
Wild Boar Effects on Fungal Abundance and Guilds from Sporocarp Sampling in a Boreal Forest Ecosystem
 ,
,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Study Area
2.2. Monitoring Design
- (1)
- We randomly placed a transect in each of the 11 study sites. In these transects, we recorded the rooting intensity per site as a percentage of soil disturbance along the 200 m long transects [11]. The rate of soil rooting was calculated as follows; a fixed bandwidth of 1 m was established and each rooting length was scored within this band [9,15].
- (2)
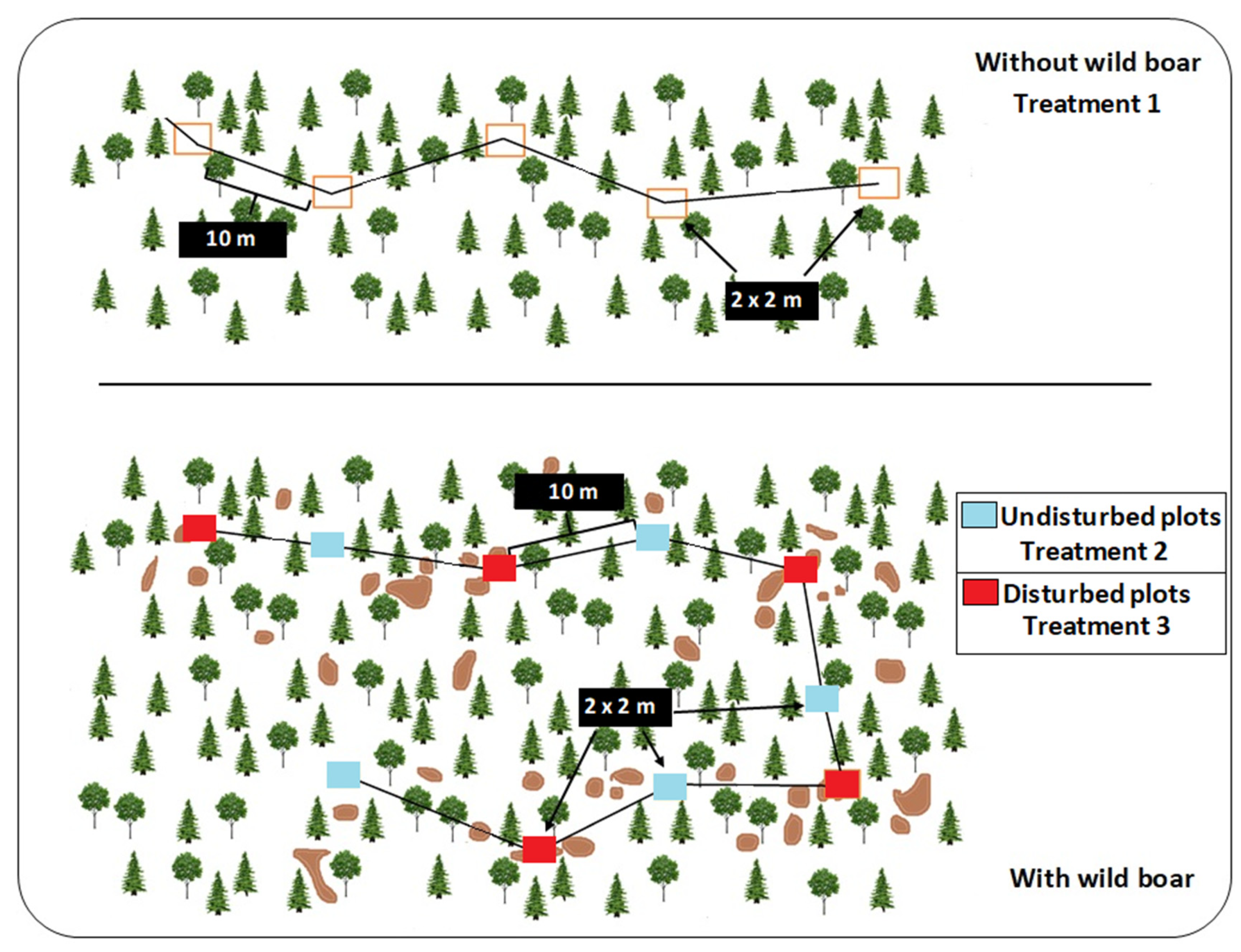
- The plots were square sampling areas of 2 × 2 m, separated by a minimum of 10 m. Ten plots per site were established in areas with wild boar presence, five in disturbed areas (i.e., with rooting) and five in undisturbed (no rooting) areas. Disturbed and undisturbed plots were placed alternately along the transect (see Figure 2). If, after a distance of 10 m from a previous plot, for instance, a disturbed area, we did not find an area free from wild boar disturbance, we continued until we found an undisturbed area. In areas without presence of wild boar, five plots were sampled in a similar fashion, that is, square sampling areas (2 × 2 m) separated by a minimum of 10 m. In each sampled rooting/disturbed plot, we estimated the rooting intensity as the percentage (as to the closest 10%) of the area of soil altered by wild boar. Therefore, we established three wild boar treatments: (1) without wild boar presence; (2) potentially with animals but no signs of rooting; and (3) with animals and rooting signs (Figure 2). Species richness and abundance of each fungal guild were also measured at the plot level. Each 2 × 2 m plot was monitored by recording and photographing the fungal sporocarps.
2.3. Abiotic Variables
- (A)
- Forest type. Classified as closed forest (>70% of forest cover), semi-open forest (between 30–70% of forest cover) and open forest (<30% of forest cover) [63]. The degree of canopy openness in a boreal forest directly influences the amount of light the forest floors receive, subsequently affecting plant composition and air and soil humidity, which are critical conditions for fungal communities [45,64,65].
- (B)
- (C)
- Soil humidity. Dry with a soil moisture < 50% and wet > 50%. The humidity of soil is related not only to foliar coverage but also to soil texture and climatic conditions, such as the amount of precipitation and wind [61,68]. We estimated the soil moisture based on wetness on the ground, which in most cases was related to nearness to the River Dalälven or other wetter areas such as bogs or fens commonly intermingled in the boreal forest.
