxia Stress. J. Guangdong Ocean Univ. 2020, 40, 1–8. [Google Scholar]
- Chen, N.; Huang, C.X.; Huang, C.H.; He, Y.; Chen, T.S.; Zhu, D.M.; Liu, H.; Wang, W.M.; Wang, H.L. The molecular characterization, expression pattern and alternative initiation of Megalobrama amblycephala Hif prolyl hydroxylase Phd1. Gene 2018, 678, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Huang, C.H.; Chen, B.X.; Liu, H.; Wang, W.M.; Gul, Y.; Wang, H.L. Alternative splicing transcription of Megalobrama amblycephala HIF prolyl hydroxylase PHD3 and up-regulation of PHD3 by HIF-1α. Biochem. Biophys. Res. Commun. 2016, 469, 737–742. [Google Scholar] [CrossRef]
- Li, X.; Li, F.; Zou, G.; Feng, C.; Sha, H.; Liu, S.; Liang, H. Physiological responses and molecular strategies in heart of silver carp (Hypophthalmichthys molitrix) under hypoxia and reoxygenation. Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 40, 100908. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Sun, J.L.; Zhao, L.L.; Wu, H.; Liu, Q.; Liao, L.; Luo, J.; Lian, W.Q.; Cui, C.; Jin, L.; Ma, J.D.; et al. Acute hypoxia changes the mode of glucose and lipid utilization in the liver of the largemouth bass (Micropterus salmoides). Sci. Total Environ. 2020, 713, 135157. [Google Scholar] [CrossRef]
- Zhang, G.; Yin, S.; Mao, J.; Liang, F.; Zhao, C.; Li, P.; Zhou, G.; Chen, S.; Tang, Z. Integrated analysis of mRNA-seq and miRNA-seq in the liver of Pelteobagrus vachelli in response to hypoxia. Sci. Rep. 2016, 6, 22907. [Google Scholar] [CrossRef] [PubMed]
- Pektas, S.; Knapp, M.J. Substrate preference of the HIF-prolyl hydroxylase-2 (PHD2) and substrate-induced conformational change. J. Inorg. Biochem. 2013, 126, 55–60. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Myllyharju, J. Prolyl 4-hydroxylases, key enzymes in the synthesis of collagens and regulation of the response to hypoxia, and their roles as treatment targets. Ann. Med. 2008, 40, 402–417. [Google Scholar] [CrossRef]
- Myllyharju, J. Prolyl 4-hydroxylases, master regulators of the hypoxia response. Acta Physiol. 2013, 208, 148–165. [Google Scholar] [CrossRef]
- Semenza, G.L.; Bels, D.D.; Corazza, F.; Kappler, M.; Eckert, A.W. Oxygen sensing, homeostasis, and disease. N. Engl. J. Med. 2011, 365, 537–547. [Google Scholar] [CrossRef]
- Minamishima, Y.A.; Moslehi, J.; Padera, R.F.; Bronson, R.T.; Liao, R.; Kaelin, W.G. A Feedback Loop Involving the Phd3 Prolyl Hydroxylase Tunes the Mammalian Hypoxic Response In Vivo. Mol. Cell. Biol. 2009, 29, 5729–5741. [Google Scholar] [CrossRef]
- Han, D.; Wen, L.; Chen, Y. Molecular Cloning of phd1 and Comparative Analysis of phd1, 2, and 3 Expression in Xenopus laevis. Sci. World J. 2012, 2012, 689287. [Google Scholar] [CrossRef]
- Moslehi, J.; Minamishima, Y.A.; Shi, J.; Neuberg, D.; Charytan, D.M.; Padera, R.F.; Signoretti, S.; Liao, R.; Kaelin, W.G. Loss of Hypoxia-Inducible Factor Prolyl Hydroxylase Activity in Cardiomyocytes Phenocopies Ischemic Cardiomyopathy. Circulation 2010, 122, 1004–1016. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.S. Characterization and comparative analysis of the EGLN gene family. Gene 2001, 275, 125–132. [Google Scholar] [CrossRef]
- Barth, S.; Edlich, F.; Berchner-Pfannschmidt, U.; Gneuss, S.; Camenisch, G. Hypoxia-inducible Factor Prolyl-4-hydroxylase PHD2 Protein Abundance Depends on Integral Membrane Anchoring of FKBP38. J. Biol. Chem. 2009, 284, 23046–23058. [Google Scholar] [CrossRef]
- Edurne, B.; Emmanuel, B.; Amandine, G.; Véronique, V.; Danièle, R.; Jacques, P. HIF prolyl-hydroxylase 2 is the key oxygen sensor setting low steady-state levels of HIF-1alpha in normoxia. EMBO J. 2003, 22, 4082–4090. [Google Scholar] [CrossRef]
