Possible Effects of Early Maternal Separation on the Gut Microbiota of Captive Adult Giant Pandas



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Feces Samples Collection
2.2. Feeding and Management
2.3. Total DNA Extraction and Library Construction
2.4. Metagenomic Shotgun Sequencing of Microbialcommunities
2.5. LC-MS/MS Analysis
2.6. Correlation Analysis
3. Results
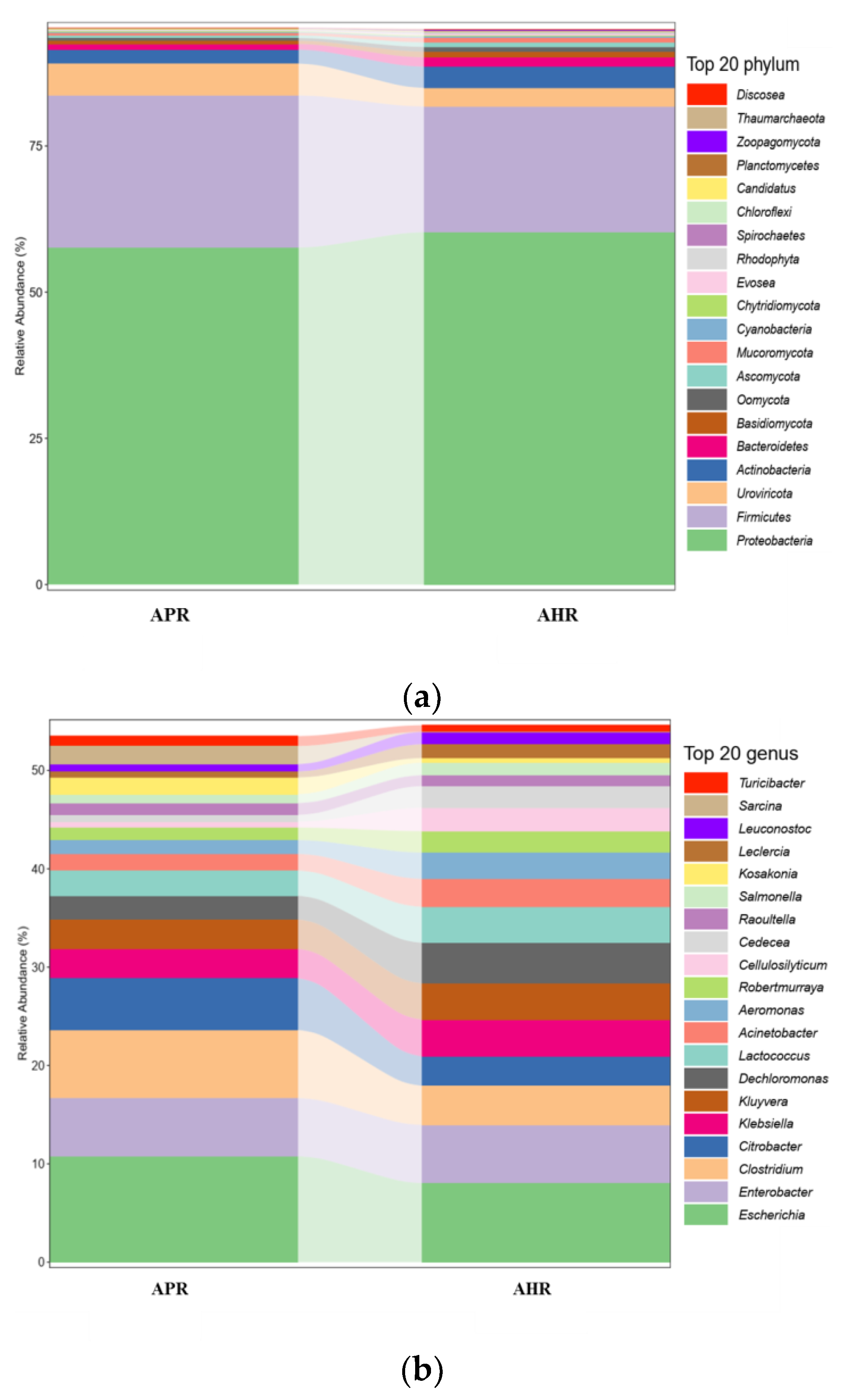
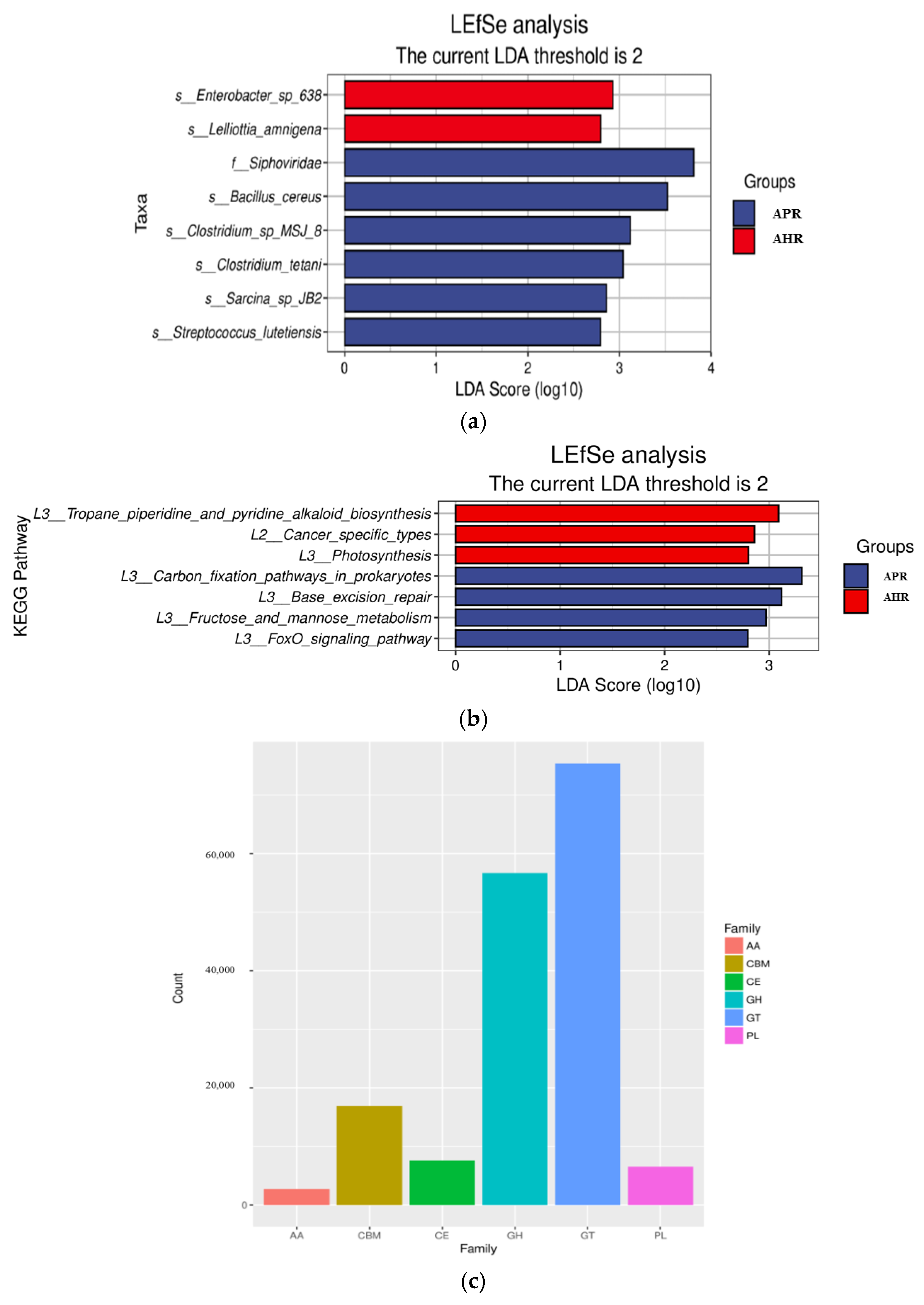
3.1. Effects of Early Nursery Environment on Gut Microbial Composition of Captive Giant Pandas
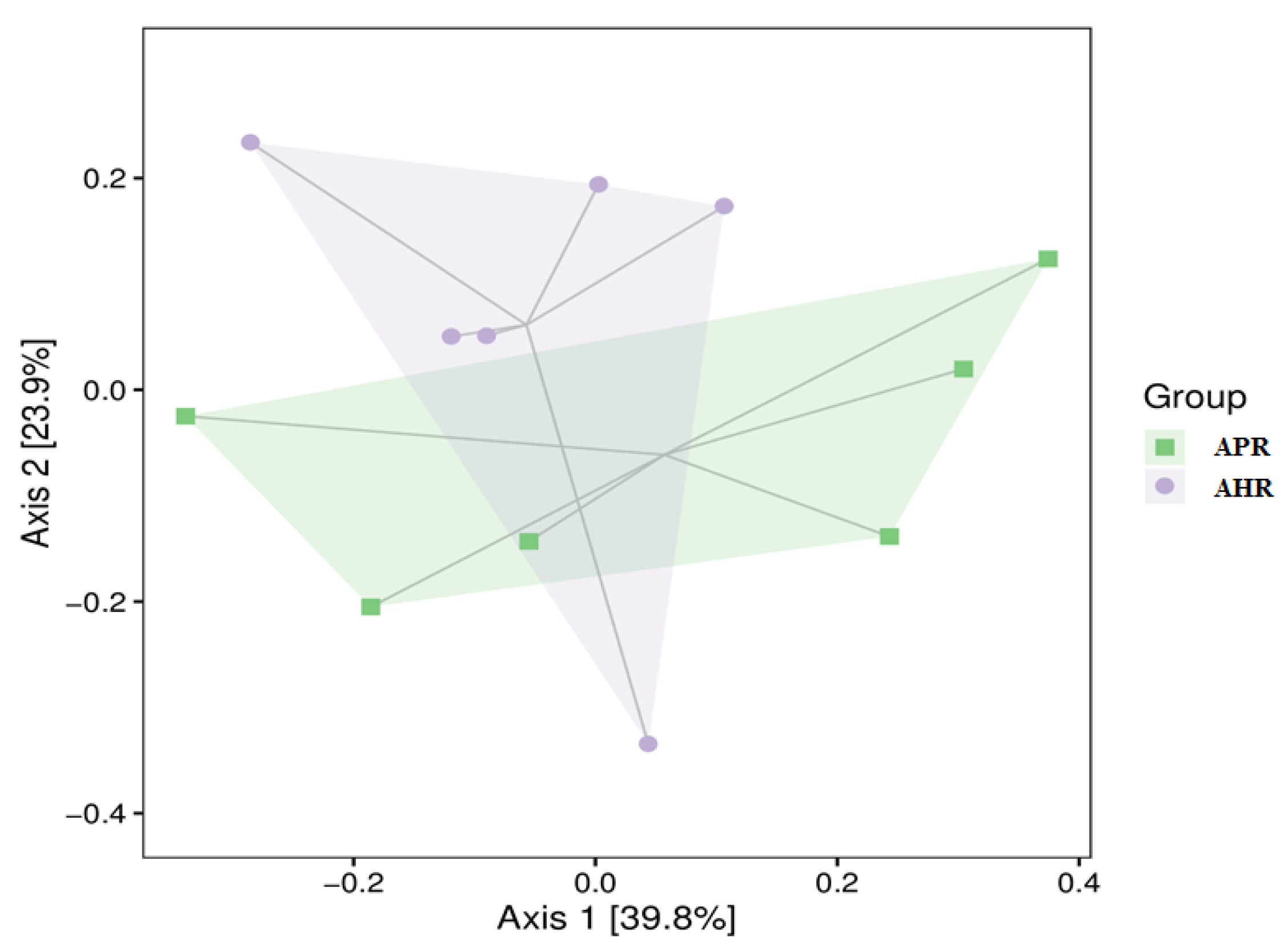
3.2. Effects of Early Nursery Environment on Gut Microbial Diversity in Captive Giant Pandas
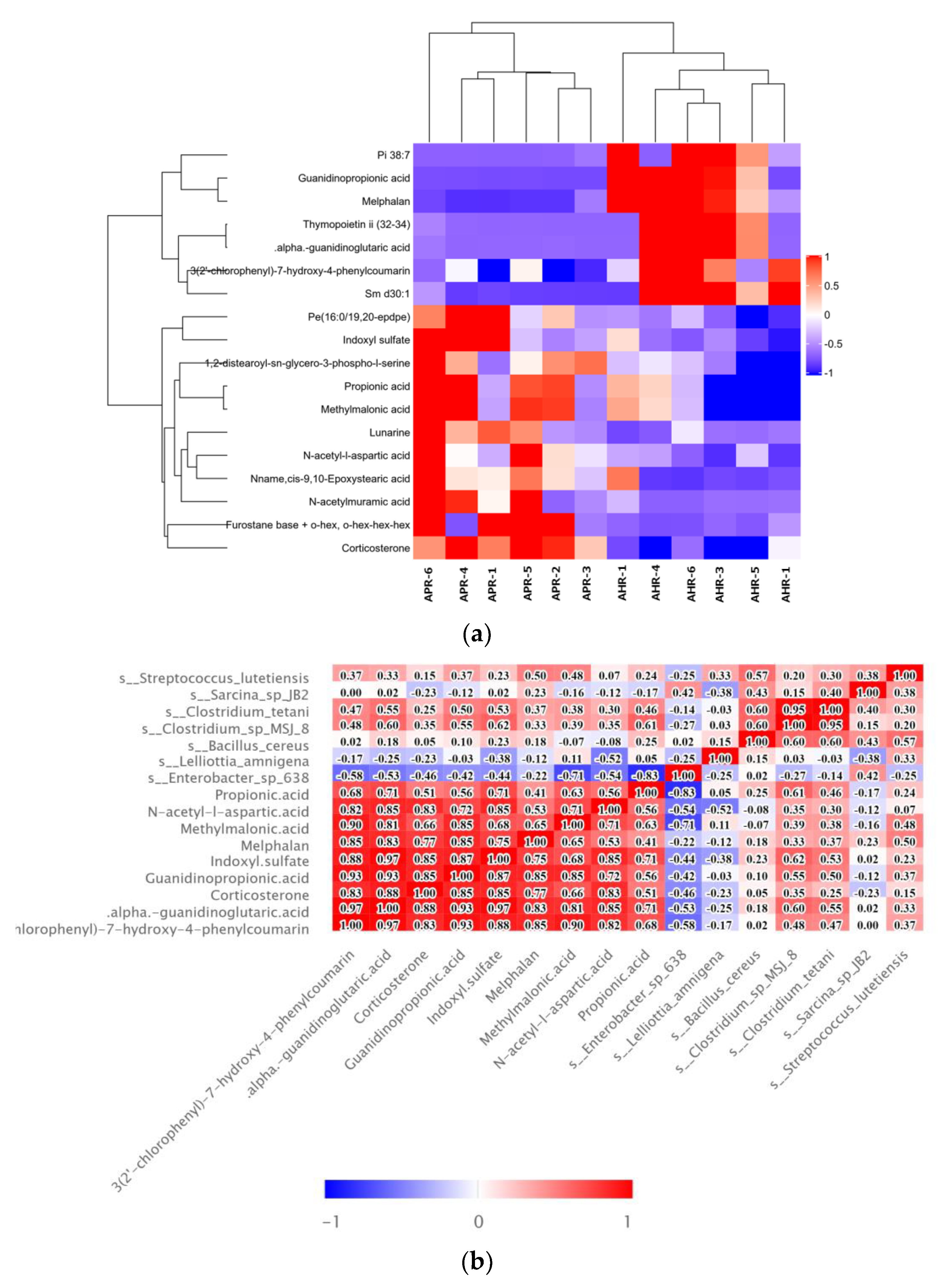
3.3. Effects of Early Nursery Environment on Fecal Metabolites in Adult Captive Giant Pandas
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sánchez, M.M.; Ladd, C.O.; Plotsky, P.M. Early adverse experience as a developmental risk factor for later psychopathology: Evidence from rodent and primate models. Dev. Psychopathol. 2001, 13, 419–449. [Google Scholar] [CrossRef]
- Kendler, K.S.; Gardner, C.O. Depressive vulnerability, stressful life events and episode onset of major depression: A longitudinal model. Psychol. Med. 2016, 46, 1865–1874. [Google Scholar] [CrossRef] [PubMed]
- Partrick, K.A.; Chassaing, B.; Beach, L.Q.; McCann, K.E.; Gewirtz, A.T.; Huhman, K.L. Acute and repeated exposure to social stress reduces gut microbiota diversity in Syrian hamsters. Behav. Brain Res. 2018, 345, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.A.; Rinaman, L.; Cryan, J.F. Stress & the gut-brain axis: Regulation by the microbiome. Neurobiol. Stress 2017, 7, 124–136. [Google Scholar] [PubMed]