2.4. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Massei, G.; Kindberg, J.; Licoppe, A.; Gačić, D.; Šprem, N.; Kamler, J.; Baubet, E.; Hohmann, U.; Monaco, A.; Ozoliņš, J.; et al. Wild boar populations up, numbers of hunters down? A review of trends and implications for Europe. Pest Manag. Sci. 2015, 71, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Lagos, L.; Picos, J.; Valero, E. Temporal pattern of wild ungulate-related traffic accidents in northwest Spain. Eur. J. Wildl. Res. 2012, 58, 661–668. [Google Scholar] [CrossRef]
- Herrero, J.; García-Serrano, A.; Couto, S.; Ortuño, V.M.; García-González, R. Diet of wild boar Sus scrofa L. and crop damage in an intensive agroecosystem. Eur. J. Wildl. Res. 2006, 52, 245–250. [Google Scholar] [CrossRef]
- Meng, X.J.; Lindsay, D.S.; Sriranganathan, N. Wild boars as sources for infectious diseases in livestock and humans. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2697–2707. [Google Scholar] [CrossRef]
- Gortázar, C.; Delahay, R.J.; Mcdonald, R.A.; Boadella, M.; Wilson, G.J.; Gavier-Widen, D.; Acevedo, P. The status of tuberculosis in European wild mammals. Mamm. Rev. 2012, 42, 193–206. [Google Scholar] [CrossRef]
- De la Torre, A.; Bosch, J.; Iglesias, I.; Muñoz, M.J.; Mur, L.; Martínez-López, B.; Martínez, M.; Sánchez-Vizcaíno, J.M. Assessing the risk of African swine fever introduction into the European Union by wild boar. Transbound. Emerg. Dis. 2015, 62, 272–279. [Google Scholar] [CrossRef]
- Bueno, C.G.; Alados, C.L.; Gómez-García, D.; Barrio, I.C.; García-González, R. Understanding the main factors in the extent and distribution of wild boar rooting on alpine grasslands. J. Zool. 2009, 279, 195–202. [Google Scholar] [CrossRef]
- Bueno, C.G.; Reiné, R.; Alados, C.L.; Gómez-García, D. Effects of large wild boar disturbances on alpine soil seed banks. Basic Appl. Ecol. 2011, 12, 125–133. [Google Scholar] [CrossRef]
- Carpio, A.J.; Guerrero-Casado, J.; Ruiz-Aizpurua, L.; Vicente, J.; Tortosa, F.S. The high abundance of wild ungulates in a Mediterranean region: Is this compatible with the European rabbit? Wildl. Biol. 2014, 20, 161–166. [Google Scholar] [CrossRef]
- Carpio, A.J.; Guerrero-Casado, J.; Tortosa, F.S.; Vicente, J. Predation of simulated red-legged partridge nests in big game estates from South Central Spain. Eur. J. Wildl. Res. 2014, 60, 391–394. [Google Scholar] [CrossRef][Green Version]
- Carpio, A.J.; Hillström, L.; Tortosa, F.S. Effects of wild boar predation on nests of wading birds in various Swedish habitats. Eur. J. Wildl. Res. 2016, 62, 423–430. [Google Scholar] [CrossRef]
- Barrios-Garcia, M.N.; Ballar, S.A. Impact of wild boar (Sus scrofa) in its introduced and native range: A review. Biol. Invasions 2012, 14, 2283–2300. [Google Scholar] [CrossRef]
- Mohr, D.; Cohnstaedt, L.W.; Topp, W. Wild boar and red deer affect soil nutrients and soil biota in steep oak stands of the Eifel. Soil Biol. Biochem. 2005, 37, 693–700. [Google Scholar] [CrossRef]
