
Toxic Effects on Bioaccumulation, Hematological Parameters, Oxidative Stress, Immune Responses and Tissue Structure in Fish Exposed to Ammonia Nitrogen: A Review



 and
and 
Abstract
Simple Summary
Abstract
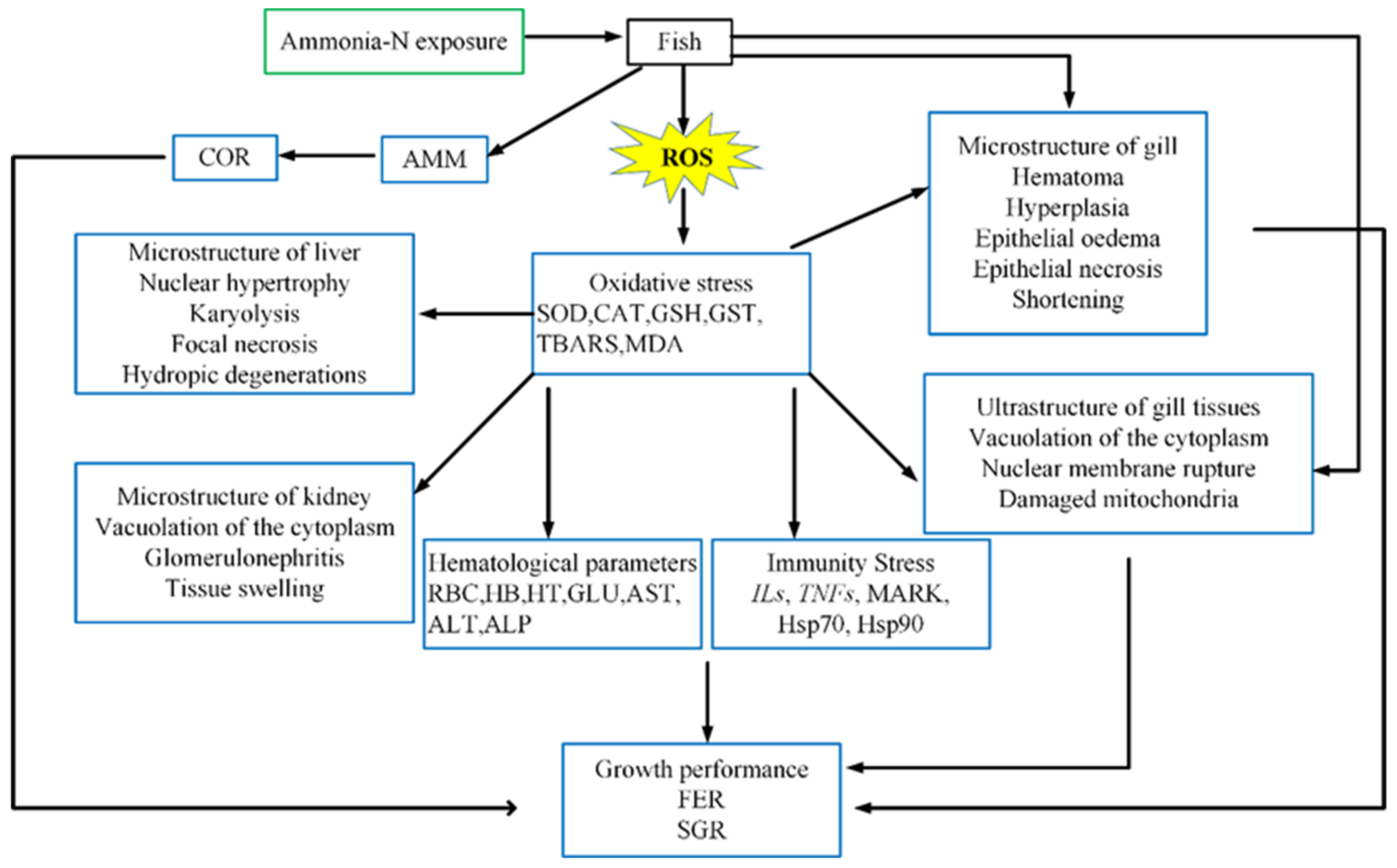
1. Introduction
2. Bioaccumulation
3. Hematological Parameters
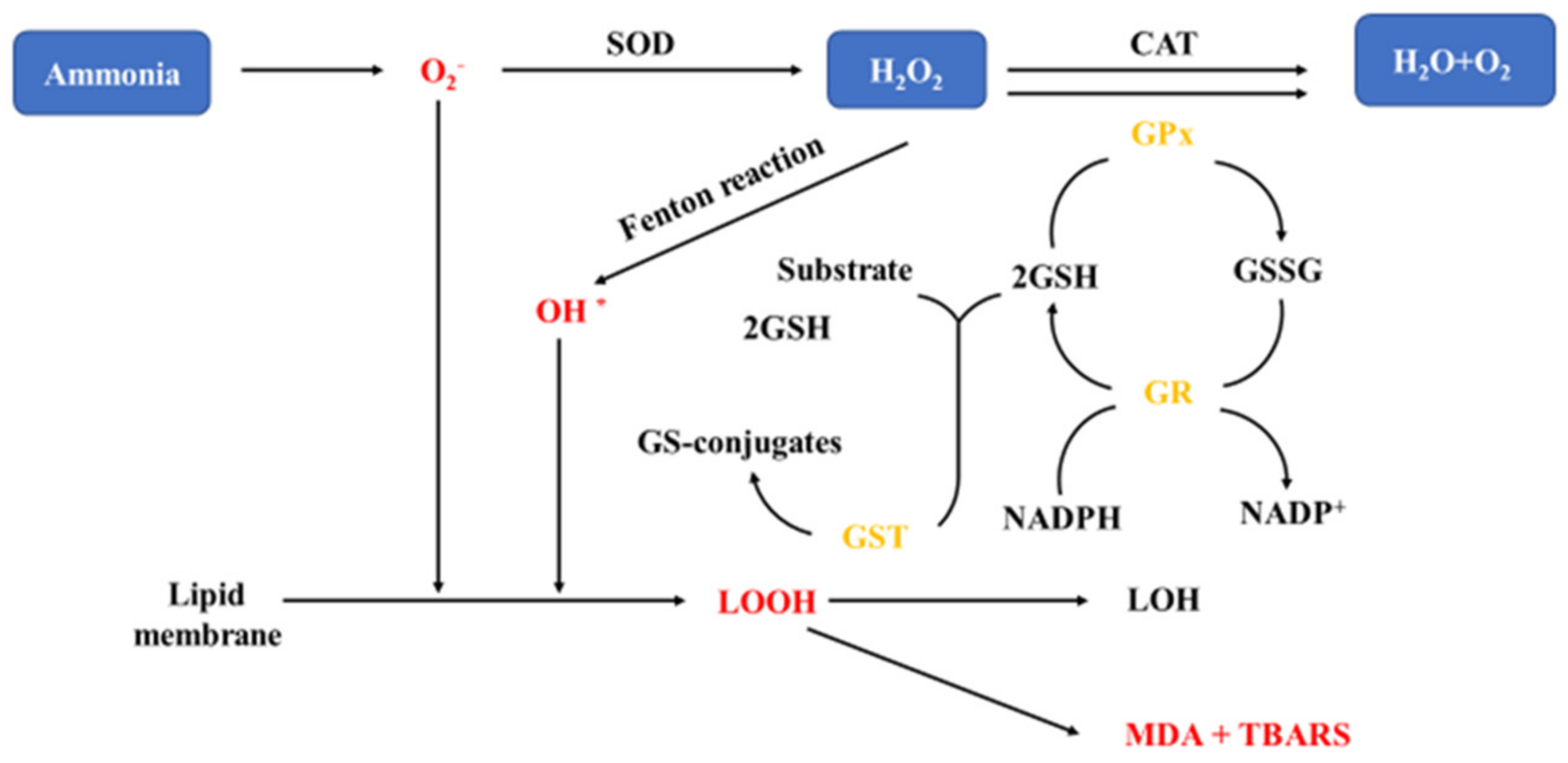
4. Oxidative Stress
5. Immune Response
6. Tissue Structure
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chang, Z.W.; Chiang, P.C.; Cheng, W.; Chang, C.C. Impact of ammonia exposure on coagulation in white shrimp, Litopenaeus vannamei. Ecotoxicol. Environ. Saf. 2015, 118, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Romano, N.; Zeng, C.S. Toxic effects of ammonia, nitrite, and nitrate to decapod crustaceans: A review on factors Influencing their toxicity, physiological consequences, and coping mechanisms. Rev. Fish. Sci. 2013, 21, 1–21. [Google Scholar] [CrossRef]
- Randall, D.J.; Tsui, T.K.N. Ammonia toxicity in fish. Mar. Pollut. Bull. 2002, 45, 17–23. [Google Scholar] [CrossRef]
- Eddy, F.B. Ammonia in estuaries and effects on fish. J. Fish Biol. 2005, 67, 1495–1513. [Google Scholar] [CrossRef]
- Armstrong, D.A.; Chippendale, D.; Knight, A.W.; Colt, J.E. Interaction of Ionized and un-Ionized ammonia on short-term survival and growth of prawn larvae, Macrobrachium rosenbergii. Biol. Bull. 1978, 154, 15–31. [Google Scholar] [CrossRef]
- Wang, L.S.; Xu, Q.Y.; Wang, C.A.; Li, J.N.; Chen, D.; Zhao, Z.G.; Luo, L.; Du, X. Effects of dietary α-ketoglutarate supplementation on the antioxidant defense system and HSP70 and HSP90 gene expression of hybrid sturgeon Acipenser schrenckii ♀ × Acipenser baerii ♂exposed to ammonia-N stress. Aquacult. Res. 2017, 48, 2266–2277. [Google Scholar] [CrossRef]
- Benli, A.C.K.; Koksal, G.; Ozkul, A. Sublethal ammonia exposure of Nile tilapia (Oreochromis niloticus L.): Effects on gill, liver and kidney histology. Chemosphere 2008, 72, 1355–1358. [Google Scholar] [CrossRef] [PubMed]
- Cong, M.; Wu, H.F.; Cao, T.F.; Ji, C.L.; Lv, J.S. Effects of ammonia nitrogen on gill mitochondria in clam Ruditapes philippinarum. Environ. Toxicol. Pharmacol. 2019, 65, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.Z.; Xue, M.Y.; Yang, S.B.; Zha, J.W.; Wang, G.X.; Ling, F. Ammonia exposure alters the expression of immune-related and antioxidant enzymes-related genes and the gut microbial community of crucian carp (Carassius auratus). Fish Shellfish Immunol. 2017, 70, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Li, L.H.; Qi, H.X. Effect of acute ammonia exposure on the glutathione redox system in FFRC strain common carp (Cyprinus carpio L.). Environ. Sci. Pollut. Res. 2019, 26, 27023–27031. [Google Scholar] [CrossRef]
- Zhang, W.; Xia, S.; Zhu, J.; Miao, L.; Ren, M.; Lin, Y.; Ge, X.; Sun, S. Growth performance, physiological response and histology changes of juvenile blunt snout bream, Megalobrama amblycephala exposed to chronic ammonia. Aquaculture 2019, 506, 424–436. [Google Scholar] [CrossRef]
- Yan, X.; Chen, Y.; Dong, X.; Tan, B.; Liu, H.; Zhang, S.; Chi, S.; Yang, Q.; Liu, H.; Yang, Y. Ammonia toxicity induces oxidative stress, inflammatory response and apoptosis in hybrid grouper (♀ Epinephelus fuscoguttatus × ♂ E. lanceolatus). Front. Mar. Sci. 2021, 8, 667432. [Google Scholar] [CrossRef]
- Liew, H.J.; Sinha, A.K.; Nawata, C.M.; Blust, R.; Wood, C.M.; de Boeck, G. Differential responses in ammonia excretion, sodium fluxes and gill permeability explain different sensitivities to acute high environmental ammonia in three freshwater teleosts. Aquat. Toxicol. 2013, 126, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Nakada, T.; Westhoff, C.M.; Kato, A.; Hirose, S. Ammonia secretion from fish gill depends on a set of Rh glycoproteins. FASEB J. 2007, 21, 1067–1074. [Google Scholar] [CrossRef]
- Dosdat, A.; Person-Le Ruyet, J.; Coves, D.; Dutto, G.; Gasset, E.; Le Roux, A.; Lemarie, G. Effect of chronic exposure to ammonia on growth, food utilisation and metabolism of the European sea bass (Dicentrarchus labrax). Aquat. Living Resour. 2003, 16, 509–520. [Google Scholar] [CrossRef]
- Lemarie, G.; Dosdat, A.; Coves, D.; Dutto, G.; Gasset, E.; Person-Le Ruyet, J. Effect of chronic ammonia exposure on growth of European seabass (Dicentrarchus labrax) juveniles. Aquaculture 2004, 229, 479–491. [Google Scholar] [CrossRef]
- Wilkie, M.P.; Pamenter, M.E.; Duquette, S.; Dhiyebi, H.; Sangha, N.; Skelton, G.; Smith, M.D.; Buck, L.T. The relationship between NMDA receptor function and the high ammonia tolerance of anoxia-tolerant goldfish. J. Exp. Biol. 2011, 214, 4107–4120. [Google Scholar] [CrossRef] [PubMed]
- Gao, N.; Zhu, L.; Guo, Z.; Yi, M.; Zhang, L. Effects of chronic ammonia exposure on ammonia metabolism and excretion in marine medaka Oryzias melastigma. Fish Shellfish. Immunol. 2017, 65, 226–234. [Google Scholar] [CrossRef]
- Smart, G.R. Investigation of the toxic mechanisms of ammonia to fish—gas exchange in rainbow trout (Salmo gairdneri) exposed to acutely lethal concentrations. J. Fish Biol. 1978, 12, 93–104. [Google Scholar] [CrossRef]