- Marin, I.A.; Goertz, J.E.; Ren, T.; Rich, S.S.; Onengut-Gumuscu, S.; Farber, E.; Wu, M.; Overall, C.C.; Kipnis, J.; Gaultier, A. Microbiota alteration is associated with the development of stress-induced despair behavior. Sci. Rep. 2017, 7, 43859. [Google Scholar] [CrossRef]
- Swaisgood, R.R.; Zhou, X.P.; Zhang, G.Q.; Lindburg, D.G.; Zhang, H.M. Application of Behavioral Knowledge to Conservation in the Giant Panda. Int. J. Comp. Psychol. 2003, 16, 65–84. [Google Scholar]
- Zhang, M.Y.; Zhang, X.H.; James, A.; Hou, R. Effects of Different Nursing Methods on the Behavioral Response of Adult Captive Giant Pandas (Ailuropoda melanoleuca). Animals 2021, 11, 626. [Google Scholar] [CrossRef]
- Snyder, R.J.; Zhang, A.J.; Zhang, Z.H.; Li, G.H.; Tian, Y.Z.; Huang, X.M.; Luo, L.; Bloomsmith, M.A.; Forthman, D.L.; Maple, T.L. Behavioral and developmental consequences of early rearing experience for captive giant pandas (Ailuropoda melanoleuca). J Comp. Psychol. 2003, 117, 235–245. [Google Scholar] [CrossRef]
- Wu, W.L.; Adame, M.D.; Liou, C.W.; Barlow, J.T.; Lai, T.T.; Sharon, G.; Schretter, C.E.; Needham, B.D.; Wang, M.I.; Tang, W.; et al. Microbiota regulate social behaviour via stress response neurons in the brain. Nature 2021, 595, 409–414. [Google Scholar] [CrossRef]
- Chevalier, G.; Siopi, E.; Guenin-Macé, L.; Pascal, M.; Laval, T.; Rifflet, A.; Boneca, I.G.; Demangel, C.; Colsch, B.; Pruvost, A.; et al. Effect of gut microbiota on depressive-like behaviors in mice is mediated by the endocannabinoid system. Nat. Commun. 2020, 11, 6363. [Google Scholar] [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.H.; Wei, F.W. Giant Panda Ex-Situ Conservation: Theory and Practice; Science Press: Beijing, China, 2006. [Google Scholar]