- Bueno, C.G.; Jiménez, J.J. Livestock grazing activities and wild boar rooting affect alpine earthworm communities in the Central Pyrenees (Spain). Appl. Soil Ecol. 2014, 83, 71–78. [Google Scholar] [CrossRef]
- Bueno, C.G.; Barrio, I.C.; García-González, R.; Alados, C.L.; Gómez-García, D. Does wild boar rooting affect livestock grazing areas in alpine grasslands? Eur. J. Wildl. Res. 2010, 56, 765–770. [Google Scholar] [CrossRef]
- Cuevas, M.F.; Novillo, A.; Campos, C.; Dacar, M.A.; Ojeda, R.A. Food habits and impact of rooting behaviour of the invasive wild boar, Sus scrofa, in a protected area of the Monte Desert, Argentina. J. Arid Environ. 2010, 74, 1582–1585. [Google Scholar] [CrossRef]
- Welander, J. Are wild boars a future threat to the Swedish flora? J. Mt. Ecol. 1995, 3, 165–167. [Google Scholar]
- Sousa, W.P. The role of disturbance in natural communities. Annu. Rev. Ecol. Evol. Syst. 1984, 15, 353–391. [Google Scholar] [CrossRef]
- Jonsson, B.G.; Esseen, P.A. Treefall disturbance maintains high bryophyte diversity in a boreal spruce forest. J. Ecol. 1990, 78, 924–936. [Google Scholar] [CrossRef]
- Lavorel, S.; Lepart, J.; Debussche, M.; Lebreton, J.D.; Beffy, J.L. Small scale disturbances and the maintenance of species diversity in Mediterranean old fields. Oikos 1994, 70, 455–473. [Google Scholar] [CrossRef]
- Welander, J. Spatial and temporal dynamics of wild boar (Sus scrofa) rooting in a mosaic landscape. J. Zool. 2000, 252, 263–271. [Google Scholar] [CrossRef]
- Fabiańska, I.; Sosa-Lopez, E.; Bucher, M. The role of nutrient balance in shaping plant root-fungal interactions: Facts and speculation. Curr. Opin. Microbiol. 2019, 49, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Reich, P.B.; Trivedi, C.; Eldridge, D.J.; Abades, S.; Alfaro, F.D.; Bastida, F.; Berhe, A.A.; Cutler, N.A.; Gallardo, A. Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat. Ecol. Evol. 2020, 4, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Diagne, N.; Ngom, M.; Djighaly, P.I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of arbuscular mycorrhizal fungi on plant growth and performance: Importance in biotic and abiotic stressed regulation. Divers 2020, 12, 370. [Google Scholar] [CrossRef]
- Rodriguez-Ramos, J.C.; Cale, J.A.; Cahill, J.F., Jr.; Simard, S.W.; Karst, J.; Erbilgin, N. Changes in soil fungal community composition depend on functional group and forest disturbance type. New Phytol. 2020, 229, 1105–1117. [Google Scholar] [CrossRef]
- Cho, H.; Kim, M.; Tripathi, B.; Adams, J. Changes in soil fungal community structure with increasing disturbance frequency. Microb. Ecol. 2017, 74, 62–77. [Google Scholar] [CrossRef]
- Kabir, Z. Tillage or no-tillage: Impact on mycorrhizae. Can. J. Plant Sci. 2005, 85, 23–29. [Google Scholar] [CrossRef]
- Jansa, J.; Wiemken, A.; Frossard, E. The effects of agricultural practices on arbuscular mycorrhizal fungi. Geol. Soc. Spec. Publ. 2006, 266, 89–115. [Google Scholar] [CrossRef]