- Wee, N.L.J.; Tng, Y.Y.M.; Cheng, H.T.; Lee, S.M.L.; Chew, S.F.; Ip, Y.K. Ammonia toxicity and tolerance in the brain of the African sharptooth catfish, Clarias gariepinus. Aquat. Toxicol. 2007, 82, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, A.R.; Norenberg, M.D. The Na-K-Cl Co-transporter in astrocyte swelling. Metab. Brain Dis. 2010, 25, 31–38. [Google Scholar] [CrossRef]
- Marquez, J.; Cardona, C.; Campos-Sandoval, J.A.; Penalver, A.; Tosina, M.; Mates, J.M.; Martin-Rufian, M. Mammalian glutaminase isozymes in brain. Metab. Brain Dis. 2013, 28, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Rao, V.L.R. Nitric oxide in hepatic encephalopathy and hyperammonemia. Neurochem. Int. 2002, 41, 161–170. [Google Scholar] [CrossRef]
- Rodrigo, R.; Erceg, S.; Felipo, V. Neurons exposed to ammonia reproduce the differential alteration in nitric oxide modulation of guanylate cyclase in the cerebellum and cortex of patients with liver cirrhosis. Neurobiol. Dis. 2005, 19, 150–161. [Google Scholar] [CrossRef]
- Rodrigo, R.; Cauli, O.; Boix, J.; ElMlili, N.; Agusti, A.; Felipo, V. Role of NMDA receptors in acute liver failure and ammonia toxicity: Therapeutical implications. Neurochem. Int. 2009, 55, 113–118. [Google Scholar] [CrossRef]
- Braissant, O.; McLin, V.A.; Cudalbu, C. Ammonia toxicity to the brain. J. Inherit. Metab. Dis. 2013, 36, 595–612. [Google Scholar] [CrossRef] [PubMed]
- Chew, S.F.; Gan, J.; Ip, Y.K. Nitrogen metabolism and excretion in the swamp eel, Monopterus albus, during 6 or 40 days of estivation in mud. Physiol. Biochem. Zool. 2005, 78, 620–629. [Google Scholar] [CrossRef]
- Van der Linden, A.; Verhoye, M.; Nilsson, G.E. Does anoxia induce cell swelling in carp brains? In vivo MRI measurements in crucian carp and common carp. J. Neurophysiol. 2001, 85, 125–133. [Google Scholar] [CrossRef]
- Ip, Y.K.; Leong, M.W.F.; Sim, M.Y.; Goh, G.S.; Wong, W.P.; Chew, S.E. Chronic and acute ammonia toxicity in mudskippers, Periophthalmodon schlosseri and Boleophthalmus boddaerti: Brain ammonia and glutamine contents, and effects of methionine sulfoximine and MK801. J. Exp. Biol. 2005, 208, 1993–2004. [Google Scholar] [CrossRef] [PubMed]
- Hegazi, M.M.; Hasanein, S.S. Effects of chronic exposure to ammonia concentrations on brain monoamines and ATPases of Nile tilapia (Oreochromis niloticus). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 151, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.H.; Yang, F.F.; Ling, R.Z.; Liao, S.A.; Miao, Y.T.; Ye, C.X.; Wang, A.L. Effects of ammonia exposure on apoptosis, oxidative stress and immune response in pufferfish (Takifugu obscurus). Aquat. Toxicol. 2015, 164, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Z.; Li, M.; Wang, R.X.; Qian, Y.X. Effects of acute ammonia toxicity on oxidative stress, immune response and apoptosis of juvenile yellow catfish Pelteobagrus fulvidraco and the mitigation of exogenous taurine. Fish Shellfish. Immunol. 2018, 79, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Fazio, F.; Saoca, C.; Sanfilippo, M.; Capillo, G.; Spano, N.; Piccione, G. Response of vanadium bioaccumulation in tissues of Mugil cephalus (Linnaeus 1758). Sci. Total Environ. 2019, 689, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Fazio, F.; Saoca, C.; Ferranteili, V.; Cammilleri, G.; Capillo, G.; Piccione, G. Relationship between arsenic accumulation in tissues and hematological parameters in mullet caught in Faro Lake: A preliminary study. Environ. Sci. Pollut. Res. 2019, 26, 8821–8827. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, Y.J.; Kim, K.I.; Kim, S.K.; Kim, J.H. Toxic effects of nitrogenous compounds (ammonia, nitrite, and nitrate) on acute toxicity and antioxidant responses of juvenile olive flounder, Paralichthys olivaceus. Environ. Toxicol. Pharmacol. 2019, 67, 73–78. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. Effects of sub-chronic exposure to lead (Pb) and ascorbic acid in juvenile rockfish: Antioxidant responses, MT gene expression, and neurotransmitters. Chemosphere 2017, 171, 520–527. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. The chromium accumulation and its physiological effects in juvenile rockfish, Sebastes schlegelii, exposed to different levels of dietary chromium (Cr6+) concentrations. Environ. Toxicol. Pharmacol. 2016, 41, 152–158. [Google Scholar] [CrossRef]
- Yang, W.; Xiang, F.; Sun, H.; Chen, Y.; Minter, E.; Yang, Z. Changes in the selected hematological parameters and gill Na+/K+ ATPase activity of juvenile crucian carp Carassius auratus during elevated ammonia exposure and the post-exposure recovery. Biochem. Syst. Ecol. 2010, 38, 557–562. [Google Scholar] [CrossRef]
- Elbialy, Z.I.; Salah, A.S.; Elsheshtawy, A.; Rizk, M.; Abualreesh, M.H.; Abdel-Daim, M.M.; Salem, S.M.R.; Askary, A.E.; Assar, D.H. Exploring the multimodal role of yucca schidigera extract in protection against chronic ammonia exposure targeting: Growth, metabolic, stress and inflammatory responses in Nile iilapia (Oreochromis niloticus L.). Animals 2021, 11, 2072. [Google Scholar] [CrossRef]