- Meyer, M.; Kircher, M. Illumina sequencing library preparation for highly multiplexed target capture and sequencing. Cold Spring Harb. Protoc. 2010, 6, 5448. [Google Scholar] [CrossRef] [PubMed]
- Truong, D.T.; Franzosa, E.A.; Tickle, T.L.; Scholz, M.; Weingart, G.; Pasolli, E.; Tett, A.; Huttenhower, C.; Segata, N. MetaPhlAn2 for enhanced metagenomic taxonomic profiling. Nat. Methods 2015, 12, 902–903. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Uchiyama, I. MBGD: A platform for microbial comparative genomics based on the automated construction of orthologous groups. Nucleic Acids Res. 2007, 35, 343–346. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Wang, X.Y.; Zhang, X.H.; Hou, R. Urine metabolomics reveals the effects of confined environment on mating choice in adult male giant pandas. Physiol. Behav. 2022, 249, 113744. [Google Scholar] [CrossRef]
- De Palma, G.; Blennerhassett, P.; Lu, J.; Deng, Y.; Park, A.J.; Green, W.; Denou, E.; Silva, M.A.; Santacruz, A.; Sanz, Y.; et al. Microbiota and host determinants of behavioural phenotype in maternally separated mice. Nat. Commun. 2015, 6, 7735. [Google Scholar] [CrossRef]
- Ning, Y.; Qi, J.Z.; Dobbins, M.T.; Liang, X.; Wang, J.X.; Chen, S.Y.; Ma, J.Z.; Jiang, G.S. Comparative Analysis of Microbial Community Structure and Function in the Gut of Wild and Captive Amur Tiger. Front. Microbiol. 2020, 11, 1665. [Google Scholar] [CrossRef]