- Schnoor, T.K.; Lekberg, Y.; Rosendahl, S.; Olsson, P.A. Mechanical soil disturbance as a determinant of arbuscular mycorrhizal fungal communities in semi-natural grassland. Mycorrhiza 2011, 21, 211–220. [Google Scholar] [CrossRef]
- Säle, V.; Aguilera, P.; Laczko, E.; Mäder, P.; Berner, A.; Zihlmann, U.; van der Heijden, M.G.A.; Oehl, F. Impact of conservation tillage and organic farming on the diversity of arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2015, 84, 38–52. [Google Scholar] [CrossRef]
- Trappe, M.J.; Cromack, K.; Trappe, J.M.; Wilson, J.; Rasmussen, M.C.; Castellano, M.A.; Miller, S.L. Relationships of current and past anthropogenic disturbance to mycorrhizal sporocarp fruiting patterns at Crater Lake National Park, Oregon. Can. J. For. Res. 2009, 39, 1662–1676. [Google Scholar] [CrossRef]
- Kranabetter, J.M.; Haeussler, S.; Wood, C. Vulnerability of boreal indicators (ground-dwelling beetles, understory plants and ectomycorrhizal fungi) to severe forest soil disturbance. For. Ecol. Manag. 2017, 402, 213–222. [Google Scholar] [CrossRef]
- Heinken, T.; Lees, R.; Raudnitschka, D.; Runge, S. Epizoochorous dispersal of bryophyte stem fragments by roe deer (Capreolus capreolus) and wild boar (Sus scrofa). J. Bryol. 2001, 23, 293–300. [Google Scholar] [CrossRef]
- Heinken, T.; Schmidt, M.; von Oheimb, G.; Kriebitzsch, W.U.; Ellenberg, H. Soil seed banks near rubbing trees indicate dispersal of plant species into forests by wild boar. Basic Appl. Ecol. 2006, 7, 31–44. [Google Scholar] [CrossRef]
- Dovrat, G.; Perevolotsky, A.; Ne’eman, G. Wild boars as seed dispersal agents of exotic plants from agricultural lands to conservation areas. J. Arid Environ. 2012, 78, 49–54. [Google Scholar] [CrossRef]
- Nuñez, M.A.; Hayward, J.; Horton, T.R.; Amico, G.C.; Dimarco, R.D.; Barrios-Garcia, M.N.; Simberloff, D. Exotic mammals disperse exotic fungi that promote invasion by exotic trees. PLoS ONE 2013, 8, e66832. [Google Scholar] [CrossRef]
- Livne-Luzon, S.; Avidan, Y.; Weber, G.; Migael, H.; Bruns, T.; Ovadia, O.; Shemesh, H. Wild boars as spore dispersal agents of ectomycorrhizal fungi: Consequences for community composition at different habitat types. Mycorrhiza 2017, 27, 165–174. [Google Scholar] [CrossRef]
- Soteras, F.; Ibarra, C.; Geml, J.; Barrios-García, M.N.; Domínguez, L.S.; Nouhra, E.R. Mycophagy by invasive wild boar (Sus scrofa) facilitates dispersal of native and introduced mycorrhizal fungi in Patagonia, Argentina. Fungal Ecol. 2017, 26, 51–58. [Google Scholar] [CrossRef]
- Nuske, S.J.; Vernes, K.; May, T.W.; Claridge, A.W.; Congdon, B.C.; Krockenberger, A.; Abell, S.E. Redundancy among mammalian fungal dispersers and the importance of declining specialists. Fungal Ecol. 2017, 27, 1–13. [Google Scholar] [CrossRef]