- Gao, X.Q.; Fei, F.; Huang, B.; Meng, X.S.; Zhang, T.; Zhao, K.F.; Chen, H.B.; Xing, R.; Liu, B.L. Alterations in hematological and biochemical parameters, oxidative stress, and immune response in Takifugu rubripes under acute ammonia exposure. Comp. Biochem. Physiol. C Toxicol. Pharm. 2021, 243, 108978. [Google Scholar] [CrossRef]
- Kavitha, C.; Ramesh, M.; Kumaran, S.S.; Lakshmi, S.A. Toxicity of Moringa oleifera seed extract on some hematological and biochemical profiles in a freshwater fish, Cyprinus carpio. Exp. Toxicol. Pathol. 2012, 64, 681–687. [Google Scholar] [CrossRef]
- Praveena, M.; Sandeep, V.; Kavitha, N.; Jayantha Rao, K. Impact of tannery effluent, chromium on hematological parameters in a fresh water fish, Labeo Rohita (Hamilton). Res. J. Anim. Vet. Fish. Sci. 2013, 1, 1–5. [Google Scholar]
- Hoseini, S.M.; Yousefi, M.; Hoseinifar, S.H.; van doan, H. Antioxidant, enzymatic and hematological responses of common carp (Cyprinus carpio) fed with myrcene- or menthol-supplemented diets and exposed to ambient ammonia. Aquaculture 2019, 506, 246–255. [Google Scholar] [CrossRef]
- Zeitoun, M.M.; El-Azrak, K.E.-D.M.; Zaki, M.A.; Nemat-Allah, B.R.; Mehana, E.-S.E. Effects of ammonia toxicity on growth performance, cortisol, glucose and hematological response of Nile Tilapia (Oreochromis niloticus). Aceh J. Anim. Sci. 2016, 1, 21–28. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Tarkhani, R. Effect of short-term treatment with potassium permanganate on stress markers and blood biochemistry in goldfish Carassius auratus. Aquacult. Res. 2013, 44, 869–875. [Google Scholar] [CrossRef]
- David, M.; Mushigeri, S.B.; Shivakumar, R.; Philip, G.H. Response of Cyprinus carpio (Linn) to sublethal concentration of cypermethrin: Alterations in protein metabolic profiles. Chemosphere 2004, 56, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Asthana, S.; Fatma, F. Effect of ammonia inhalation on serum protein of Albino rat. Natl. Acad. Sci. Lett. India 2008, 31, 117–119. [Google Scholar]
- Kim, J.H.; Cho, J.H.; Kim, S.R.; Hur, Y.B. Toxic effects of waterborne ammonia exposure on hematological parameters, oxidative stress and stress indicators of juvenile hybrid grouper, Epinephelus lanceolatus ♂× Epinephelus fuscoguttatus ♀. Environ. Toxicol. Pharmacol. 2020, 80, 103453. [Google Scholar] [CrossRef] [PubMed]
- Das, P.C.; Ayyappan, S.; Jena, J.K.; Das, B.K. Acute toxicity of ammonia and its sub-lethal effects on selected haematological and enzymatic parameters of mrigal, Cirrhinus mrigala (Hamilton). Aquacult. Res. 2004, 35, 134–143. [Google Scholar] [CrossRef]
- Iheanacho, S.C.; Odo, G.E. Neurotoxicity, oxidative stress biomarkers and haematological responses in African catfish (Clarias gariepinus) exposed to polyvinyl chloride microparticles. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2020, 232, 10. [Google Scholar] [CrossRef]
- Roda, J.F.B.; Lauer, M.M.; Risso, W.E.; Martinez, C.B.D. Microplastics and copper effects on the neotropical teleost Prochilodus lineatus: Is there any interaction? Comp. Biochem. Physiol. A-Mol. Integr. Physiol. 2020, 242, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Peng, K.; Wang, G.; Mo, W.; Huang, Y.; Cao, J. Metabolic changes, antioxidant status, immune response and resistance to ammonia stress in juvenile yellow catfish (Pelteobagrus fulvidraco) fed diet supplemented with sodium butyrate. Aquaculture 2021, 536, 736441. [Google Scholar] [CrossRef]
- Cui, Y.; Ren, X.; Li, J.; Zhai, Q.; Feng, Y.; Xu, Y.; Ma, L. Effects of ammonia-N stress on metabolic and immune function via the neuroendocrine system in Litopenaeus vannamei. Fish Shellfish. Immunol. 2017, 64, 270–275. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. Changes in hematological parameters, plasma cortisol, and acetylcholinesterase of juvenile rockfish, Sebastes schlegelii supplemented with the dietary ascorbic acid. Aquacult. Rep. 2016, 4, 80–85. [Google Scholar] [CrossRef][Green Version]
- Habte-Tsion, H.M.; Liu, B.; Ge, X.P.; Xie, J.; Xu, P.; Ren, M.C.; Zhou, Q.L.; Pan, L.K.; Chen, R.L. Effects of dietary protein level on growth performance, muscle composition, blood composition, and digestive enzyme activity of wuchang bream (Megalobrama amblycephala) fry. Isr. J. Aquacult. Bamidgeh. 2013, 65, 9. [Google Scholar]
- Peyghan, R.; Takamy, G.A. Histopathological, serum enzyme, cholesterol and urea changes in experimental acute toxicity of ammonia in common carp Cyprinus carpio and use of natural zeolite for prevention. Aquac. Int. 2002, 10, 317–325. [Google Scholar] [CrossRef]
- Barbieri, E.; Bondioli, A.C.V. Acute toxicity of ammonia in Pacu fish (Piaractus mesopotamicus, Holmberg, 1887) at different temperatures levels. Aquac. Res. 2015, 46, 565–571. [Google Scholar] [CrossRef]
- Mirghaed, A.T.; Fayaz, S.; Hoseini, S.M. Effects of dietary 1,8-cineole supplementation on serum stress and antioxidant markers of common carp (Cyprinus carpio) acutely exposed to ambient ammonia. Aquaculture 2019, 509, 8–15. [Google Scholar] [CrossRef]
