
Stress-Induced Immunosuppression Affects Immune Response to Newcastle Disease Virus Vaccine via Circulating miRNAs

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Grouping and Tissues Collection
2.2. Antibody Level Determination and Organ Coefficient Analysis
2.3. Reverse Transcription and Quantitative Real-Time PCR (qRT-PCR)
2.4. Bioinformatics Analysis
2.5. Statistical Analysis
3. Results
3.1. Analysis of Organ Coefficient and Serum Antibody
3.2. Differential Expression Analysis of Serum Circulating miRNAs between Dex Group and Control Group
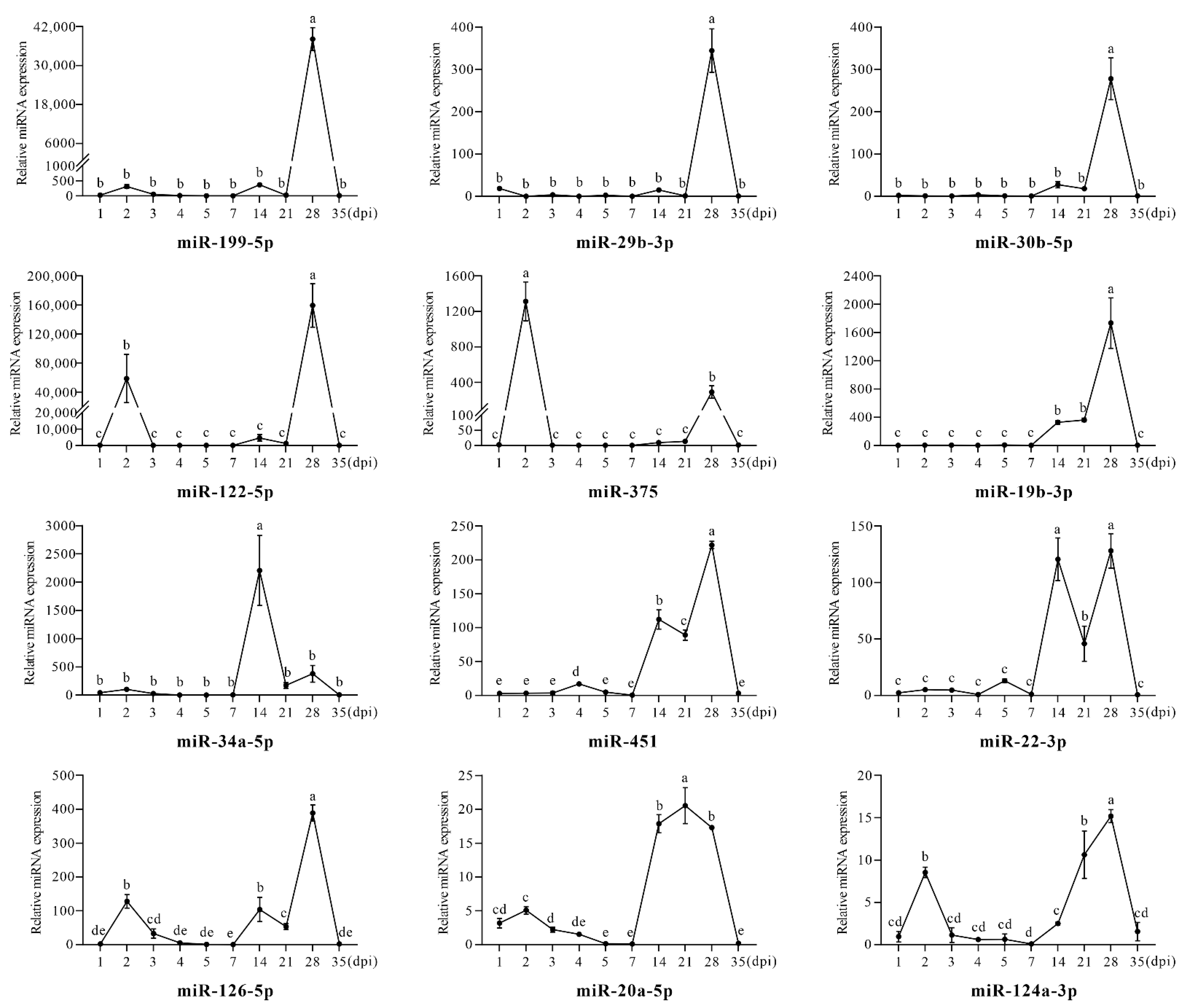
3.3. The Differential Regulation of Circulating miRNAs in the Process of Stress-Induced Immunosuppression Affecting Immune Response to NDV Vaccine
3.4. Functional Analysis and Pathway Prediction of Candidate Circulating miRNAs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Matur, E.; Akyazi, İ.; Eraslan, E.; Ergul Ekiz, E.; Eseceli, H.; Keten, M.; Metiner, K.; Aktaran Bala, D. The effects of environmental enrichment and transport stress on the weights of lymphoid organs, cell-mediated immune response, heterophil functions and antibody production in laying hens. Anim. Sci. J. 2016, 87, 284–292. [Google Scholar] [CrossRef] [PubMed]
- El-Kassas, S.; Abdo, S.E.; El-Naggar, K.; Abdo, W.; Kirrella, A.A.; Nashar, T.O. Ameliorative effect of dietary supplementation of copper oxide nanoparticles on inflammatory and immune reponses in commercial broiler under normal and heat-stress housing conditions. J. Therm. Biol. 2018, 78, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Ding, M.; Guo, Y.; Su, A.; Zhai, M.; Tian, Y.; Li, K.; Sun, G.; Jiang, R.; Han, R. Use of transcriptomic analysis to identify microRNAs related to the effect of stress on thymus immune function in a chicken stress model. Res. Vet. Sci. 2021, 140, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Su, A.; Guo, Y.; Tian, H.; Zhou, Y.; Li, W.; Tian, Y.; Li, K.; Sun, G.; Jiang, R.; Yan, F. Analysis of miRNA and mRNA reveals core interaction networks and pathways of dexamethasone-induced immunosuppression in chicken bursa of Fabricius. Mol. Immunol. 2021, 134, 34–47. [Google Scholar] [CrossRef]
- Zhen, Y.; Wang, F.; Liang, W.; Liu, J.; Gao, G.; Wang, Y.; Xu, X.; Su, Q.; Zhang, Q.; Liu, B. Identification of differentially expressed non-coding RNA in porcine alveolar macrophages from Tongcheng and large white pigs responded to PRRSV. Sci. Rep. 2018, 8, 15621. [Google Scholar] [CrossRef]
- Yoshikawa, Y.; Hirata, N.; Terada, H.; Sawashita, Y.; Yamakage, M. Identification of candidate genes and pathways in dexmedetomidine-induced cardioprotection in the rat heart by bioinformatics analysis. Int. J. Mol. Sci. 2019, 20, 1614. [Google Scholar] [CrossRef]