- Jiang, F.; Song, P.F.; Wang, H.J.; Zhang, J.J.; Liu, D.X.; Cai, Z.Y.; Gao, H.M.; Chi, X.W.; Zhang, T.Z. Comparative analysis of gut microbial composition and potential functions in captive forest and alpine musk deer. Appl. Microbiol. Biotechnol. 2022, 106, 1325–1339. [Google Scholar] [CrossRef]
- Zhu, L.F.; Wu, Q.; Dai, J.Y.; Zhang, S.N.; Wei, F.W. Evidence of cellulose metabolism by the giant panda gut microbiome. Proc. Natl. Acad. Sci. USA 2011, 108, 17714–17719. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Fang, Z.M.; Zhou, P.; Chang, F.; Hong, Y.Z.; Zhang, X.C.; Peng, H.; Xiao, Y.Z. Evidence for lignin oxidation by the giant panda fecal microbiome. PLoS ONE 2012, 7, e50312. [Google Scholar]
- Zhang, J.; Guo, Z.; Xue, Z.S.; Sun, Z.H.; Zhang, M.H.; Wang, L.F.; Wang, G.Y.; Wang, F.; Xu, J.; Cao, H.F.; et al. A phylo-functional core of gut microbiota in healthy young Chinese cohorts across lifestyles, geography and ethnicities. ISME J. 2015, 9, 1979–1990. [Google Scholar] [CrossRef] [PubMed]
- Montagne, L.; Pluske, J.R.; Hampson, D.J. A review of interactions between dietary fibre and the intestinal mucosa, and their consequences on digestive health in young non-ruminant animals. Anim. Feed Sci. Tech. 2003, 108, 95–117. [Google Scholar] [CrossRef]
- Bailey, M.T.; Dowd, S.E.; Galley, J.D.; Hufnagle, A.R.; Allen, R.G.; Lyte, M. Exposure to a social stressor alters the structure of the intestinal microbiota: Implications for stressor-induced immunomodulation. Brain Behav. Immun. 2011, 25, 397–407. [Google Scholar] [CrossRef]
- Inglese, M.; Rusinek, H.; George, I.C.; Babb, J.S.; Grossman, R.I.; Gonen, O. Global average gray and white matter N-acetylaspartate concentration in the human brain. Neuroimage 2008, 41, 270–276. [Google Scholar] [CrossRef]