- Read, D.J.; Leake, J.R.; Perez-Moreno, J. Mycorrhizal fungi as drivers of ecosystem processes in heathland and boreal forest biomes. Can. J. Bot. 2004, 82, 1243–1263. [Google Scholar] [CrossRef]
- Bueno, C.G.; Azorín, J.; Gómez-García, D.; Alados, C.L.; Badía, D. Occurrence and intensity of wild boar disturbances, effects on the physical and chemical soil properties of alpine grasslands. Plant Soil 2013, 373, 243–256. [Google Scholar] [CrossRef]
- Piattoni, F.; Oir, F.; Morara, M.; Iotti, M.; Zambonelli, A. The role of wild boars in spore dispersal of hypogeous fungi. Acta Mycol. 2012, 47, 145–153. [Google Scholar] [CrossRef][Green Version]
- Herrero, J.; Irizar, I.; Laskurain, N.A.; García-Serrano, A.; García-González, R. Fruits and roots: Wild boar foods during the cold season in the southwestern Pyrenees. Ital. J. Zool. 2005, 72, 49–52. [Google Scholar] [CrossRef]
- Summerbell, R.C. Root endophyte and mycorrhizosphere fungi of black spruce, Picea mariana, in a boreal forest habitat: Influence of site factors on fungal distributions. Stud. Mycol. 2005, 53, 121–145. [Google Scholar] [CrossRef]
- Gómez-Hernández, M.; Williams-Linera, G.; Guevara, R.; Lodge, D.J. Patterns of macromycete community assemblage along an elevation gradient: Options for fungal gradient and metacommunity analyse. Biodivers. Conserv. 2012, 21, 2247–2268. [Google Scholar] [CrossRef]
- Deepika, S.; Kothamasi, D. Soil moisture—A regulator of arbuscular mycorrhizal fungal community assembly and symbiotic phosphorus uptake. Mycorrhiza 2015, 25, 67–75. [Google Scholar] [CrossRef]
- Goldmann, K.; Schröter, K.; Pena, R.; Schöning, I.; Schrumpf, M.; Buscot, F.; Polle, A.; Wubet, T. Divergent habitat filtering of root and soil fungal communities in temperate beech forests. Sci. Rep. 2016, 6, 31439. [Google Scholar] [CrossRef]
- Swedish Forest Agency 2010. Available online: https://www.skogsstyrelsen.se/en/ (accessed on 1 December 2018).
- Viltdata. Available online: www.viltdata.se (accessed on 1 December 2018).
- Thurfjell, H.; Ball, J.P.; Åhlén, P.A.; Kornacher, P.; Dettki, H.; Sjöberg, K. Habitat use and spatial patterns of wild boar Sus scrofa (L.): Agricultural fields and edges. Eur. J. Wildl. Res. 2009, 55, 517–523. [Google Scholar] [CrossRef]
- Fattorini, N.; Ferretti, F. Estimating wild boar density and rooting activity in a Mediterranean protected area. Mamm. Biol. 2020, 100, 241–251. [Google Scholar] [CrossRef]
- Wästerlund, I.; Ingelög, T. Fruit body production of larger fungi in some young Swedish forests with special reference to logging waste. For. Ecol. Manag. 1980, 3, 269–294. [Google Scholar] [CrossRef]
- Nylén, B. Svampar i Nordenoch Europa; Natur och kultur/LT: Stockholm, Sweden, 2000; 704p. [Google Scholar]
- FUNGuild Online Database. Available online: http://www.funguild.org/ (accessed on 1 December 2018).