- Muneer, M.; Farah, I.; Newman, J.; Goyal, S. Immunosuppression in animals. Br. Vet. J. 1988, 144, 288–301. [Google Scholar] [CrossRef]
- Nie, F.; Zhang, J.; Li, M.; Chang, X.; Duan, H.; Li, H.; Zhou, J.; Ji, Y.; Guo, L. Transcriptome analysis of thymic tissues from Chinese Partridge Shank chickens with or without Newcastle disease virus LaSota vaccine injection via high-throughput RNA sequencing. Bioengineered 2022, 13, 9131–9144. [Google Scholar] [CrossRef]
- Jia, Y.-Q.; Wang, X.-L.; Wang, X.-W.; Yan, C.-Q.; Lv, C.-J.; Li, X.-Q.; Chu, Z.-L.; Adam, F.E.A.; Xiao, S.; Zhang, S.-X. Common microRNA–mRNA interactions in different Newcastle disease virus-infected chicken embryonic visceral tissues. Int. J. Mol. Sci. 2018, 19, 1291. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, W.; Xu, H.; Liu, J.; Deng, Y.; Cheng, H.; Zhu, S.; Pei, Y.; Hu, J.; Hu, Z. MicroRNA expression profiling in Newcastle disease virus-infected DF-1 cells by deep sequencing. Front. Microbiol. 2019, 10, 1659. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Jia, Y.; Ren, J.; Liu, H.; Xiao, S.; Wang, X.; Yang, Z. MicroRNA gga-miR-455-5p suppresses Newcastle disease virus replication via targeting cellular suppressors of cytokine signaling 3. Vet. Microbiol. 2019, 239, 108460. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jia, Y.; Wang, X.; Wang, C.; Lv, C.; Li, X.; Chu, Z.; Han, Q.; Xiao, S.; Zhang, S. MiR-375 has contrasting effects on Newcastle disease virus growth depending on the target gene. Int. J. Biol. Sci. 2019, 15, 44. [Google Scholar] [CrossRef]
- Chen, Y.; Zhu, S.; Hu, J.; Hu, Z.; Liu, X.; Wang, X.; Gu, M.; Hu, S.; Liu, X. gga-miR-1603 and gga-miR-1794 directly target viral L gene and function as a broad-spectrum antiviral factor against NDV replication. Virulence 2021, 12, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Tian, W.; Zhang, M.; Ren, T.; Sun, G.; Jiang, R.; Han, R.; Kang, X.; Yan, F. Transcriptom analysis revealed regulation of dexamethasone induced microRNAs in chicken thymus. J. Cell. Biochem. 2019, 120, 6570–6579. [Google Scholar] [CrossRef] [PubMed]
- Su, A.; Zhou, Y.; Guo, Y.; Yang, X.; Zhang, Y.; Li, W.; Tian, Y.; Li, K.; Sun, G.; Jiang, R. Identification and expression analysis of MicroRNAs in chicken spleen in a corticosterone-induced stress model. Res. Vet. Sci. 2021, 136, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, W.; Xu, H.; Liu, J.; Deng, Y.; Cheng, H.; Zhan, T.; Lu, X.; Liao, T.; Guo, L. Gga-miR-19b-3p inhibits newcastle disease virus replication by suppressing inflammatory response via targeting RNF11 and ZMYND11. Front. Microbiol. 2019, 10, 2006. [Google Scholar] [CrossRef]
