
Morphological and Molecular Identification of Anisakis spp. (Nematoda: Anisakidae) in Commercial Fish from the Canary Islands Coast (Spain): Epidemiological Data

 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
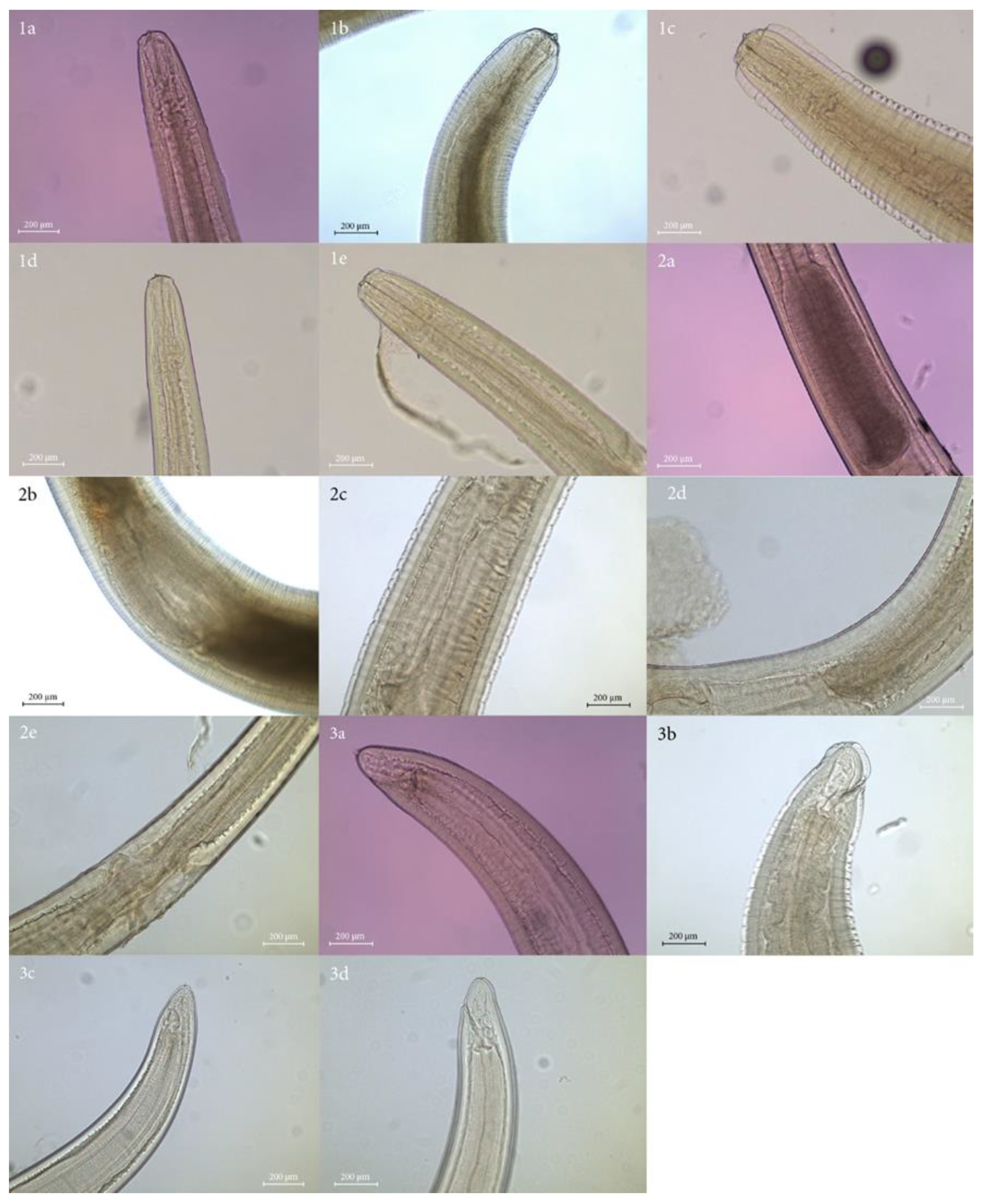
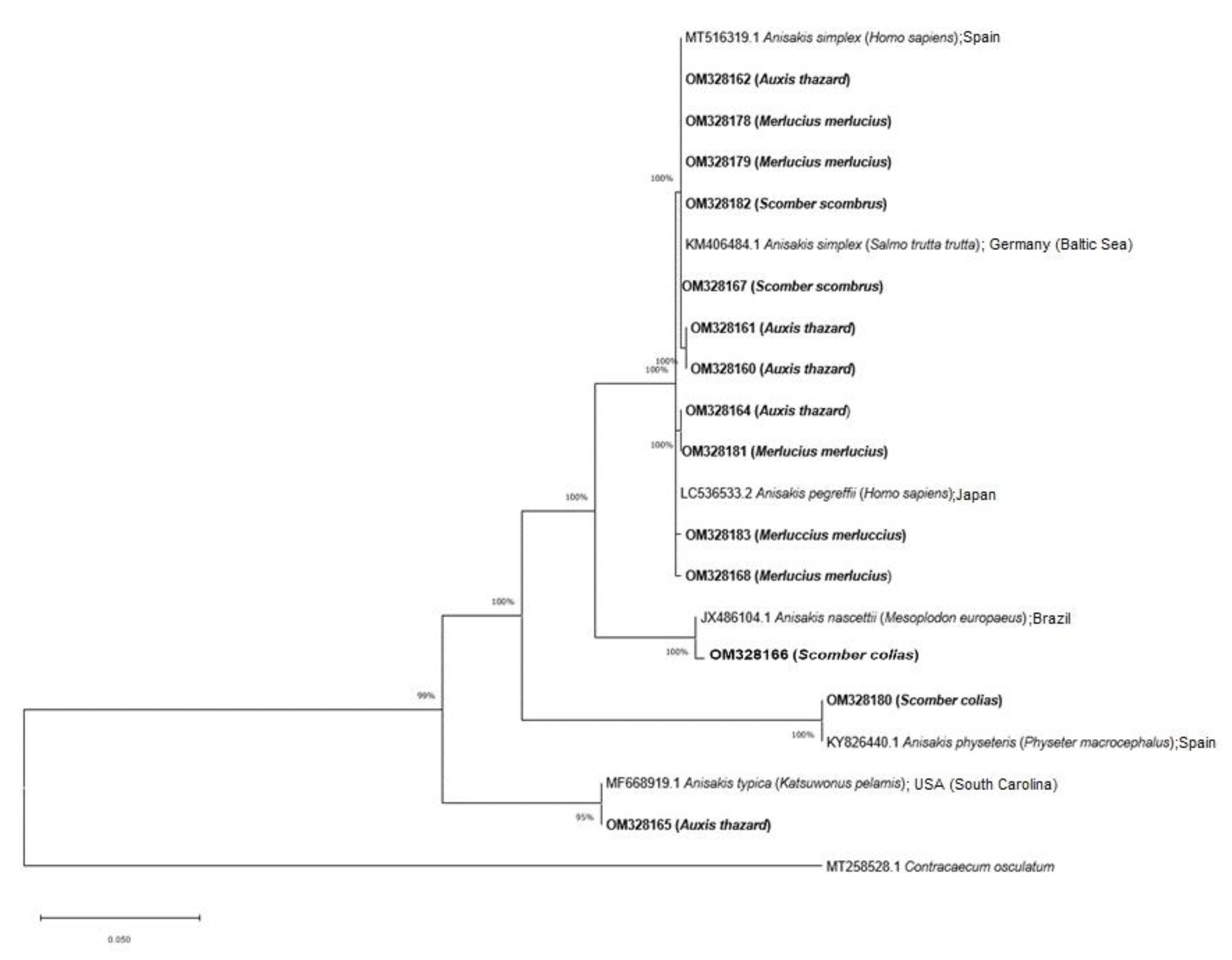
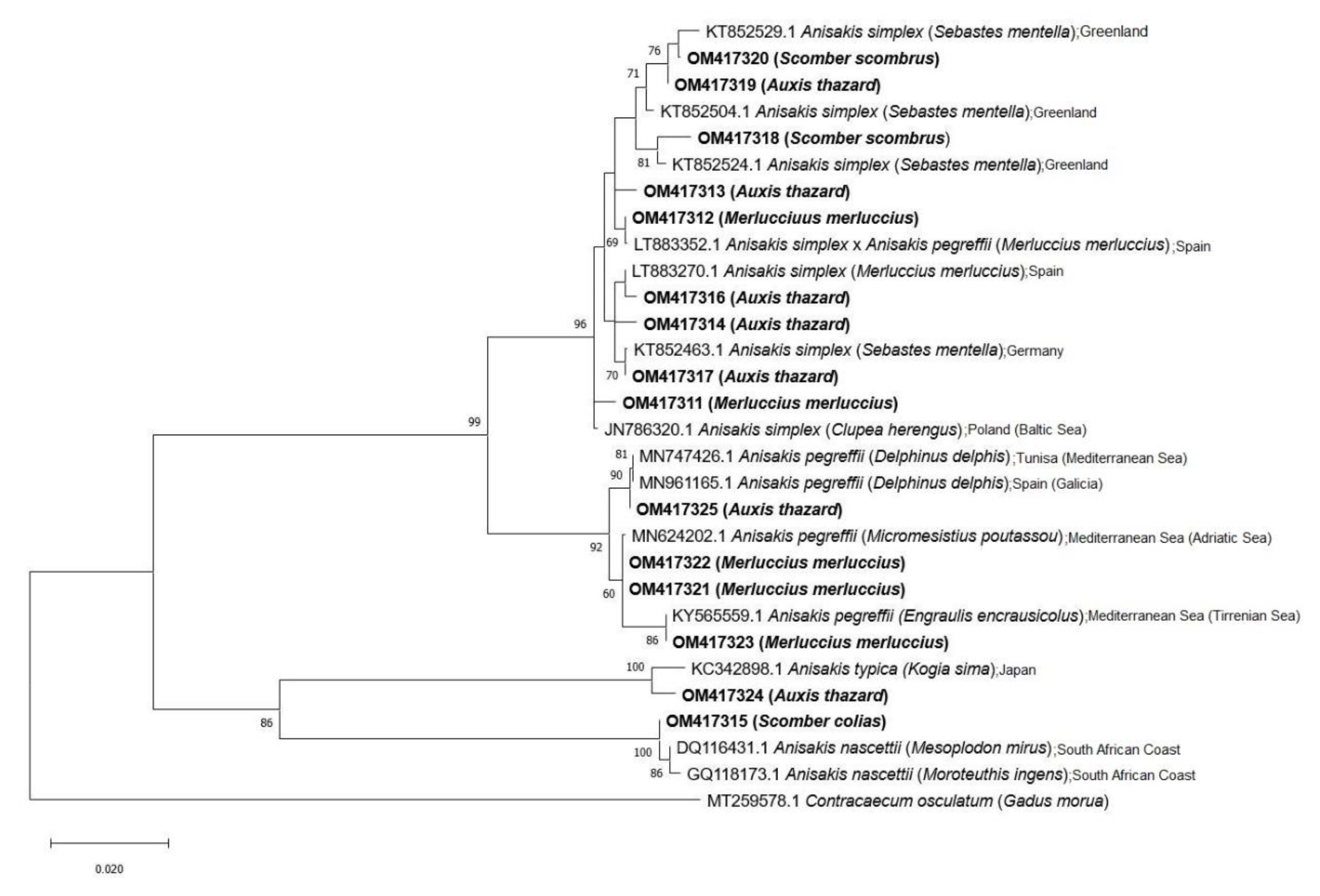
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, H.H.; Wootten, R. Anisakis and anisakiasis. Adv. Parasitol. 1978, 16, 93–163. [Google Scholar]
- Bao, M.; Pierce, G.J.; Pascual, S.; González-Muñoz, M.; Mattiucci, S.; Mladineo, I.; Cipriani, P.; Bušelić, I.; Strachan, N.J. Assessing the risk of an emerging zoonosis of worldwide concern: Anisakiasis. Sci. Rep. 2017, 7, 43699. [Google Scholar] [CrossRef]
- Lymbery, A.J.; Cheah, F.Y. Anisakid nematodes and anisakiasis. In Food-Borne Parasitic Zoonoses; Springer: Boston, MA, USA, 2007; pp. 185–207. [Google Scholar]
- Chai, J.Y.; Murrell, K.D.; Lymbery, A.J. Fish-borne parasitic zoonoses: Status and issues. Int. J. Parasitol. 2005, 35, 1233–1254. [Google Scholar] [CrossRef]
- Klimpel, S.; Palm, H.W. Anisakid nematode (Ascaridoidea) life cycles and distribution: Increasing zoonotic potential in the time of climate change? In Progress in Parasitology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 201–222. [Google Scholar]
- Mattiucci, S.; Nascetti, G. Advances and trends in the molecular systematics of anisakid nematodes, with implications for their evolutionary ecology and host parasite co-evolutionary processes. Adv. Parasitol. 2008, 66, 47–148. [Google Scholar]