- Xing, X.D.; Li, M.; Yuan, L.X.; Song, M.Z.; Ren, Q.Y.; Shi, G.; Meng, F.X.; Wang, R.X. The protective effects of taurine on acute ammonia toxicity in grass carp Ctenopharynodon idellus. Fish Shellfish Immunol. 2016, 56, 517–522. [Google Scholar] [CrossRef]
- Murthy, C.R.K.; Rao, K.V.R.; Bai, G.; Norenberg, M.D. Ammonia-induced production of free radicals in primary cultures of rat astrocytes. J. Neurosci. Res. 2001, 66, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Antioxidant defence mechanisms: From the beginning to the end (of the beginning). Free. Radic. Res. 1999, 31, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Yanaka, A.; Shibahara, T.; Matsui, H.; Nakahara, A.; Tanaka, N.; Muto, H.; Momoi, T.; Uchiyama, Y. Ammonia-induced apoptosis is accelerated at higher pH in gastric surface mucous cells. Am. J. Physiol. Gastroint. Liver Physiol. 2002, 283, G986–G995. [Google Scholar] [CrossRef] [PubMed]
- Madeira, D.; Narciso, L.; Cabral, H.N.; Vinagre, C.; Diniz, M.S. Influence of temperature in thermal and oxidative stress responses in estuarine fish. Comp. Biochem. Physiol. A-Mol. Integr. Physiol. 2013, 166, 237–243. [Google Scholar] [CrossRef]
- Rama, S.; Manjabhat, S.N. Protective effect of shrimp carotenoids against ammonia stress in common carp, Cyprinus carpio. Ecotoxicol. Environ. Saf. 2014, 107, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, D.R. Contaminant-stimulated reactive oxygen species production and oxidative damage in aquatic organisms. Mar. Pollut. Bull. 2001, 42, 656–666. [Google Scholar] [CrossRef]
- Xu, K.H.; Zhang, Y.D.; Huang, Y.M.; Wang, J. Toxicological effects of microplastics and phenanthrene to zebrafish (Danio rerio). Sci. Total Environ. 2021, 757, 8. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. Oxidative stress, neurotoxicity, and non-specific immune responses in juvenile red sea bream, Pagrus major, exposed to different waterborne selenium concentrations. Chemosphere 2015, 135, 46–52. [Google Scholar] [CrossRef]
- Gurer-Orhan, H.; Sabir, H.U.; Ozgunes, H. Correlation between clinical indicators of lead poisoning and oxidative stress parameters in controls and lead-exposed workers. Toxicology 2004, 195, 147–154. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. Oxidative stress, neurotoxicity, and metallothionein (MT) gene expression in juvenile rock fish Sebastes schlegelii under the different levels of dietary chromium (Cr6+) exposure. Ecotoxicol. Environ. Saf. 2016, 125, 78–84. [Google Scholar] [CrossRef]
- Chen, D.C.; Ning, F.Y.; Zhang, J.Y.; Tang, Y.; Teng, X.H. NF-kappa B pathway took part in the development of apoptosis mediated by miR-15a and oxidative stress via mitochondrial pathway in ammonia-treated chicken splenic lymphocytes. Sci. Total Environ. 2020, 729, 9. [Google Scholar] [CrossRef]
- Han, Q.; Zhang, J.Y.; Sun, Q.; Xu, Y.M.; Teng, X.H. Oxidative stress and mitochondrial dysfunction involved in ammonia-induced nephrocyte necroptosis in chickens. Ecotoxicol Environ. Saf. 2020, 203, 9. [Google Scholar] [CrossRef]
- Liu, C.; Wu, F.C.; Que, H.Y.; Zhang, G.F. Relationships of growth and mortality to enzymatic activity, and the relative mRNA expression of cultured scallops Patinopecten yessoensis in the Yellow Sea, China. J. Oceanol. Limnol. 2019, 37, 1409–1422. [Google Scholar] [CrossRef]
- Hang, W.W.; Du, M.R.; Fang, J.G.; Gao, Y.P.; Mao, Y.Z.; Chen, Q.L.; Lin, F.; Jiang, Z.J. Response of Yesso scallop Patinopecten yessoensis to acute temperature challenge: Physiological and biochemical parameters. J. Oceanol. Limnol. 2019, 37, 321–329. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, J.H.; Park, M.A.; Hwang, S.D.; Kang, J.C. The toxic effects of ammonia exposure on antioxidant and immune responses in Rockfish, Sebastes schlegelii during thermal stress. Environ. Toxicol. Pharmacol. 2015, 40, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.K.; AbdElgawad, H.; Giblen, T.; Zinta, G.; de Rop, M.; Asard, H.; Blust, R.; de Boeck, G. Anti-oxidative defences are modulated differentially in three freshwater teleosts in response to ammonia-induced oxidative stress. PLoS ONE 2014, 9, 19. [Google Scholar] [CrossRef]
- Sinha, A.K.; AbdElgawad, H.; Zinta, G.; Dasan, A.F.; Rasoloniriana, R.; Asard, H.; Blust, R.; de Boeck, G. Nutritional status as the key modulator of antioxidant responses induced by high environmental ammonia and salinity stress in European sea bass (Dicentrarchus labrax). PLoS ONE 2015, 10, 29. [Google Scholar] [CrossRef]
- Lin, Y.C.; Chen, J.C. Acute toxicity of ammonia on Litopenaeus vannamei Boone juveniles at different salinity levels. J. Exp. Mar. Biol. Ecol. 2001, 259, 109–119. [Google Scholar] [CrossRef]