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Põlme, S.; Abarenkov, K.; Henrik Nilsson, R.; Lindahl, B.D.; Clemmensen, K.E.; Kauserud, H.; Nguyen, N.; Kjøller, R.; Bates, S.T.; Baldrian, P.; et al. FungalTraits: A user-friendly traits database of fungi and fungus-like stramenopiles. Fungal Divers. 2020, 105, 1–16. [Google Scholar] [CrossRef]
- Zanne, A.E.; Abarenkov, K.; Afkhami, M.E.; Aguilar-Trigueros, C.A.; Bates, S.; Bhatnagar, J.M.; Busby, P.E.; Christian, N.; Cornwell, W.; Crowther, T.W.; et al. Fungal functional ecology: Bringing a trait-based approach to plant-associated fungi. Biol. Rev. 2020, 95, 409–433. [Google Scholar] [CrossRef] [PubMed]
- Dahlberg, A.; Jonsson, L.; Nylund, J.E. Species diversity and distribution of biomass above and below ground among ectomycorrhizal fungi in an old-growth Norway spruce forest in south Sweden. Can. J. Bot. 1997, 75, 1323–1335. [Google Scholar] [CrossRef]
- Kernaghan, G.; Harper, K.A. Community structure of ectomycorrhizal fungi across an alpine/subalpine ecotone. Ecography 2001, 24, 181–188. [Google Scholar] [CrossRef]
- Danks, M.; Lebel, T.; Vernes, K.; Andrew, N. Truffle-like fungi sporocarps in a eucalypt-dominated landscape: Patterns in diversity and community structure. Fungal Divers. 2013, 58, 143–157. [Google Scholar] [CrossRef]
- Solly, E.F.; Lindahl, B.D.; Dawes, M.A.; Peter, M.; Souza, R.C.; Rixen, C.; Hagedorn, F. Experimental soil warming shifts the fungal community composition at the alpine treeline. New Phytol. 2017, 215, 766–778. [Google Scholar] [CrossRef]
- Ruiz-Almenara, C.; Gándara, E.; Gómez-Hernández, M. Comparison of diversity and composition of macrofungal species between intensive mushroom harvesting and non-harvesting areas in Oaxaca, Mexico. PeerJ 2019, 7, e8325. [Google Scholar] [CrossRef]
- Łuczaj, Ł.; Sadowska, B. Edge effect in different groups of organisms: Vascular plant, bryophyte and fungi species richness across a forest-grassland border. Folia Geobot. Phytotaxon. 1997, 32, 343–353. [Google Scholar] [CrossRef]
- Junninen, K.; Similä, M.; Kouki, J.; Kotiranta, H. Assemblages of wood-inhabiting fungi along the gradients of succession and naturalness in boreal pine-dominated forests in Fennoscandia. Ecography 2006, 29, 75–83. [Google Scholar] [CrossRef]
- Caiafa, M.V.; Gómez-Hernández, M.; Williams-Linera, G.; Ramírez-Cruz, V. Functional diversity of macromycete communities along an environmental gradient in a Mexican seasonally dry tropical forest. Fungal Ecol. 2017, 28, 66–75. [Google Scholar] [CrossRef]
- Cousin, I.; Nicoullaud, B.; Coutadeur, C. Influence of rock fragments on the water retention and water percolation in a calcareous soil. Catena 2003, 53, 97–114. [Google Scholar] [CrossRef]
- Hlaváčiková, H.; Novák, V. A relatively simple scaling method for describing the unsaturated hydraulic functions of stony soils. J. Plant Nutr. Soil Sci. 2014, 177, 560–565. [Google Scholar] [CrossRef]
- Dighton, J.; White, J.F.; White, J.; Oudemans, P. The Fungal Community: Its Organization and Role in the Ecosystem, 3rd ed.; CRC: Boca Raton, FL, USA; Taylor & Francis: New York, NY, USA, 2005. [Google Scholar]
- Fleming, P.A.; Anderson, H.; Prendergast, A.S.; Bretz, M.R.; Valentine, L.E.; Hardy, G.E.S. Is the loss of Australian digging mammals contributing to a deterioration in ecosystem function? Mamm. Rev. 2014, 44, 94–108. [Google Scholar] [CrossRef]