- Mattiucci, S.; Paoletti, M.; Webb, S.C. Anisakis nascettii n. sp. (Nematoda: Anisakidae) from beaked whales of the southern hemisphere: Morphological description, genetic relationships between congeners and ecological data. Syst. Parasitol. 2009, 74, 199–217. [Google Scholar] [CrossRef] [PubMed]
- Cavallero, S.; Martini, A.; Migliara, G.; De Vito, C.; Iavicoli, S.; D’Amelio, S. Anisakiasis in Italy: Analysis of hospital discharge records in the years 2005–2015. PLoS ONE 2018, 13, e0208772. [Google Scholar] [CrossRef]
- Carrascosa, M.F.; Mones, J.C.; Salcines-Caviedes, J.R.; Román, J.G. A man with unsuspected marine eosinophilic gastritis. Lancet Infect. Dis. 2015, 15, 248. [Google Scholar] [CrossRef]
- Pons, F.R.; Beltran, J.G.; Gonzalez, R.A.; Gonzalez, M.A.Á.; Cusco, J.M.D.; Priego, L.B.; Urgorri, A.S. An unusual presentation of anisakiasis in the colon (with video). Gastrointest. Endos. 2015, 81, 1050–1051. [Google Scholar] [CrossRef] [PubMed]
- Van Thiel, P.H.; Kuipers, F.C.; Roskam, R.T. A nematode parasitic to herring, causing acute abdominal syndromes in man. Trop. Geogr. Med. 1960, 12, 97–113. [Google Scholar] [PubMed]
- Plath, F.; Holle, A.; Zendeh, D.; Möller, F.W.; Barten, M.; Reisinger, E.C.; Liebe, S. Anisakiasis of the stomach—A case report from Germany. Z. Gastroenterol. 2001, 39, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Mattiucci, S.; Fazii, P.; De Rosa, A.; Paoletti, M.; Megna, A.S.; Glielmo, A.; De Angelis, M.; Costa, A.; Meucci, C.; Calvaruso, V.; et al. Anisakiasis and gastroallergic reactions associated with Anisakis pegreffii infection, Italy. Emerg. Infect. Dis. 2013, 19, 496–499. [Google Scholar] [CrossRef]
- Mumoli, N.; Merlo, A. Colonic anisakiasis. CMAJ. 2013, 185, 652. [Google Scholar] [CrossRef] [PubMed]