- Zhou, C.; Tan, L.; Sun, Y.; Qiu, X.; Meng, C.; Liao, Y.; Song, C.; Liu, W.; Nair, V.; Ding, C. Exosomes carry microRNAs into neighboring cells to promote diffusive infection of Newcastle disease virus. Viruses 2019, 11, 527. [Google Scholar] [CrossRef]
- Kaufmann, L.; Syedbasha, M.; Vogt, D.; Hollenstein, Y.; Hartmann, J.; Linnik, J.E.; Egli, A. An optimized hemagglutination inhibition (HI) assay to quantify influenza-specific antibody titers. J. Vis. Exp. 2017, 130, e55833. [Google Scholar] [CrossRef]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Cellular and Molecular Immunology, 7th ed.; Saunders: Philadelphia, PA, USA, 2012. [Google Scholar]
- Cao, X. Medical Immunology, 6th ed.; People’s Medical Publishing House: Beijing, China, 2013. [Google Scholar]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar] [CrossRef]
- Tian, Y.; Wang, Q.; Han, J.; Wen, J.; Wu, Y.; Man, C. Stress-induced immunosuppression affecting avian influenza virus vaccine immune response through miR-20a-5p/NR4A3 pathway in chicken. Vet. Microbiol. 2022, 273, 109546. [Google Scholar] [CrossRef]
- Wu, Y.; Wen, J.; Han, J.; Tian, Y.; Man, C. Stress-induced immunosuppression increases levels of certain circulating miRNAs and affects the immune response to an infectious bursal disease virus vaccine in chickens. Res. Vet. Sci. 2022, 142, 141–148. [Google Scholar] [CrossRef]
- Cecchini, S.; Rossetti, M.; Di Tomaso, F.; Caputo, A.R. Evaluation of the effects of dexamethasone-induced stress on levels of natural antibodies in immunized laying hens. Vet. Immunol. Immunopathol. 2016, 177, 35–41. [Google Scholar] [CrossRef]
- Kong, Y.; Li, S.; Cheng, X.; Ren, H.; Zhang, B.; Ma, H.; Li, M.; Zhang, X.-A. Brain ischemia significantly alters microRNA expression in human peripheral blood natural killer cells. Front. Immunol. 2020, 11, 759. [Google Scholar] [CrossRef]
- Santangelo, L.; Bordoni, V.; Montaldo, C.; Cimini, E.; Zingoni, A.; Battistelli, C.; D’Offizi, G.; Capobianchi, M.R.; Santoni, A.; Tripodi, M. Hepatitis C virus direct-acting antivirals therapy impacts on extracellular vesicles microRNAs content and on their immunomodulating properties. Liver Int. 2018, 38, 1741–1750. [Google Scholar] [CrossRef]
- Rosenberger, C.M.; Podyminogin, R.L.; Navarro, G.; Zhao, G.-W.; Askovich, P.S.; Weiss, M.J.; Aderem, A. miR-451 regulates dendritic cell cytokine responses to influenza infection. J. Immunol. 2012, 189, 5965–5975. [Google Scholar] [CrossRef]
- Min, S.; Liang, X.; Zhang, M.; Zhang, Y.; Mei, S.; Liu, J.; Liu, J.; Su, X.; Cao, S.; Zhong, X. Multiple tumor-associated microRNAs modulate the survival and longevity of dendritic cells by targeting YWHAZ and Bcl2 signaling pathways. J. Immunol. 2013, 190, 2437–2446. [Google Scholar] [CrossRef]