- Wingfield, J.C. Control of behavioural strategies for capricious environments. Anim. Behav. 2003, 66, 807–815. [Google Scholar] [CrossRef]
- Wingfield, J.C.; Kitaysky, A.S. Endocrine responses to unpredictable environmental events: Stress or anti-stress hormones? Integr. Comp. Biol. 2002, 42, 600–609. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Name | Gender | Age | Single or Twins |
|---|---|---|---|---|
| Adult Hand-Raised (AHR) | Gong Zai | Male | 13 years | single |
| Adult Hand-Raised (AHR) | Ying Ying | Male | 13 years | single |
| Adult Hand-Raised (AHR) | Cheng Shuang | Male | 9 years | twins |
| Adult Hand-Raised (AHR) | Ni Da | Female | 6 years | twins |
| Adult Hand-Raised (AHR) | Cheng Da | Female | 10 years | single |
| Adult Hand-Raised (AHR) | Mei Lun | Female | 8 years | twins |
| Adult Parent-Raised (APR) | Mei Lan | Female | 15 years | single |
| Adult Parent-Raised (APR) | Lou ABao | Male | 14 years | single |
| Adult Parent-Raised (APR) | Mei ABao | Female | 11 years | single |
| Adult Parent-Raised (APR) | ZhiZhi | Female | 12 years | single |
| Adult Parent-Raised (APR) | YaZai | Female | 15 years | single |
| Adult Parent-Raised (APR) | Zhao Mei | Female | 11 years | single |
| Sample | Simpson | Chao1 | Shannon |
|---|---|---|---|
| APR | 0.9654 ± 0.0147 | 4683.64 ± 969.04 | 6.8373 ± 0.7607 |
| AHR | 0.9761 ± 0.0067 | 5835.38 ± 734.65 | 7.3393 ± 0.3735 |
| Items | Df | Sums of Sqs | Mean Sqs | F. Model | R2 | Pr(>F) |
|---|---|---|---|---|---|---|
| Treat2 | 1 | 0.12481 | 0.12481 | 0.97122 | 0.08852 | 0.412 |