- Mladineo, I.; Trumbić, Ž.; Radonić, I.; Vrbatović, A.; Hrabar, J.; Bušelić, I. Anisakis simplex complex: Ecological significance of recombinant genotypes in an allopatric area of the Adriatic Sea inferred by genome-derived simple sequence repeats. Int. J. Parasitol. 2017, 47, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Dupouy-Camet, J.; Touabet-Azouzi, N.; Fréalle, E.; Van Cauteren, D.; Yera, H.; Moneret-Vautrin, A. Incidence de l’anisakidose en France. Enquête rétrospective 2010–2014. Bull. Epidemiol. Hebd. 2016, 5, 64–70. [Google Scholar]
- Lucas, S.B.; Cruse, J.P.; Lewis, A.A.M. Anisakiasis in the United Kingdom. Lancet. 1985, 326, 843–844. [Google Scholar] [CrossRef]
- Cocheton, J.J.; Cabou, I.; Lecomte, I. Anisakiase et infections par les Anisakidés. Ann. Med. Interne 1991, 142, 121–130. [Google Scholar]
- Eiras, J.C.; Pavanelli, G.C.; Takemoto, R.M.; Nawa, Y. Fish-borne nematodiases in South America: Neglected emerging diseases. J. Helminthol. 2018, 92, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Hiramoto, J.T.; Tokeshi, J. Anisakiasis in Hawaii: A radiological diagnosis. Hawaii J. Med. Public. Health. 1991, 50, 202–203. [Google Scholar]
- Couture, C.; Gagnon, J.; Desbiens, C. Human intestinal anisakiosis due to consumption of raw salmon. Am. J. Surg. Pathol. 2003, 27, 1167–1172. [Google Scholar] [CrossRef]
- Audicana, M.T.; Kennedy, M.W. Anisakis simplex: From obscure infectious worm to inducer of immune hypersensitivity. Clin. Microbiol. Rev. 2008, 21, 360–379. [Google Scholar] [CrossRef]
- Li, S.W.; Shiao, S.H.; Weng, S.C.; Liu, T.H.; Su, K.E.; Chen, C.C. A case of human infection with Anisakis simplex in Taiwan. Gastrointest. Endosc. 2015, 82, 757–758. [Google Scholar] [CrossRef] [PubMed]