- Wang, J.; Li, J.J.; Xu, N.; Li, J.; Li, Z.H.; Chen, Y.F.; Yang, Z. Responses of Takifugu obscurus fertilized eggs and larvae to increased ammonia exposure. Environ. Sci. Pollut. Res. 2015, 22, 15976–15984. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yu, N.; Qin, J.G.; Li, E.C.; Du, Z.Y.; Chen, L.Q. Effects of ammonia stress, dietary linseed oil and Edwardsiella ictaluri challenge on juvenile darkbarbel catfish Pelteobagrus vachelli. Fish Shellfish. Immunol. 2014, 38, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.J.; Lu, K.; Minter, E.J.A.; Chen, Y.F.; Yang, Z.; Montagnes, D.J.S. Combined effects of ammonia and microcystin on survival, growth, antioxidant responses, and lipid peroxidation of bighead carp Hypophthalmythys nobilis larvae. J. Hazard. Mater. 2012, 221, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Yu, Y.B.; Choi, J.H. Toxic effects on bioaccumulation, hematological parameters, oxidative stress, immune responses and neurotoxicity in fish exposed to microplastics: A review. J. Hazard Mater. 2021, 413, 125423. [Google Scholar] [CrossRef]
- Li, C.H.; Ni, D.J.; Song, L.S.; Zhao, J.; Zhang, H.; Li, L. Molecular cloning and characterization of a catalase gene from Zhikong scallop Chlamys farreri. Fish Shellfish. Immunol. 2008, 24, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.Q.; Lin, J.W.; Han, Y.; Han, Y. Ammonia stress-induced apoptosis by p53-BAX/BCL-2 signal pathway in hepatopancreas of common carp (Cyprinus carpio). Aquac. Int. 2021, 29, 1895–1907. [Google Scholar] [CrossRef]
- Zhang, T.; Yan, Z.; Zheng, X.; Wang, S.; Fan, J.; Liu, Z. Effects of acute ammonia toxicity on oxidative stress, DNA damage and apoptosis in digestive gland and gill of Asian clam (Corbicula fluminea). Fish Shellfish. Immunol. 2020, 99, 514–525. [Google Scholar] [CrossRef]
- Wang, X.Q.; Wang, L.L.; Yao, C.; Qiu, L.M.; Zhang, H.; Zhi, Z.; Song, L.S. Alternation of immune parameters and cellular energy allocation of Chlamys farreri under ammonia-N exposure and Vibrio anguillarum challenge. Fish Shellfish. Immunol. 2012, 32, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Su, Y.; Lin, R.Z.; Shi, H.H.; Wang, X.R. 2-Chlorophenol induced ROS generation in fish Carassius auratus based on the EPR method. Chemosphere 2006, 65, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Hegazi, M.M.; Attia, Z.I.; Ashour, O.A. Oxidative stress and antioxidant enzymes in liver and white muscle of Nile tilapia juveniles in chronic ammonia exposure. Aquat. Toxicol. 2010, 99, 118–125. [Google Scholar] [CrossRef]
- Maltez, L.C.; Stringhetta, G.R.; Enamorado, A.D.; Okamoto, M.H.; Romano, L.A.; Monserrat, J.M.; Sampaio, L.A.; Garcia, L. Ammonia exposure and subsequent recovery trigger oxidative stress responses in juveniles of Brazilian flounder Paralichthys orbignyanus. Fish Physiol. Biochem. 2017, 43, 1747–1759. [Google Scholar] [CrossRef]
- Ponton, D.E.; Caron, A.; Hare, L.; Campbell, P.G.C. Hepatic oxidative stress and metal subcellular partitioning are affected by selenium exposure in wild yellow perch (Perca flavescens). Environ. Pollut. 2016, 214, 608–617. [Google Scholar] [CrossRef]
- Li, Z.H.; Zlabek, V.; Velisek, J.; Grabic, R.; Machova, J.; Randak, T. Modulation of antioxidant defence system in brain of rainbow trout (Oncorhynchus mykiss) after chronic carbamazepine treatment. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 151, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Antonio-Garcia, M.T.; Masso-Gonzalez, E.L. Toxic effects of perinatal lead exposure on the brain of rats: Involvement of oxidative stress and the beneficial role of antioxidants. Food Chem. Toxicol. 2008, 46, 2089–2095. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Gong, S.; Li, Q.; Yuan, L.; Meng, F.; Wang, R. Ammonia toxicity induces glutamine accumulation, oxidative stress and immunosuppression in juvenile yellow catfish Pelteobagrus fulvidraco. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2016, 183–184, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Almroth, B.C.; Sturve, J.; Berglund, A.; Forlin, L. Oxidative damage in eelpout (Zoarces viviparus), measured as protein carbonyls and TBARS, as biomarkers. Aquat. Toxicol. 2005, 73, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar]
- Barhoumi, B.; Clerandeau, C.; Gourves, P.Y.; Le Menach, K.; El Megdiche, Y.; Peluhet, L.; Budzinski, H.; Baudrimont, M.; Driss, M.R.; Cachot, J. Pollution biomonitoring in the Bizerte lagoon (Tunisia), using combined chemical and biomarker analyses in grass goby, Zosterisessor ophiocephalus (Teleostei, Gobiidae). Mar. Environ. Res. 2014, 101, 184–195. [Google Scholar] [CrossRef]
- Zhang, Z.W.; Liu, Q.; Cai, J.Z.; Yang, J.; Shen, Q.; Xu, S.W. Chlorpyrifos exposure in common carp (Cyprinus carpio L.) leads to oxidative stress and immune responses. Fish Shellfish. Immunol. 2017, 67, 604–611. [Google Scholar] [CrossRef]
- Al-Ghanim, K.A.; Ahmad, Z.; Al-Balawi, H.F.A.; Al-Misned, F.; Maboob, S.; Suliman, E.M. Effects of a low-radiotoxicity uranium salt (uranyl acetate) on biochemical and hematological parameters of the catfish, Clarias gariepinus. Chin. J. Oceanol. Limnol. 2016, 34, 109–117. [Google Scholar] [CrossRef]
- Jia, R.; Liu, B.L.; Han, C.; Huang, B.; Lei, J.L. Effects of ammonia exposure on stress and immune response in juvenile turbot (Scophthalmus maximus). Aquac. Res. 2017, 48, 3149–3162. [Google Scholar] [CrossRef]