| Residuals | 10 | 1.28509 | 0.12851 | 0.91148 | ||
| Total | 11 | 1.40990 | 1.00000 |
| Method Name | R Statistic | p-Value | Number of Permutations |
|---|---|---|---|
| ANOSIM | 0.0722 | 0.256 | 999 |
| ID | Adduct | Name | VIP | Fold Change | p-Value | m/z | rt(s) |
|---|---|---|---|---|---|---|---|
| M173T275 | [M+H]+ | Arcaine | 15.98198784 | 6.63298872 | 0.007585569 | 173.13961 | 275.432 |
| M406T344 | [M+Na]+ | Calpain inhibitor i | 1.069037758 | 0.105522126 | 0.028039335 | 406.25683 | 344.348 |
| M316T296 | [M+H+2i]+ | Hexaconazole | 1.010155018 | 9.092924318 | 0.030197868 | 316.09947 | 296.492 |
| M431T215 | [M+Na]+ | 3beta,7beta,12beta-trihydroxy-5beta-cholan-24-oic acid | 1.302609451 | 3.620198553 | 0.031882452 | 431.27555 | 215.146 |
| M375T491_2 | [M+Na]+ | 15-ketoprostaglandin f2.alpha. | 2.406682636 | 1.815201502 | 0.034250987 | 375.22361 | 490.8645 |
| M618T37 | [M+Na]+ | 1-palmitoyl-2-oleoyl-sn-glycerol | 1.74647584 | 0.33129875 | 0.039691999 | 617.51147 | 37.1175 |
| M615T38 | [M+Na]+ | 1-palmitoyl-2-linoleoyl-rac-glycerol | 1.8091764 | 0.447178675 | 0.040387859 | 615.49474 | 37.989 |
| M218T271 | [M+H]+ | L-propionylcarnitine | 1.21936108 | 0.293714647 | 0.040439675 | 218.13761 | 271.0745 |
| M138T211 | [M+H]+ | Tyramine | 2.710329374 | 4.316237507 | 0.04166704 | 138.09027 | 210.6815 |
| M635T37 | [M+NH4]+ | 1,2-dilinoleoylglycerol | 5.638486128 | 0.171020689 | 0.042208688 | 634.53805 | 36.9215 |
| M563T168_1 | [M+H]+ | Protoporphyrin ix | 2.179714192 | 1.796078135 | 0.044486683 | 563.26432 | 167.687 |
| M131T324_2 | [M+H]+ | N-acetylputrescine | 4.312577468 | 2.540127734 | 0.047607989 | 131.11717 | 323.774 |