- Sohn, W.M.; Na, B.K.; Kim, T.H.; Park, T.J. Anisakiasis: Report of 15 gastric cases caused by Anisakis type I larvae and a brief review of Korean anisakiasis cases. Korean. J. Parasitol. 2015, 53, 465. [Google Scholar] [CrossRef] [PubMed]
- Paltridge, G.P.; Faoagali, J.L.; Angus, H.B. Intestinal anisakiasis: A new New Zealand disease. N. Z. Med. J. 1984, 97, 558–559. [Google Scholar] [PubMed]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific opinion on risk assessment of parasites in fishery products. EFSA 2010, 8, 1543. [Google Scholar]
- Leuckart, R. Die menschlichen Parasiten und die von ihren herriihrend Krankheiten. Leipzig 1876, 2 Pt 3, 513–882. [Google Scholar]
- Van Thiel, P.H. Anisakiasis. Parasitology 1962, 52, 3–4. [Google Scholar]
- Villazanakretzer, D.L.; Napolitano, P.G.; Cumming, K.F.; Magann, E.F. Fish parasites a growing concern during pregnancy. Obstet. Gynecol. Surv. 2016, 71, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Moore David, A.J.; Girdwood, R.W.A.; Chiodini Peter, L. Treatment of anisakiasis with albendazole. Lancet 2002, 360, 54. [Google Scholar] [CrossRef]
- Dziekonska-Rynko, J.; Rokicki, J.; Jablonowski, Z. Effects of ivermectin and albendazole against Anisakis simplex in vitro and in guinea pigs. J. Parasitol. 2002, 88, 395–398. [Google Scholar] [CrossRef]
- Arias Diaz, J.; Zuloaga, J.; Vara, E.; Balibrea, J.; Balibrea, J.L. Efficacy of albendazole against Anisakis simplex larvae in vitro. Dig. Liver Dis. 2006, 38, 24–26. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, J.; Moore, T.A. Emerging helminth zoonoses. Int. J. Parasitol. 2000, 30, 1351–1359. [Google Scholar] [CrossRef]
- Reglamento (CE) Nº 854/2004 del Parlamento Europeo y del Consejo, de 29 de Abril de 2004. Available online: https://www.boe.es/buscar/doc.php?id=DOUE-L-2004-81037 (accessed on 21 December 2021).