- Lu, W.; You, R.; Yuan, X.; Yang, T.; Samuel, E.L.; Marcano, D.C.; Sikkema, W.K.; Tour, J.M.; Rodriguez, A.; Kheradmand, F. The microRNA miR-22 inhibits the histone deacetylase HDAC4 to promote TH17 cell–dependent emphysema. Nat. Immunol. 2015, 16, 1185–1194. [Google Scholar] [CrossRef]
- Liang, X.; Liu, Y.; Mei, S.; Zhang, M.; Xin, J.; Zhang, Y.; Yang, R. MicroRNA-22 impairs anti-tumor ability of dendritic cells by targeting p38. PLoS ONE 2015, 10, e0121510. [Google Scholar] [CrossRef]
- Fu, X.; Qie, J.; Fu, Q.; Chen, J.; Jin, Y.; Ding, Z. miR-20a-5p/TGFBR2 axis affects pro-inflammatory macrophages and aggravates liver fibrosis. Front. Oncol. 2020, 10, 107. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Liu, X.; Wang, W.; Cai, Y.; Li, S.; Chen, Q.; Liao, M.; Zhang, M.; Zeng, G.; Zhou, B. Down-regulation of miR-20a-5p triggers cell apoptosis to facilitate mycobacterial clearance through targeting JNK2 in human macrophages. Cell Cycle 2016, 15, 2527–2538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, W.; Chang, R.; Luo, Q.; Liu, G.; Xu, T. The long noncoding RNA MIR122HG is a precursor for miR-122-5p and negatively regulates the TAK1-induced innate immune response in teleost fish. J. Biol. Chem. 2022, 298, 101773. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.B.; Li, Z.-L.; Luo, D.H.; Huang, B.J.; Chen, Y.-S.; Zhang, X.S.; Cui, J.; Zeng, Y.X.; Li, J. Tumor-derived exosomes promote tumor progression and T-cell dysfunction through the regulation of enriched exosomal microRNAs in human nasopharyngeal carcinoma. Oncotarget 2014, 5, 5439. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, S.; Fan, M.; Ruan, G.; Xi, T.; Zheng, L.; Guo, L.; Ye, F.; Xing, Y. Helicobacter pylori induces gastric cancer via down-regulating miR-375 to inhibit dendritic cell maturation. Helicobacter 2021, 26, e12813. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Che, L.; Xu, C.; Lu, D.; Xu, Y.; Liu, M.; Chai, W. XIST/miR-34a-5p/PDL1 axis regulated the development of lung cancer cells and the immune function of CD8+ T cells. J. Recept. Signal. Transduct. 2022, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.N.; Ye, Y.X.; Niu, J.W.; Li, Y.; Li, X.; You, X.; Chen, H.; Zhao, L.D.; Zeng, X.F.; Zhang, F.C. Defective PTEN regulation contributes to B cell hyperresponsiveness in systemic lupus erythematosus. Sci. Transl. Med. 2014, 6, 246ra99. [Google Scholar] [CrossRef]
- Liu, W.-H.; Kang, S.G.; Huang, Z.; Wu, C.-J.; Jin, H.Y.; Maine, C.J.; Liu, Y.; Shepherd, J.; Sabouri-Ghomi, M.; Gonzalez-Martin, A. A miR-155–Peli1–c-Rel pathway controls the generation and function of T follicular helper cells. J. Exp. Med. 2016, 213, 1901–1919. [Google Scholar] [CrossRef]