- Li, M.; Zhang, M.Z.; Qian, Y.X.; Shi, G.; Wang, R.X. Ammonia toxicity in the yellow catfish (Pelteobagrus fulvidraco): The mechanistic insight from physiological detoxification to poisoning. Fish Shellfish. Immunol. 2020, 102, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Svobodova, Z.; Vykusova, B. The effects of pollutants on selected haematological and biochemical parameters in fish. Biomédica 1994, 31, 209–215. [Google Scholar]
- He, K.; Luo, X.; Wen, M.; Wang, C.; Qin, C.; Shao, J.; Gan, L.; Dong, R.; Jiang, H. Effect of acute ammonia toxicity on inflammation, oxidative stress and apoptosis in head kidney macrophage of Pelteobagrus fulvidraco and the alleviation of curcumin. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 248, 109098. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wu, X.Q.; Zheng, L.J.; Dai, Z.Y.; Wu, L.F. Effect of acute exposure to ammonia and BFT alterations on Rhynchocypris lagowski: Digestive enzyme, inflammation response, oxidative stress and immunological parameters. Environ. Toxicol. Pharmacol. 2020, 78, 11. [Google Scholar] [CrossRef] [PubMed]
- Savan, R.; Sakai, M. Genomics of fish cytokines. Comp. Biochem. Physiol. D Genom. Proteom. 2006, 1, 89–101. [Google Scholar] [CrossRef]
- Dai, J.; Zhang, L.B.; Du, X.Y.; Zhang, P.J.; Li, W.; Guo, X.Y.; Li, Y.H. Effect of lead on antioxidant ability and immune responses of Crucian Carp. Biol. Trace Elem. Res. 2018, 186, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Marshall, W.S.; Ossum, C.G.; Hoffmann, E.K. Hypotonic shock mediation by p38 MAPK, JNK, PKC, FAK, OSR1 and SPAK in osmosensing chloride secreting cells of killifish opercular epithelium. J. Exp. Biol. 2005, 208, 1063–1077. [Google Scholar] [CrossRef] [PubMed]
- Feidantsis, K.; Portner, H.O.; Markou, T.; Lazou, A.; Michaelidis, B. Involvement of p38 MAPK in the induction of Hsp70 during acute thermal stress in red blood cells of the gilthead sea bream, Sparus aurata. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2012, 317A, 303–310. [Google Scholar] [CrossRef]
- Cao, J.L.; Chen, J.J.; Wang, J.D.; Klerks, P.; Xie, L.T. Effects of sodium fluoride on MAPKs signaling pathway in the gills of a freshwater teleost, Cyprinus carpio. Aquat. Toxicol. 2014, 152, 164–172. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Bortoletti, M.; Olivotto, I.; Ratti, S.; Poltronieri, C.; Negrato, E.; Caberlotto, S.; Radaelli, G.; Bertotto, D. Salinity, temperature and ammonia acute stress response in seabream (Sparus aurata) juveniles: A multidisciplinary study. Animals 2021, 11, 97. [Google Scholar] [CrossRef]
- Deng, Z.; Sun, S.; Gao, T.; Han, Z. The Hsp70 gene family in boleophthalmus pectinirostris: Genome-wide identification and expression analysis under high ammonia stress. Animals 2019, 9, 36. [Google Scholar] [CrossRef]
- Sivakumar, R.; Jayaprakash, M.; Natesan, M. Effects of heavy metals on antioxidants and expression of HSP70 in different tissues of Milk fish (Chanos chanos) of Kaattuppalli Island, Chennai, India. Ecotoxicol. Environ. Saf. 2013, 98, 8–18. [Google Scholar]
- Rodrigues, R.V.; Romano, L.A.; Schwarz, M.H.; Delbos, B.; Sampaio, L.A. Acute tolerance and histopathological effects of ammonia on juvenile maroon clownfish Premnas biaculeatus (Block 1790). Aquacult. Res. 2014, 45, 1133–1139. [Google Scholar] [CrossRef]
- Le Ruyet, J.P.; Boeuf, G.; Infante, J.Z.; Helgason, S.; Le Roux, A. Short-term physiological changes in turbot and seabream juveniles exposed to exogenous ammonia. Comp. Biochem. Physiol. A-Mol. Integr. Physiol. 1998, 119, 511–518. [Google Scholar] [CrossRef]
- Van der Oost, R.; Beyer, J.; Vermeulen, N.P.E. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Bernet, D.; Schmidt, H.; Meier, W.; Burkhardt-Holm, P.; Wahli, T. Histopathology in fish: Proposal for a protocol to assess aquatic pollution. J. Fish Dis. 1999, 22, 25–34. [Google Scholar] [CrossRef]
- Ortiz, J.B.; de Canales, M.L.G.; Sarasquete, C. Histopathological changes induced by lindane (gamma-HCH) in various organs of fishes. Sci. Mar. 2003, 67, 53–61. [Google Scholar]
- Lease, H.M.; Hansen, J.A.; Bergman, H.L.; Meyer, J.S. Structural changes in gills of lost river suckers exposed to elevated pH and ammonia concentrations. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2003, 134, 491–500. [Google Scholar] [CrossRef]
- Kolarevic, J.; Selset, R.; Felip, O.; Good, C.; Snekvik, K.; Takle, H.; Ytteborg, E.; Baeverfjord, G.; Asgard, T.; Terjesen, B.F. Influence of long term ammonia exposure on Atlantic salmon (Salmo salar L.) parr growth and welfare. Aquacult. Res. 2013, 44, 1649–1664. [Google Scholar] [CrossRef]
- Miron, D.D.; Moraes, B.; Becker, A.G.; Crestani, M.; Spanevello, R.; Loro, V.L.; Baldisserotto, B. Ammonia and pH effects on some metabolic parameters and gill histology of silver catfish, Rhamdia quelen (Heptapteridae). Aquaculture 2008, 277, 192–196. [Google Scholar] [CrossRef]
- Zhang, W.; Sun, S.; Ge, X.; Xia, S.; Zhu, J.; Miao, L.; Lin, Y.; Liang, H.; Pan, W.; Su, Y.; et al. Acute effects of ammonia exposure on the plasma and haematological parameters and histological structure of the juvenile blunt snout bream, Megalobrama amblycephala, and post-exposure recovery. Aquacult. Res. 2018, 49, 1008–1019. [Google Scholar] [CrossRef]