- Berland, B. Nematodes from some Norwegian marine fishes. Sarsia 1961, 2, 1–50. [Google Scholar] [CrossRef]
- Petter, A.J.; Maillard, C. Larves d’ascarides parasites de poissons en Méditerranée occidentale. Bull. Mus. Natl. Hist. Nat. 1988, 10, 347–369. [Google Scholar]
- Shamsi, S.; Gasser, R.; Beveridge, I. Description and genetic characterisation of Hysterothylacium (Nematoda: Raphidascarididae) larvae parasitic in Australian marine fishes. Parasitol. Int. 2013, 62, 320–328. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Tibshirani, R.J.; Efron, B. An introduction to the bootstrap. Monogr. Stat. Appl. Probab. 1993, 57, 1–436. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; URL. [Google Scholar]
- López, C.; Clemente, S.; Almeida, C.; Brito, A.; Hernández, M. A genetic approach to the origin of Millepora sp. in the eastern Atlantic. Coral Reefs. 2015, 34, 631–638. [Google Scholar] [CrossRef]
- Zhu, X.; Gasser, R.B.; Podolska, M.; Chilton, N.B. Characterisation of anisakid nematodes with zoonotic potential by nuclear ribosomal DNA sequences. Int. J. Parasitol. 1998, 28, 1911–1921. [Google Scholar] [CrossRef]
- Nadler, S.A.; Hudspeth, D.S.S. Phylogeny of the Ascaridoidea (Nematoda: Ascaridida) based on three genes and morphology: Hypotheses of structural and sequence evolution. J. Parasitol. 2000, 86, 380–393. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed]
- Bas, C.; Castro, J.J.; Hernández-García, V.; Lorenzo, J.M.; Moreno, T.; Pajuelo, J.G.; Ramos, A.J. La pesca en Canarias y áreas de influencia; Cabildo Insular de Gran Canaria: Las Palmas de Gran Canaria, España, 1995; pp. 1–331.