- Gehring, C.A.; Wolf, J.E.; Theimer, T.C. Terrestrial vertebrates promote arbuscular mycorrhizal fungal diversity and inoculum potential in a rain forest soil. Ecol. Lett. 2002, 5, 540–548. [Google Scholar] [CrossRef]
- Davies, G.T.O.; Kirkpatrick, J.B.; Cameron, E.Z.; Carver, S.; Johnson, C.N. Ecosystem engineering by digging mammals: Effects on soil fertility and condition in Tasmanian temperate woodland. R. Soc. Open Sci. 2019, 6, 180621. [Google Scholar] [CrossRef]
- Miranda, V.; Rothen, C.; Yela, N.; Aranda-Rickert, A.; Barros, J.; Calcagno, J.; Fracchia, S. Subterranean desert rodents (genus Ctenomys) create soil patches enriched in root endophytic fungal propagules. Microb. Ecol. 2019, 77, 451–459. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Zechmeister-Boltenstern, S.; Richter, A.A. Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources. Front. Microbiol. 2014, 5, 1–10. [Google Scholar] [CrossRef]
- Chen, W.; Wang, J.; Meng, Z.; Xu, R.; Chen, J.; Zhang, Y.; Hu, T. Fertility-related interplay between fungal guilds underlies plant richness-productivity relationships in natural grasslands. New Phytol. 2020, 226, 1129–1143. [Google Scholar] [CrossRef]
- Kranabetter, J.M.; Harman-Denhoed, R.; Hawkins, B.J. Saprotrophic and ectomycorrhizal fungal sporocarp stoichiometry (C: N: P) across temperate rainforests as evidence of shared nutrient constraints among symbionts. New Phytol. 2019, 221, 482–492. [Google Scholar] [CrossRef]
- Carpio, A.J.; Oteros, J.; Lora, A.; Tortosa, F.S. Effects of the overabundance of wild ungulates on natural grassland in Southern Spain. Agrofor. Syst. 2015, 89, 637–644. [Google Scholar] [CrossRef]
- Singer, F.J.; Swank, W.T.; Clebsch, E.E. Effects of wild pig rooting in a deciduous forest. J. Wildl. Manag. 1984, 48, 464–473. [Google Scholar] [CrossRef]
- Johnson, N.C. Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales. New Phytol. 2010, 185, 631–647. [Google Scholar] [CrossRef]
- Amaranthus, M.P.; Page-Dumroese, D.S.; Harvey, A.; Cazares, E.; Bednar, L.F. Soil Compaction and Organic Matter Affect Conifer Seedling Nonmycorrhizal and Ectomycorrhizal Root Tip Abundance and Diversity; Res. Pap. PNW-RP-494; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1996; 12p. [Google Scholar]
- Page-Dumroese, D.S.; Harvey, A.E.; Jurgensen, M.F.; Amaranthus, M.P. Impacts of soil compaction and tree stump removal on soil properties and outplanted seedlings in northern Idaho, USA. Can. J. Soil Sci. 1998, 78, 29–34. [Google Scholar] [CrossRef]
- Palacio, S.; Bueno, C.G.; Azorín, J.; Maestro, M.; Gómez-García, D. Wild-boar disturbance increases nutrient and C stores of geophytes in subalpine grasslands. Am. J. Bot. 2013, 100, 1790–1799. [Google Scholar] [CrossRef]
- Vahter, T.; Bueno, C.G.; Davison, J.; Herodes, K.; Hiiesalu, I.; Kasari-Toussaint, L.; Oja, J.; Olsson, P.A.; Siim-Kaarel, S.; Zobel, M.; et al. Co-introduction of native mycorrhizal fungi and plant seeds accelerates restoration of post-mining landscapes. J. Appl. Ecol. 2020, 57, 1741–1751. [Google Scholar] [CrossRef]