- Rodrigues, R.V.; Schwarz, M.H.; Delbos, B.C.; Carvalho, E.L.; Romano, L.A.; Sampaio, L.A. Acute exposure of juvenile cobia Rachycentron canadum to nitrate induces gill, esophageal and brain damage. Aquaculture 2011, 322, 223–226. [Google Scholar] [CrossRef]
- Pesce, S.F.; Cazenave, J.; Monferran, M.V.; Frede, S.; Wunderlin, D.A. Integrated survey on toxic effects of lindane on neotropical fish: Corydoras paleatus and Jenynsia multidentata. Environ. Pollut. 2008, 156, 775–783. [Google Scholar] [CrossRef]
- Tang, C.H.; Hwang, L.Y.; Shen, I.D.; Chiu, Y.H.; Lee, T.H. Immunolocalization of chloride transporters to gill epithelia of euryhaline teleosts with opposite salinity-induced Na+/K+-ATPase responses. Fish Physiol. Biochem. 2011, 37, 709–724. [Google Scholar] [CrossRef]
- Williams, T.A.; Bonham, L.A.; Bernier, N.J. High environmental ammonia exposure has developmental-stage specific and long-term consequences on the cortisol stress response in zebrafish. Gen. Comp. Endocrinol. 2017, 254, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Mooney, T.J.; Pease, C.; Trenfield, M.; van Dam, R.; Harford, A.J. Modeling the ph-ammonia toxicity relationship for hydra viridissima in soft waters with low Ionic concentrations. Environ. Toxicol. Chem. 2018, 37, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Souza-Bastos, L.R.; Val, A.L.; Wood, C.M. Are Amazonian fish more sensitive to ammonia? Toxicity of ammonia to eleven native species. Hydrobiologia 2017, 789, 143–155. [Google Scholar] [CrossRef]
- Ravindrababu, G.; Neeraja, P. Histological changes in certain tissues of fish on ambient ammonia stress and post ammonia state (recovery). Int. J. Adv. Biotechnol. Res. 2012, 2, 430–435. [Google Scholar]



{kind=link}
{kind=link}
| Exposure Route | Exposure Type | Fish Specie | Ammonia Concentration | Exposure Time | Response Concentration | Response * | Reference |
|---|---|---|---|---|---|---|---|
| RBC (Billion/mL) | |||||||
| Sea water | Waterborne exposure | Takifugu rubripes | 5, 50, 100, 150 mg/L | 24, 48, 96 h | 100, 150 mg/L | − | Gao et al. [40] 2021 |
| Freshwater | Waterborne exposure | Cyprinus carpio | 106mg/L | 24 h | 106 mg/L | − | Hoseini et al. [43] 2019 |
| Megalobrama amblycephala | 5, 10, 15, 20 mg/L | 9 weeks | 20 mg/L | − | Zhang et al. [11] 2019 | ||
| Ht (%) | |||||||
| Sea water | Waterborne exposure | Takifugu rubripes | 5, 50, 100, 150 mg/L | 24, 48, 96 h | 50, 100, 150 mg/L | − | Gao et al. [40] 2021 |
| Piaractus mesopotamicus | 1, 2, 3 mg/L | 96 h | 2, 3 mg/L | + | Edison et al. [57] 2015 | ||
| Freshwater | Waterborne exposure | Cyprinus carpio | 106 mg/L | 24 h | 10 6mg/L | − | Hoseini et al. [43] 2019 |
| Megalobrama amblycephala | 5, 10, 15, 20 mg/L | 9 weeks | 20 mg/L | − | Zhang et al. [11] 2019 | ||
| Hb (g/L) | |||||||
| Sea water | Waterborne exposure | Takifugu rubripes | 5, 50, 100, 150 mg/L | 24, 48, 96 h | 50, 100, 150 mg/L | − | Gao et al. [40] 2021 |
| Piaractus mesopotamicus | 1, 2, 3 mg/L | 96 h | 2, 3 mg/L | − | Edison et al. [57] 2015 | ||
| Freshwater | Waterborne exposure | Cyprinus carpio | 106 mg/L | 24 h | 106 mg/L | − | Hoseini et al. [43] 2019 |
| Megalobrama amblycephala | 5, 10, 15, 20 mg/L | 9 weeks | 20 mg/L | − | Zhang et al. [11] 2019 | ||
| Glucose (mg/dL) | |||||||
| Sea water | Waterborne exposure | Takifugu rubripes | 5, 50, 100, 150 mg/L | 24, 48, 96 h | 50, 100, 150 mg/L | + | Gao et al. [40] 2021 |
| Litopenaeus vannamei | 0.32, 0.44, 0.60 mg/L | 6 h, 12 h, 1 day, 2 days | 0.32, 0.44, 0.60 mg/L | + | Cui et al. [53] 2017 | ||
| Piaractus mesopotamicus | 1, 2, 3 mg/L | 96 h | 2, 3 mg/L | + | Edison et al. [57] 2015 | ||
| Freshwater | Waterborne exposure | Pelteobagrus fulvidraco | 100 mg/L | 24, 48, 72 h | 100 mg/L | + | Zhao et al. [52] 2021 |
| Megalobrama amblycephala | 5, 10, 15, 20 mg/L | 9 weeks | 20 mg/L | − | Zhang et al. [11] 2019 | ||
| Cyprinus carpio | 0.5 mg/L | 24 h | 0.5 mg/L | + | Mirghaed et al. [58] 2019 | ||
| Total protein (g/dL) | |||||||
| Sea water | Waterborne exposure | Takifugu rubripes | 5, 50, 100, 150 mg/L | 24, 48, 96 h | 50, 100, 150 mg/L | − | Gao et al. [40] 2021 |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | 1, 2, 4, 8 mg/L | 1week, 2 weeks | 8 mg/L | − | Kim et al. [48] 2020 | ||
| Freshwater | Waterborne exposure | Pelteobagrus fulvidraco | 100 mg/L | 24, 48, 72 h | 100 mg/L | + | Zhao et al. [52] 2021 |
| Inject | Ctenopharynodon idellus | 9 μL | 96 h | 9 μL | × | Xing et al. [59] 2016 | |
| AST (U/L) | |||||||
| Freshwater | Waterborne exposure | Pelteobagrus fulvidraco | 100 mg/L | 24, 48, 72 h | 100 mg/L | + | Zhao et al. [52] 2021 |
| Megalobrama amblycephala | 5, 10, 15, 20 mg/L | 9 weeks | 20 mg/L | + | Zhang et al. [11] 2019 | ||
| Cyprinus carpio | 106 mg/L | 24 h | 106 mg/L | + | Hoseini et al. [43] 2019 | ||
| ALT (U/L) | |||||||
| Sea water | Waterborne exposure | Takifugu rubripes | 5, 50, 100, 150 mg/L | 24, 48, 96 h | 50, 100, 150 mg/L | + | Gao et al. [40] 2021 |
| Freshwater | Waterborne exposure | Pelteobagrus fulvidraco | 100 mg/L | 24, 48, 72 h | 100 mg/L | + | Zhao et al. [52] 2021 |