- Aristegui, J.; Barton, E.D.; Álvarez-Salgado, X.A.; Santos, A.M.P.; Figueiras, F.G. Subregion al ecosystem variability in the Canary Current upwelling. Prog. Oceanogr. 2009, 83, 33–48. [Google Scholar] [CrossRef]
- Aristegui, J.; Sangrá, P.; Hernández-León, S.; Cantón, M.; Hernández-Guerra, A.; Kerling, J.L. Island-induced eddies in the Canary Islands. Depp-Sea Res. I Oceanog. Res. Pap. 1994, 41, 1509–1525. [Google Scholar] [CrossRef]
- Sangrà, P.; Pascual, A.; Rodríguez-Santana, Á.; Machín, F.; Mason, E.; McWilliams, J.C.; Auladell, M. The Canary Eddy Corridor: A major pathway for long-lived eddies in the subtropical North Atlantic. Depp-Sea Res. I Oceanog. Res. Pap. 2009, 56, 2100–2114. [Google Scholar] [CrossRef]
- Lobel, P.S.; Robinson, A.R. Transport and entrapment of fish larvae by ocean mesoscale eddies and currents in Hawaiian waters. Deep Sea Res. 1986, 33, 483–500. [Google Scholar] [CrossRef]
- Lobel, P.S.; Robinson, A.R. Larval fishes and zooplankton in cyclonic eddy in Hawaiian waters. J. Plankton Res. 1988, 10, 1209–1223. [Google Scholar] [CrossRef]
- Crawford, R.; Jorgenson, J. Density distribution of fish in the presence of whales at the Admiralty Inlet land fast ice edge. Arctic 1990, 43, 215–222. [Google Scholar] [CrossRef][Green Version]
- Gómez, M. Biomasa y Actividad Metabólica del Zooplancton en Relación con un Efecto de Masa de isla en Aguas de Gran Canaria; Universidad de Las Palmas de Gran Canaria: Las Palmas de Gran Canaria, Spain, 1991. [Google Scholar]
- Rodríguez, J.M.; Barton, E.D.; Hernández-León, S.; Arístegui, J. The influence of mesoscale physical processes on the larval fish community in the Canaries CTZ, in summer. Prog. Oceanogr. 2004, 62, 171–188. [Google Scholar] [CrossRef]
- Moro, L.; Martín, J.L.; Garrido, M.J.; Izquiero, I. (Eds.) Lista de Especies Marinas de Canarias (Algas, Hongos, Plantas y Animales); Consejería de Política Territorial y Medio Ambiente del Gobierno de Canarias: La Laguna, Islas Canarias, 2003; pp. 1–248.
- Nagasawa, K.; Moravec, F. Larval anisakid nematodes of Japanese common squid (Todarodes pacificus) from the Sea of Japan. J. Parasitol. 1995, 81, 69–75. [Google Scholar] [CrossRef]
- Abollo, E.; Gestal, C.; Pascual, S. Anisakis infestation in marine fish and cephalopods from Galician waters: An updated perspective. Parasitol. Res. 2001, 87, 492–499. [Google Scholar] [PubMed]
- Vidaček, S.; de las Heras, C.; Solas, M.T.; Mendizábal, A.; Rodriguez-Mahillo, A.I.; González-Muñoz, M.; Tejada, M. Anisakis simplex allergens remain active after conventional or microwave heating and pepsin treatments of chilled and frozen L3 larvae. J. Sci. Food Agric. 2009, 89, 997–2002. [Google Scholar] [CrossRef]
- Cipriani, P.; Sbaraglia, G.L.; Palomba, M.; Giulietti, L.; Bellisario, B.; Bušelić, I.; Mladineo, I.; Cheleschi, R.; Nascetti, G.; Mattiucci, S. Anisakis pegreffii (Nematoda: Anisakidae) in European anchovy Engraulis encrasicolus from the Mediterranean Sea: Fishing ground as a predictor of parasite distribution. Fish. Res. 2018, 202, 59–68. [Google Scholar] [CrossRef]
- Gutiérrez-Galindo, J.F.; Osanz-Mur, A.C.; Mora-Ventura, M.T. Occurrence and infection dynamics of anisakid larvae in Scomber scombrus, Trachurus trachurus, Sardina pilchardus, and Engraulis encrasicolus from Tarragona (NE Spain). Food Control 2010, 21, 1550–1555. [Google Scholar] [CrossRef]
- Mattiucci, S.; Cipriani, P.; Levsen, A.; Paoletti, M.; Nascetti, G. Molecular epidemiology of Anisakis and Anisakiasis: An ecological and evolutionary road map. Adv. Parasitol. 2018, 99, 93–263. [Google Scholar] [PubMed]
- Rello, F.J.; Adroher, F.J.; Benítez, R.; Valero, A. The fishing area as a possible indicator of the infection by anisakids in anchovies (Engraulis encrasicolus) from southwestern Europe. Int. J. Food Microbiol. 2009, 129, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Klapper, R.; Kuhn, T.; Münster, J.; Levsen, A.; Karl, H.; Klimpel, S. Anisakid nematodes in beaked redfish (Sebastes mentella) from three fishing grounds in the North Atlantic, with special notes on distribution in the fish musculature. Vet. Parasitol. 2015, 207, 72–80. [Google Scholar] [CrossRef]
- Rahmati, A.R.; Kiani, B.; Afshari, A.M.; Moghaddas, E.; Williams, M.; Shamsi, S. World-wide prevalence of Anisakis larvae in fish and its relationship to human allergic anisakiasis: A systematic review. Parasitol. Res. 2020, 119, 3585–3594. [Google Scholar] [CrossRef] [PubMed]
- FAO. Organización de las Naciones Unidas para la Alimentación Agricultura y la Agricultura Alimentación, Fishery and Aquaculture Statistics—Estadísticas de Pesca y Acuicultura 2021. Available online: https://www.fao.org/statistics/es/ (accessed on 11 February 2022).