- Du, X.; Zhu, M.; Zhang, T.; Wang, C.; Tao, J.; Yang, S.; Zhu, Y.; Zhao, W. The recombinant Eg. P29-mediated miR-126a-5p promotes the differentiation of mouse naive CD4+ T cells via DLK1-mediated notch1 signal pathway. Front. Immunol. 2022, 13, 773276. [Google Scholar] [CrossRef]
- Kamada, R.; Kudoh, F.; Ito, S.; Tani, I.; Janairo, J.I.B.; Omichinski, J.G.; Sakaguchi, K. Metal-dependent Ser/Thr protein phosphatase PPM family: Evolution, structures, diseases and inhibitors. Pharmacol. Ther. 2020, 215, 107622. [Google Scholar] [CrossRef]
- Schilling, S.H.; Datto, M.B.; Wang, X.-F. A phosphatase controls the fate of receptor-regulated Smads. Cell 2006, 125, 838–840. [Google Scholar] [CrossRef]
- Xiang, W.; Zhang, Q.; Lin, X.; Wu, S.; Zhou, Y.; Meng, F.; Fan, Y.; Shen, T.; Xiao, M.; Xia, Z. PPM1A silences cytosolic RNA sensing and antiviral defense through direct dephosphorylation of MAVS and TBK1. Sci. Adv. 2016, 2, e1501889. [Google Scholar] [CrossRef] [Green Version]
- Xin, Y.; Wang, D.; Huang, M.; Yu, J.; Fang, L.; Xiao, S. Proteome analysis of differential protein expression in porcine alveolar macrophages regulated by porcine reproductive and respiratory syndrome virus nsp1β protein. Virus Genes 2018, 54, 385–396. [Google Scholar] [CrossRef]
- Daams, R.; Massoumi, R. Nemo-like kinase in development and diseases: Insights from mouse studies. Int. J. Mol. Sci. 2020, 21, 9203. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-Z.; Shu, Q.-P.; Song, Y.; Zhang, H.-H.; Liu, Y.; Jin, B.-X.; Liuyu, T.-Z.; Li, C.; Huang, X.-C.; Du, R.-L. Phosphorylation of MAVS/VISA by Nemo-like kinase (NLK) for degradation regulates the antiviral innate immune response. Nat. Commun. 2019, 10, 3233. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Cao, Y.; Huang, J.; Tan, Y.; Wei, J.; Xiao, J.; Zou, J.; Feng, H. NLK suppresses MAVS-mediated signaling in black carp antiviral innate immunity. Dev. Comp. Immunol. 2021, 122, 104105. [Google Scholar] [CrossRef]
- Lv, Z.; Li, C.; Zhang, W.; Jin, C.; Shao, Y.; Xuemei, D.; Qingxi, H. Nemo like kinase negatively regulates NF-κB activation and coelomocytes apoptosis in Apostichopus japonicus. Dev. Comp. Immunol. 2016, 54, 109–115. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Primer (5′–3′) |
|---|---|
| miR-375 | RT-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTAACGCGA |
| F-ACACTCCAGCTGGGTTTGTTCGTTCGGCTC | |
| miR-20a-5p | RT-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCTACCTGC |
| F-ACACTCCAGCTGGGTAAAGTGCTTATAGTGC | |
| miR-451 | RT-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGAAACTCAG |
| F-ACACTCCAGCTGGGAAACCGTTACCATTACT | |
| miR-124a-3p | RT-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTGGCATTC |
| F-ACACTCCAGCTGGGTTAAGGCACGCGGTGA | |
| miR-199-5p | RT-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGAACAGGT |
| F-ACACTCCAGCTGGGCCCAGTGTTCAGACTAT | |
| miR-126-5p | RT-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCGCGTACC |
| F-ACACTCCAGCTGGGCATTATTACTTTTGG | |
| miR-19b-3p | RT-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTCAGTTTT |
| F-ACACTCCAGCTGGGTGTGCAAATCCATGCAA | |
| miR-200b-3p | RT-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGATCATCAT |
| F-ACACTCCAGCTGGGTAATACTGCCTGGTAAT | |
| miR-34a-5p | RT-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGAACAACCA |
| F-ACACTCCAGCTGGGTGGCAGTGTCTTAGCTG | |
| miR-122-5p | RT-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGACAAACAC |
| F-ACACTCCAGCTGGGTGGAGTGTGACAATGGT | |
| miR-22-3p | RT-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGACAGTTCT |
| F-ACACTCCAGCTGGGAAGCTGCCAGTTGAAG | |
| miR-31-5p | RT-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCAGCTATG |
| F-ACACTCCAGCTGGGAGGCAAGATGTTGGCA | |
| miR-30b-5p | RT-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGAGCTGAGT |
| F-ACACTCCAGCTGGGTGTAAACATCCTACAC | |
| miR-198 | RT-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGAACCTAT |
| F-ACACTCCAGCTGGGGGTCCAGAGGGGAGAT | |
| miR-29b-3p | RT-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGAACACTGA |
| F-ACACTCCAGCTGGGTAGCACCATTTGAAATC | |
| U6 | F-CTCGCTTCGGCAGCACA |
| R-AACGCTTCACGAATTTGCGT | |
| miRNA | R-TGGTGTCGTGGAGTCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Y.; Liu, Y.; Wang, Q.; Wen, J.; Wu, Y.; Han, J.; Man, C. Stress-Induced Immunosuppression Affects Immune Response to Newcastle Disease Virus Vaccine via Circulating miRNAs. Animals 2022, 12, 2376. https://doi.org/10.3390/ani12182376
Tian Y, Liu Y, Wang Q, Wen J, Wu Y, Han J, Man C. Stress-Induced Immunosuppression Affects Immune Response to Newcastle Disease Virus Vaccine via Circulating miRNAs. Animals. 2022; 12(18):2376. https://doi.org/10.3390/ani12182376
Chicago/Turabian StyleTian, Yufei, Yang Liu, Qiuyuan Wang, Jie Wen, Yiru Wu, Jianwei Han, and Chaolai Man. 2022. "Stress-Induced Immunosuppression Affects Immune Response to Newcastle Disease Virus Vaccine via Circulating miRNAs" Animals 12, no. 18: 2376. https://doi.org/10.3390/ani12182376
APA StyleTian, Y., Liu, Y., Wang, Q., Wen, J., Wu, Y., Han, J., & Man, C. (2022). Stress-Induced Immunosuppression Affects Immune Response to Newcastle Disease Virus Vaccine via Circulating miRNAs. Animals, 12(18), 2376. https://doi.org/10.3390/ani12182376