| M325T376 | [M+H-H2O]+ | Melibiose | 2.887633512 | 3.193731624 | 0.048006152 | 325.11229 | 375.8195 |
| M345T201 | [M-H]- | Corticosterone | 1.334636069 | 3.073188761 | 0.000006843 | 345.22681 | 200.549 |
| M607T40 | [2M-H]- | Melphalan | 1.621588379 | 0.046578615 | 0.000467639 | 607.14828 | 40.001 |
| M392T336 | [3M-H]- | Guanidinopropionic acid | 1.572600471 | 0.005931119 | 0.001473817 | 392.19985 | 336.089 |
| M706T175 | [M+CH3COOH-H]- | Sm d30:1 | 1.479218847 | 0.04733475 | 0.001823034 | 705.51643 | 174.804 |
| M879T235 | [M-H]- | Pi 38:7 | 1.404372826 | 0.013405137 | 0.01078511 | 879.49408 | 234.755 |
| M1082T38 | [M-H]- | Furostane base + o-hex, o-hex-hex-hex | 1.234234036 | 15.77960671 | 0.011283726 | 1081.52055 | 38.444 |
| M347T37 | [M-H]- | 3(2’-chlorophenyl)-7-hydroxy-4-phenylcoumarin | 2.42651536 | 0.438614595 | 0.011422429 | 347.04313 | 37.466 |
| M779T44 | [M-H]- | Pe(16:0/19,20-epdpe) | 2.281257798 | 3.036982278 | 0.013858763 | 778.51345 | 43.697 |
| M566T401 | [3M-H]- | .alpha.-guanidinoglutaric acid | 1.265640564 | 0.014467482 | 0.015477295 | 566.21791 | 400.6375 |
| M416T444 | [M-H]- | Thymopoietin ii (32-34) | 1.260199839 | 0.02135598 | 0.01605087 | 416.2132 | 443.875 |
| M690T288 | [M+Cl]- | 7-benzyl-11,14-dimethyl-16-(2-methylpropyl)-10,13-di(propan-2-yl)-17-oxa-1,5,8,11,14-pentazabicyclo[17.3.0]docosane-2,6,9,12,15,18-hexone | 1.289120759 | 0.004845138 | 0.016215125 | 690.36559 | 287.696 |
| M774T44 | [M-H-NH3]- | 1,2-distearoyl-sn-glycero-3-phospho-l-serine | 1.217162339 | 1.999427667 | 0.026779114 | 773.53 | 44.129 |
| M292T298 | [M-H]- | N-acetylmuramic acid | 1.021977643 | 9.110003872 | 0.030490542 | 292.10291 | 298.445 |