- D’Amelio, S.; Mathiopoulos, K.D.; Brandonisio, O.; Lucarelli, G.; Doronzo, F.; Paggi, L. Diagnosis of a case of gastric anisakidosis by PCR-based restriction fragment length polymorphism analysis. Parassitologia 1999, 41, 591–593. [Google Scholar]
- Audicana, M.T.; Ansotegui, I.J.; Fernandez de Corres, L.; Kennedy, M.W. Anisakis simplex: Dangerous—Dead and alive? Trends Parasitol. 2002, 18, 20–25. [Google Scholar] [CrossRef]
- Del Pozo, M.D.; Audicana, M.; Diez, J.M.; Muñoz, D.; Ansotegui, I.J.; Fernández, E.; García, M.; Etxenagusia, M.; Moneo, I.; Fernández de Corres, L. Anisakis simplex, a relevant etiologic factor in acute urticaria. Allergy 1997, 52, 576–579. [Google Scholar] [CrossRef]
- Suzuki, J.; Murata, R.; Hosaka, M.; Araki, J. Risk factors for human Anisakis infection and association between the geographic origins of Scomber japonicus and anisakid nematodes. Int. J. Food Microbiol. 2010, 137, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Arizono, N.; Yamada, M.; Tegoshi, T.; Yoshikawa, M. Anisakis simplex sensu stricto and Anisakis pegreffii: Biologicalcharacteristics and pathogenetic potential in human anisakiasis. Foodborne Pathog. Dis. 2012, 9, 517–521. [Google Scholar] [CrossRef]
- Jeon, C.H.; Kim, J.H. Pathogenic potential of two sibling species, Anisakis simplex (ss) and Anisakis pegreffii (Nematoda: Anisakidae): In Vitro and in vivo studies. BioMed Res. Int. 2015, 2015, 983656. [Google Scholar] [CrossRef] [PubMed]
- Umehara, A.; Kawakami, Y.; Araki, J.; Uchida, A. Molecular identification of the etiological agent of the human anisakiasis in Japan. Parasitol. Int. 2007, 56, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Mattiucci, S.; Abaunza, P.; Damiano, S.; Garcia, A.; Santos, M.N.; Nascetti, G. Distribution of Anisakis larvae, identified by genetic markers, and their use for stock characterization of demersal and pelagic fish from European waters: An update. J. Helminthol. 2007, 81, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Mattiucci, S.; Paoletti, M.; Borrini, F.; Palumbo, M.; Palmieri, R.M.; Gomes, V.; Nascetti, G. First molecular identification of the zoonotic parasite Anisakis pegreffii (Nematoda: Anisakidae) in a paraffin-embedded granuloma taken from a case of human intestinal anisakiasis in Italy. BMC. Infect. Dis. 2011, 11, 82. [Google Scholar] [CrossRef] [PubMed]
- Fumarola, L.; Monno, R.; Ierardi, E.; Rizzo, G.; Giannelli, G.; Lalle, M.; Pozio, E. Anisakis pegreffi etiological agent of gastric infections in two Italian women. Foodborne Pathog. Dis. 2009, 6, 1157–1159. [Google Scholar] [CrossRef]
- Palma, R.; Mattiucci, S.; Panetta, C.; Raniolo, M.; Magliocca, F.M.; Pontone, S. Paucisymptomatic gastric anisakiasis: Endoscopical removal of Anisakis sp. larva. Minim. Invasive Surg. 2018, 2, 1. [Google Scholar] [CrossRef][Green Version]
- Costa, G.; Cavallero, S.; D’Amelio, S.; Paggi, L.; Santamaria, M.; Perera, C.; Khadem, M. Helminth parasites of the Atlantic chub mackerel, Scomber colias Gmelin, 1789 from Canary Islands, Central North Atlantic, with comments on their relations with other Atlantic regions. Acta Parasitol. 2011, 56, 98–104. [Google Scholar] [CrossRef]
- Dupouy-Camet, J. Helminth Infections and Their Impact on Global Public Health; Bruschi, F., Ed.; Springer: Vienna, Austria, 2014; pp. 229–279. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Host Species (Common Name) | Prevalence of Anisakis Larvae P (%) (+/n) (CI 95%) | Ncoll | Mean Intensity (CI 95%) | Mean Abundance (CI 95%) | Mean T.W. ± S.D (CI 95%) | Mean T.L. ± S.D (CI 95%) |