| Megalobrama amblycephala | 5, 10, 15, 20 mg/L | 9 weeks | 20 mg/L | × | Zhang et al. [11] 2019 | ||
| Cyprinus carpio | 106 mg/L | 24 h | 106 mg/L | + | Hoseini et al. [43] 2019 | ||
| ALP (U/L) | |||||||
| Freshwater | Waterborne exposure | Pelteobagrus fulvidraco | 100 mg/L | 24, 48, 72 h | 100 mg/L | × | Zhao et al. [52] 2021 |
| Megalobrama amblycephala | 5, 10, 15, 20 mg/L | 9 weeks | 20 mg/L | × | Zhang et al. [11] 2019 | ||
| Cyprinus carpio | 106 mg/L | 24 h | 106 mg/L | + | Hoseini et al. [43] 2019 | ||
| Exposure Route | Exposure Type | Fish Specie | Ammonia Concentration | Exposure Periods | Response Concentration | Target Organs | Response * | Reference |
|---|---|---|---|---|---|---|---|---|
| SOD (Superoxide dismutase) | ||||||||
| Sea water | Waterborne exposure | Dicentrarchus labrax | 20 mg/L | 12, 48, 84, 180 h | 20 mg/L | Blood | × | Sinha et al. [76] 2015 |
| Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂ | 1, 2, 4, 8 mg/L | 1week, 2 wk | 4, 8 mg/L | Liver, Gill | + | Kim et al. [48] 2020 | ||
| Scophthalmus maximus | 5, 20, 40 mg/L | 24, 48, 96 h | 20, 40 mg/L | Liver | + | Jia et al. [98] 2020 | ||
| Chlamys farreri | 20 mg/L | 1, 12, 24 d | 20 mg/L | Blood | + | Wang et al. [85] 2012 | ||
| Takifugu rubripes | 5, 50, 100, 150 mg/L | 24 h | 50, 100, 150 mg/L | Gill | + | Gao et al. [40] 2021 | ||
| 48, 96 h | 50, 100, 150 mg/L | Gill | − | |||||
| Freshwater | Waterborne exposure | Carassius auratus | 10, 50 mg/L | 30 d | 10, 50 mg/L | Liver | − | Qi et al. [9] 2017 |
| Megalobrama amblycephala | 5, 10, 15, 20 mg/L | 9 weeks | 20 mg/L | Liver | − | Zhang et al. [11] 2019 | ||
| Cyprinus carpio | 106 mg/L | 24 h | 106 mg/L | Blood | × | Hoseini et al. [43] 2019 | ||
| Oreochromis niloticus | 5, 10 mg/L | 70 days | 5, 10 mg/L | Liver, Muscle | + | Hegazi et al. [87] 2010 | ||
| CAT (Catalase) | ||||||||
| Sea water | Waterborne exposure | Dicentrarchus labrax | 20 mg/L | 12, 48, 84, 180 h | 20 mg/L | Blood | + | Sinha et al. [76] 2015 |
| Scophthalmus maximus | 5, 20, 40 mg/L | 24, 48, 96 h | 20, 40 mg/L | Liver | + | Jia et al. [98] 2020 | ||
| Takifugu rubripes | 5, 50, 100, 150 mg/L | 24 h | 50, 100, 150 mg/L | Gill | + | Gao et al. [40] 2021 | ||
| 48, 96 h | 50, 100, 150 mg/L | Gill | − | |||||
| Freshwater | Waterborne exposure | Carassius auratus | 10, 50 mg/L | 30 days | 10, 50 mg/L | Liver | × | Qi et al. [9] 2017 |
| Megalobrama amblycephala | 5, 10, 15, 20 mg/L | 9 weeks | 20 mg/L | Liver | − | Zhang et al. [11] 2019 | ||
| Cyprinus carpio | 106 mg/L | 24 h | 106 mg/L | Blood | − | Hoseini et al. [43] 2019 | ||
| Corbicula fluminea | 10, 25 mg/L | 24, 48 h | 10 mg/L | Digestive gland | + | Zhang et al. [84] 2020 | ||
| 10, 25 mg/L | 24, 48 h | 10 mg/L | Gill | × | ||||
| 10, 25 mg/L | 24, 48 h | 25 mg/L | Digestive gland | − | ||||
| 10, 25 mg/L | 24, 48 h | 25 mg/L | Gill | + | ||||
| GST (Glutathione-S-transferase) | ||||||||
| Sea water | Waterborne exposure | Dicentrarchus labrax | 20 mg/L | 12, 48, 84, 180 h | 20 mg/L | Blood | + | Sinha et al. [76] 2015 |
| Takifugu rubripes | 5, 50, 100, 150 mg/L | 24 h | 50, 100, 150 mg/L | Gill | + | Gao et al. [40] 2021 | ||
| 48, 96 h | 50, 100, 150 mg/L | Gill | − | |||||
| Epinephelus fuscoguttatus♀ × E. lanceolatus ♂ | 1, 2, 4, 8 mg/L | 1 week | 4, 8 mg/L | Liver, Gill | + | Kim et al. [48] 2020 | ||
| Epinephelus fuscoguttatus♀ × E. lanceolatus ♂ | 1, 2, 4, 8 mg/L | 2 weeks | 4, 8 mg/L | Liver, Gill | − | |||
| Freshwater | Waterborne exposure | Carassius auratus | 10, 50 mg/L | 30 days | 10, 50 mg/L | Liver | × | Qi et al. [9] 2017 |
| Paralichthys orbignyanus | 5, 10 mg/L | 70 days | 5, 10 mg/L | Liver, Muscle | + | Hoseini et al. [43] 2019 | ||
| Cyprinus carpio L. | 10, 20, 30 mg/L | 6, 24, 48 h | 30 mg/L | Liver | + | Li et al. [10] 2019 | ||
| 10, 20, 30 mg/L | 6, 24, 48 h | 10, 20, 30 mg/L | Gill | + | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Z.; Cao, J.; Qin, X.; Qiu, W.; Mei, J.; Xie, J. Toxic Effects on Bioaccumulation, Hematological Parameters, Oxidative Stress, Immune Responses and Tissue Structure in Fish Exposed to Ammonia Nitrogen: A Review. Animals 2021, 11, 3304. https://doi.org/10.3390/ani11113304
Xu Z, Cao J, Qin X, Qiu W, Mei J, Xie J. Toxic Effects on Bioaccumulation, Hematological Parameters, Oxidative Stress, Immune Responses and Tissue Structure in Fish Exposed to Ammonia Nitrogen: A Review. Animals. 2021; 11(11):3304. https://doi.org/10.3390/ani11113304
Chicago/Turabian StyleXu, Zhenkun, Jie Cao, Xiaoming Qin, Weiqiang Qiu, Jun Mei, and Jing Xie. 2021. "Toxic Effects on Bioaccumulation, Hematological Parameters, Oxidative Stress, Immune Responses and Tissue Structure in Fish Exposed to Ammonia Nitrogen: A Review" Animals 11, no. 11: 3304. https://doi.org/10.3390/ani11113304
APA StyleXu, Z., Cao, J., Qin, X., Qiu, W., Mei, J., & Xie, J. (2021). Toxic Effects on Bioaccumulation, Hematological Parameters, Oxidative Stress, Immune Responses and Tissue Structure in Fish Exposed to Ammonia Nitrogen: A Review. Animals, 11(11), 3304. https://doi.org/10.3390/ani11113304