- Welander, J. Spatial and temporal dynamic of a disturbance regime: Wild boar (Sus scrofa L.) rooting and its effects on plant species diversity. In Department of Conservation Biology; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2000. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Level | ||
| Models | Predictors | Response variable |
| Model 1a | Presence/absence wild boar | Abundance of fungal sporocarps |
| Model 1b | Presence/absence rooting | Abundance of fungal sporocarps |
| Plot level | ||
| Model 2 | Treatment/environmental variables | Abundance of fungal sporocarps |
| Model 3 | Treatment/environmental variables | Abundance of symbiotic |
| Model 4 | Treatment/environmental variables | Abundance of saprophytic |
| Model 5 | Treatment/environmental variables | Abundance of pathogenic |
| Variable | df | F-Value | Coefficient ± E.S. |
|---|---|---|---|
| Abundance of fungi (model 2) | |||
| Intercept | 1 | 19.16 *** | 4.14 ± 0.22 |
| Treatment | 2 | 8.39 * |
Wild boar no rooting = −1.41 ± 0.14 No wild boar = −1.49 ± 0.29 |
| Type of soil | 1 | 0.18 | Soft = 0.09 ± 0.21 |
| Humidity | 1 | 46.42 *** | Dry = −0.88 ± 0.13 |
| Forest type | 2 | 24.05 *** |
Semi-open = 0.56 ± 0.13 Closed = 0.33 ± 0.15 |
| Variable | df | F-Value | Coefficient ± E.S. |
|---|---|---|---|
| Abundance of symbiotic (model 3) | |||
| Treatment | 2 | 5.75 ** |
Wild boar no rooting = 0.40 ± 0.12 No wild boar = 0.53 ± 0.88 |
| Forest type | 2 | 26.72 *** | Closed = 1.30 ± 0.32 Semi-open = −0.23 ± 0.29 |
| Type of soil | 1 | 3.64 | Rocky = −1.29 ± 0.68 |
| Humidity | 1 | 12.15 *** | Dry = −0.93 ± 0.27 |
| Abundance of saprophytic (model 4) | |||
| Treatment | 2 | 25.1 *** |
Wild boar no rooting = −0.66 ± 0.09 No wild boar = −1.19 ± 0.68 |
| Forest type | 2 | 42.78 *** | Closed = −0.45 ± 0.21 Semi-open = 1.63 ± 0.19 |
| Type of soil | 1 | 3.71 | Rocky = −1.47 ± 0.78 |
| Humidity | 1 | 15.21 ** | Dry = −0.70 ± 0.18 |
| Abundance of pathogenic (model 5) | |||
| Treatment | 2 | 3.98 * |
Wild boar no rooting = 1.87 ± 0.75 No wild boar = 0.81 ± 1.00 |
| Forest type | 2 | 4.01 * | Closed = 1.19 ± 1.31 Semi-open = 2.80 ± 1.00 |
| Type of soil | 1 | 1.2 | Rocky = 0.84 ± 0.77 |
| Humidity | 1 | 3.09 | Dry = −1.16 ± 0.66 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carpio, A.J.; García, M.; Hillström, L.; Lönn, M.; Carvalho, J.; Acevedo, P.; Bueno, C.G. Wild Boar Effects on Fungal Abundance and Guilds from Sporocarp Sampling in a Boreal Forest Ecosystem. Animals 2022, 12, 2521. https://doi.org/10.3390/ani12192521
Carpio AJ, García M, Hillström L, Lönn M, Carvalho J, Acevedo P, Bueno CG. Wild Boar Effects on Fungal Abundance and Guilds from Sporocarp Sampling in a Boreal Forest Ecosystem. Animals. 2022; 12(19):2521. https://doi.org/10.3390/ani12192521
Chicago/Turabian StyleCarpio, Antonio J., Marta García, Lars Hillström, Mikael Lönn, Joao Carvalho, Pelayo Acevedo, and C. Guillermo Bueno. 2022. "Wild Boar Effects on Fungal Abundance and Guilds from Sporocarp Sampling in a Boreal Forest Ecosystem" Animals 12, no. 19: 2521. https://doi.org/10.3390/ani12192521
APA StyleCarpio, A. J., García, M., Hillström, L., Lönn, M., Carvalho, J., Acevedo, P., & Bueno, C. G. (2022). Wild Boar Effects on Fungal Abundance and Guilds from Sporocarp Sampling in a Boreal Forest Ecosystem. Animals, 12(19), 2521. https://doi.org/10.3390/ani12192521