|---|---|---|---|---|---|---|
| Auxis thazard (frigate mackerel) | 21.43% (3/14) (4.66%, 50.8%) | 14 | 4.67 (1.00, 7.67) | 1.00 (0.07, 3.62) | 340.18 ± 29.97 (322.88, 357.49) | 29.24 ± 0.87 (26.14, 28.97) |
| Diplodus sargus sargus (white seabream) | 0% (0/2) - | 0 | - | - | 257.745 ± 55.22 - | 25.50 ± 2.12 - |
| Merluccius merluccius (European hake) | 85.18% (23/27) (66.27%, 95.81%) | 205 | 8.91 (5.44, 16.76) | 7.59 (4.44, 14.45) | 271.69 ± 57.34 (249.01, 294.38) | 27.56 ± 3.58 (26.24, 28.86) |
| Mullus surmuletus (red mullet) | 0% (0/6) - | 0 | - | - | 219.07 ± 61.92 (190.74, 295.3) | 23.30 ± 0.84 (22.4, 23.8) |
| Pagellus erythinus (common Pandora) | 0% (0/11) - | 0 | - | - | 222.06 ± 69.04 (190.03, 270.69) | 25.59 ± 2.55 (24.41, 27.27) |
| Sardinella aurita (sardinelle) | 0% (0/21) - | 0 | - | - | 117.78 ± 22.00 (108.66, 126.89) | 21.16 ± 2.37 (20.08, 22.24) |
| Sarpa salpa (salema) | 0% (0/5) - | 0 | - | - | 269.90 ± 26.83 (251.57, 295.93) | 26.90 ± 0.74 (26.3, 27.4) |
| Scomber colias (Atlantic chub mackerel) | 26.6% (8/30) (12.28%, 45.89%) | 10 | 1.25 (1.00, 1.50) | 0.33 (0.13, 0.57) | 237.73 ± 88.67 (210.05, 274.06) | 29.99 ± 3.58 (28.65, 31.32) |
| Scomber scombrus (mackerel) | 62.5% (10/16) (35.43%, 84.8%) | 266 | 26.60 (8.80, 56.65) | 16.63 (5.12, 39.25) | 330.96 ± 51.37 (308.65, 357.14) | 29.95 ± 2.81 (28.93, 31.84) |
| Serranus atricauda (black tail comber) | 0% (0/6) | 0 | - | - | 138.36 ± 19.27 (124.37, 152.25) | 22.08 ± 1.02 (21.33, 22.75) |
| Trachurus picturatus (blue jack mackerel) | 0% (0/34) | 0 | - | - | 52.90 ± 13.10 (49.19, 57.87) | 18.69 ± 1.36 (18.22, 19.17) |
| Host Species | Molecularly Identified Species of Anisakis |
|---|---|
| Auxis thazard | Anisakis simplex (s.s.) Anisakis pegreffii Anisakis typica Anisakis simplex × Anisakis pegreffii |
| Merluccius merluccius | Anisakis simplex (s.s.) Anisakis pegreffii Anisakis simplex × Anisakis pegreffii |
| Scomber colias | Anisakis nascettii Anisakis physeteris |
| Scomber scombrus | Anisakis simplex (s.s.) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin-Carrillo, N.; García-Livia, K.; Baz-González, E.; Abreu-Acosta, N.; Dorta-Guerra, R.; Valladares, B.; Foronda, P. Morphological and Molecular Identification of Anisakis spp. (Nematoda: Anisakidae) in Commercial Fish from the Canary Islands Coast (Spain): Epidemiological Data. Animals 2022, 12, 2634. https://doi.org/10.3390/ani12192634
Martin-Carrillo N, García-Livia K, Baz-González E, Abreu-Acosta N, Dorta-Guerra R, Valladares B, Foronda P. Morphological and Molecular Identification of Anisakis spp. (Nematoda: Anisakidae) in Commercial Fish from the Canary Islands Coast (Spain): Epidemiological Data. Animals. 2022; 12(19):2634. https://doi.org/10.3390/ani12192634
Chicago/Turabian StyleMartin-Carrillo, Natalia, Katherine García-Livia, Edgar Baz-González, Néstor Abreu-Acosta, Roberto Dorta-Guerra, Basilio Valladares, and Pilar Foronda. 2022. "Morphological and Molecular Identification of Anisakis spp. (Nematoda: Anisakidae) in Commercial Fish from the Canary Islands Coast (Spain): Epidemiological Data" Animals 12, no. 19: 2634. https://doi.org/10.3390/ani12192634
APA StyleMartin-Carrillo, N., García-Livia, K., Baz-González, E., Abreu-Acosta, N., Dorta-Guerra, R., Valladares, B., & Foronda, P. (2022). Morphological and Molecular Identification of Anisakis spp. (Nematoda: Anisakidae) in Commercial Fish from the Canary Islands Coast (Spain): Epidemiological Data. Animals, 12(19), 2634. https://doi.org/10.3390/ani12192634