| M73T372_2 | [M-H]- | Propionic acid | 1.666044111 | 1.931336222 | 0.032458363 | 73.0297 | 372.0775 |
| M117T372_2 | [M-H]- | Methylmalonic acid | 4.081076931 | 1.994118874 | 0.036897873 | 117.01951 | 371.987 |
| M212T34 | [M-H]- | Indoxyl sulfate | 2.006047308 | 3.797263074 | 0.037033194 | 212.00198 | 34.181 |
| M174T378_2 | [M-H]- | N-acetyl-l-aspartic acid | 2.274041379 | 4.23154483 | 0.037634414 | 174.04059 | 377.882 |
| M436T132_2 | [M-H]- | Lunarine | 1.143728194 | 3.927292997 | 0.037699018 | 436.23573 | 132.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Wang, X.; Ayala, J.; Liu, Y.; An, J.; Wang, D.; Cai, Z.; Hou, R.; Zhang, M. Possible Effects of Early Maternal Separation on the Gut Microbiota of Captive Adult Giant Pandas. Animals 2022, 12, 2587. https://doi.org/10.3390/ani12192587
Zhang X, Wang X, Ayala J, Liu Y, An J, Wang D, Cai Z, Hou R, Zhang M. Possible Effects of Early Maternal Separation on the Gut Microbiota of Captive Adult Giant Pandas. Animals. 2022; 12(19):2587. https://doi.org/10.3390/ani12192587
Chicago/Turabian StyleZhang, Xiaohui, Xueying Wang, James Ayala, Yuliang Liu, Junhui An, Donghui Wang, Zhigang Cai, Rong Hou, and Mingyue Zhang. 2022. "Possible Effects of Early Maternal Separation on the Gut Microbiota of Captive Adult Giant Pandas" Animals 12, no. 19: 2587. https://doi.org/10.3390/ani12192587
APA StyleZhang, X., Wang, X., Ayala, J., Liu, Y., An, J., Wang, D., Cai, Z., Hou, R., & Zhang, M. (2022). Possible Effects of Early Maternal Separation on the Gut Microbiota of Captive Adult Giant Pandas. Animals, 12(19), 2587. https://doi.org/10.3390/ani12192587