- Shen, R.J.; Jiang, X.Y.; Pu, J.W.; Zou, S.M. HIF-1α and -2α genes in a hypoxia-sensitive teleost species Megalobrama amblycephala: cDNA cloning, expression and different responses to hypoxia. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2010, 157, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Lieb, M.E.; Menzies, K.; Moschella, M.C.; Ni, R.; Taubman, M.B. Mammalian EGLN genes have distinct patterns of mRNA expression and regulation. Biochem. Cell Biol. 2002, 80, 421–426. [Google Scholar] [CrossRef]
- Appelhoff, R.J.; Tian, Y.M.; Raval, R.R.; Turley, H.; Harris, A.L.; Pugh, C.W.; Ratcliffe, P.J.; Gleadle, J.M. Differential function of the prolyl hydroxylases PHD1, PHD2, and PHD3 in the regulation of hypoxia-inducible factor. J. Biol. Chem. 2004, 279, 38458–38465. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Cowan, A.; Fong, G.H. Essential role for prolyl hydroxylase domain protein 2 in oxygen homeostasis of the adult vascular system. Circulation 2007, 116, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Gu, J.; Li, L.; Liu, J.; Luo, B.; Cheung, H.W.; Boehm, J.S.; Ni, M.; Geisen, C.; Root, D.E.; et al. Control of cyclin D1 and breast tumorigenesis by the EglN2 prolyl hydroxylase. Cancer Cell 2009, 16, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Gillis, J.A.; Tidswell, O. The Origin of Vertebrate Gills. Curr. Biol. 2017, 27, 729–732. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Mao, J.; Liang, F.; Chen, J.; Zhao, C.; Yin, S.; Wang, L.; Tang, Z.; Chen, S. Modulated expression and enzymatic activities of Darkbarbel catfish, Pelteobagrus vachelli for oxidative stress induced by acute hypoxia and reoxygenation. Chemosphere 2016, 151, 271–279. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Names | Primer Sequences (5′–3′) | Application |
|---|---|---|
| PHD1-P-F | GCTTTTGAAGGAGTTGAAGAGTGGGT | CDS amplification |
| PHD1-P-R | GTCCGTCGCCGTTAGGATTGTC | |
| PHD2-P-F | GGGAAAATGGAGAACCTGATGAAGTG | |
| PHD2-P-R | TTGTGGCATAGGCTGGCTGGAC | |
| PHD3-P-F | CCGAGGCTACTTTTACGTGGATAATTT | |
| PHD3-P-R | GCATACCTTGTAGCATAGGACGGTTG | |
| UPM short | CTAATACGACTCACTATAGGGC | 5′ and 3′ RACE |
| UPM | CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT | |
| PHD1-GSP5-1 | ACCCATTTCAAGCCTCCTCCTTTT | 5′ RACE |
| PHD1-GSP5-2 | AAGTCCTGTCCCCGTGAGCATG | |
| PHD2-GSP5-1 | GCACGTCCTCCAGAATACTGCGTC | |
| PHD2-GSP5-2 | CACCCGCTTGTGCTTCTTCCAG | |
| PHD3-GSP5-1 | ATTTTATCCCCTCTGATGTTTGTCCTG | |
| PHD3-GSP5-2 | CCGTCGTTGAGAATCCCACAGTAA | |
| PHD1-GSP3-1 | TGGACAATCCTAACGGCGACG | 3′ RACE |
| PHD1-GSP3-2 | TGGGAGGGGAAGGGGCAGGGAGGTT | |
| PHD2-GSP3-1 | ATAACCCTAACGGAGATGG | |
| PHD2-GSP3-2 | CATTGAGCCCAAGTTTGA | |
| PHD3-GSP3-1 | GTGTGGGTCAACTGGGCAAAAGCAT | |
| PHD3-GSP3-2 | GCCGCTGCGTCACCTGTAT | |
| PHD1-Y-F | TCGGAAATGCCTAATGGACTG | RT-qPCR |
| PHD1-Y-R | GCTTTGTGCCCATTGTGATG | |
| PHD2-Y- F | CGACACTGTAACGGGAAACTGG | |
| PHD2-Y-R | TCTGGAAAGATCCGCAAAAGG | |
| PHD3-Y- F | AGTTCGAGACTTTGGCTGTC | |
| PHD3-Y-R | CCCCTCTGATGTTTGTCCTG | |
| β-actin F | GAACCCCAAAGCCAACAG | |
| β-actin R | CAGAGTCCATCACGATACCAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhang, M.; Ling, C.; Sha, H.; Zou, G.; Liang, H. Molecular Characterization and Response of Prolyl Hydroxylase Domain (PHD) Genes to Hypoxia Stress in Hypophthalmichthys molitrix. Animals 2022, 12, 131. https://doi.org/10.3390/ani12020131
Li X, Zhang M, Ling C, Sha H, Zou G, Liang H. Molecular Characterization and Response of Prolyl Hydroxylase Domain (PHD) Genes to Hypoxia Stress in Hypophthalmichthys molitrix. Animals. 2022; 12(2):131. https://doi.org/10.3390/ani12020131
Chicago/Turabian StyleLi, Xiaohui, Meidong Zhang, Chen Ling, Hang Sha, Guiwei Zou, and Hongwei Liang. 2022. "Molecular Characterization and Response of Prolyl Hydroxylase Domain (PHD) Genes to Hypoxia Stress in Hypophthalmichthys molitrix" Animals 12, no. 2: 131. https://doi.org/10.3390/ani12020131
APA StyleLi, X., Zhang, M., Ling, C., Sha, H., Zou, G., & Liang, H. (2022). Molecular Characterization and Response of Prolyl Hydroxylase Domain (PHD) Genes to Hypoxia Stress in Hypophthalmichthys molitrix. Animals, 12(2), 131. https://doi.org/10.3390/ani12020131