1. Introduction
Rotavirus (RV) is the main pathogen that induces severe diarrhea in infants, children, and young animals [
1,
2]. RV infection will damage the intestinal mucosa of the host, cause the disorder of nutrient absorption and water electrolytes, and finally lead to host diarrhea [
3]. As a double-stranded RNA virus, after entering the small intestine, RV will invade the epithelial cells and induce an inflammatory response, immune dysfunction, and oxidative stress, which can be the main causes of RV damaging gut barrier function and inducing diarrhea [
4,
5,
6,
7,
8]. The comprehensive infection rate of piglets (1–4 weeks old) is over 80%, and the mortality rate is as high as 20%. Moreover, infected breeding sows can become an invisible source of infection and further result in the continuous occurrence of RV infection in the local area [
9].
Lactose, known as a kind of carbohydrate that exists in mammal milk, is resolved into glucose and galactose by lactase. This can supply energy to infants, children, and young animals [
10]. In the large intestine, unabsorbed lactose may be utilized and fermented into short-chain fatty acids and lactic acid, and then the pH value of the enteric internal environment will be decreased, which will be beneficial for gut microflora [
11,
12,
13]. In particular, the activity of lactase in the gut will decrease with the increasing age of piglets. This increases the level of lactose in the large intestine, where its functions are mainly embodied via itself or short-chain fatty acids and lactic acid after fermentation by gut microflora. [
11,
12,
13]. Recently, research has reported that lactose administration could stimulate the generation of host defense peptides (HDPs) in human intestinal cells and phagocytes [
10]. Our previous study also showed that lactose treatment upregulated HDPs (such as porcine β-defensins (pBD), protegrin 1-5 (PG1-5)), mucins, tight-junction-related proteins, and antivirus cytokines (e.g., interferons (IFN)) in IPEC-J2 cells and/or 3D4/31 cells (data not published). Lactose has a potential role in maintaining gut health.
Based on the physiological functions of lactose, it is possible that dietary lactose supplementation could improve immunity and gut health and attenuate RV-induced diarrhea. Whey has already been used in the feed of young animals (especially piglets), the main component of which is lactose [
14]. According to the addition percentage of whey, generally, the content of lactose is approximately 4% in piglet feed. It is unknown whether increasing the content of lactose is more effective than 4% lactose. Therefore, the purpose of this research was to (1) test the hypothesis that supplementing different lactose levels in the diets relieves RV-induced diarrhea and improves growth performance and (2) analyze the potential change in gut health in this process.
2. Materials and Methods
2.1. Ethical Approval
The experimental protocol was approved by the Animal Care Advisory Committee of Sichuan Agricultural University (Chengdu 611130, China). All experimental protocols were approved by the Institutional Animal Care and Use Committee of the Laboratory Animal Center at Sichuan Agricultural University on 24 June 2019 (SICAU-2019-026). Animal testing was enforced at the Experimental Farm of Sichuan Agricultural University.
2.2. Animals and Diets
A total of 36 Duroc × Landrace × Yorkshire (DLY) 21-day-old weaned piglets (barrow) with similar initial body weight (BW, 6.30 ± 0.10 kg) from 12 litters were individually fed in metabolic cages (1.50 × 0.70 × 1.00 m3). All piglets were fed powders 4 times daily at 08:00, 12:00, 16:00, and 20:00 and received water ad libitum. The quantity of the feeding diet was based on the principle that there was a little remaining feed in the trough after feeding, and the remaining feed in the trough was cleaned, weighed, and recorded at 10:00 p.m. every day.
The basal diet was formulated to meet or exceed the nutrient recommendation for 7–11 kg pigs in NRC [
15], of which the composition and nutrient contents are exhibited in
Table 1. The experimental diets were formed by supplementing 0, 4%, and 6% lactose in the basal diet by replacing corn starch. The lactose was purchased from Baiying Biotechnology Co., Ltd., Jiangxi, China, with a purity of 99%.
2.3. Experimental Design and Sample Collection
After 3 days of orientation, on the basis of initial body weight and litter origin, all piglets were randomly allocated into 3 groups (
n = 12) and were fed with the experimental diets containing 0, 4%, and 6% lactose for 20 days. During Days 11–14, nutrient digestion was evaluated as described previously [
16].
RV preparation and virus titer measurement (tissue culture infective dose 50 (TCID
50) value) were carried out as described previously [
17].
On Day 15, all piglets were gavaged, and the specific gavage methods were carried out as described previously [
18]. Briefly, all piglets (39 days old) were administered 5 mL of sterile 100 mmol/L NaHCO
3 solution via oral gavage. After 15 min, half of the piglets in each group were orally infused with 5 mL (10
6 TCID
50/mL) of rotavirus (RV+), while the other piglets orally received 5 mL of sterile essential medium (RV−). Then, the diarrhea status of all piglets was observed and recorded each day. The scoring method of fecal consistency was carried out as described previously [
18]. In brief, normal, 0; pasty, 1; semiliquid, 2; liquid, 3. A fecal consistency score of ≥2 was considered diarrhea. Feed intake was recorded on each day of the experiment, and animals were weighed on Days 1, 15, and 21. The growth performance indexes (including average daily gain (ADG), average daily feed intake (ADFI), and feed/weight gain ratio (F/G)) of all piglets were calculated from the above-recorded data.
On Day 21, following 12 h of starvation, the piglets were weighed and infused with sterile 10 mg/mL D-xylose solution (1 mL/kg of body weight). The D-xylose was purchased from Haoboyou Biotechnology Co., Ltd., Chengdu, China, with a purity of 99%. One hour later, the blood sample of all piglets was gathered from the jugular vein, centrifuged at 3500× g for 10 min, and serum was collected. Then, after refeeding for 2 h, all piglets were euthanized via intracardiac injection with sodium pentobarbital (50 mg/kg of body weight) and exsanguinated. The whole intestine was removed. The jejunum was quickly separated and flushed with ice-cold saline solution. Lymph nodes were immediately excised. The sample of jejunal mucosa was collected by scraping the gut wall with the glass microscope slide. About 2 cm jejunal segments were fixed in 4% paraformaldehyde. About 3 g of cecal digesta was gathered in sterile tubes, and the other cecal digesta were used to measure the pH value using a pH meter (PHS-3C). The samples of lymph nodes, jejunal mucosa, and cecal digesta were immediately frozen in liquid nitrogen and stored at −80 °C.
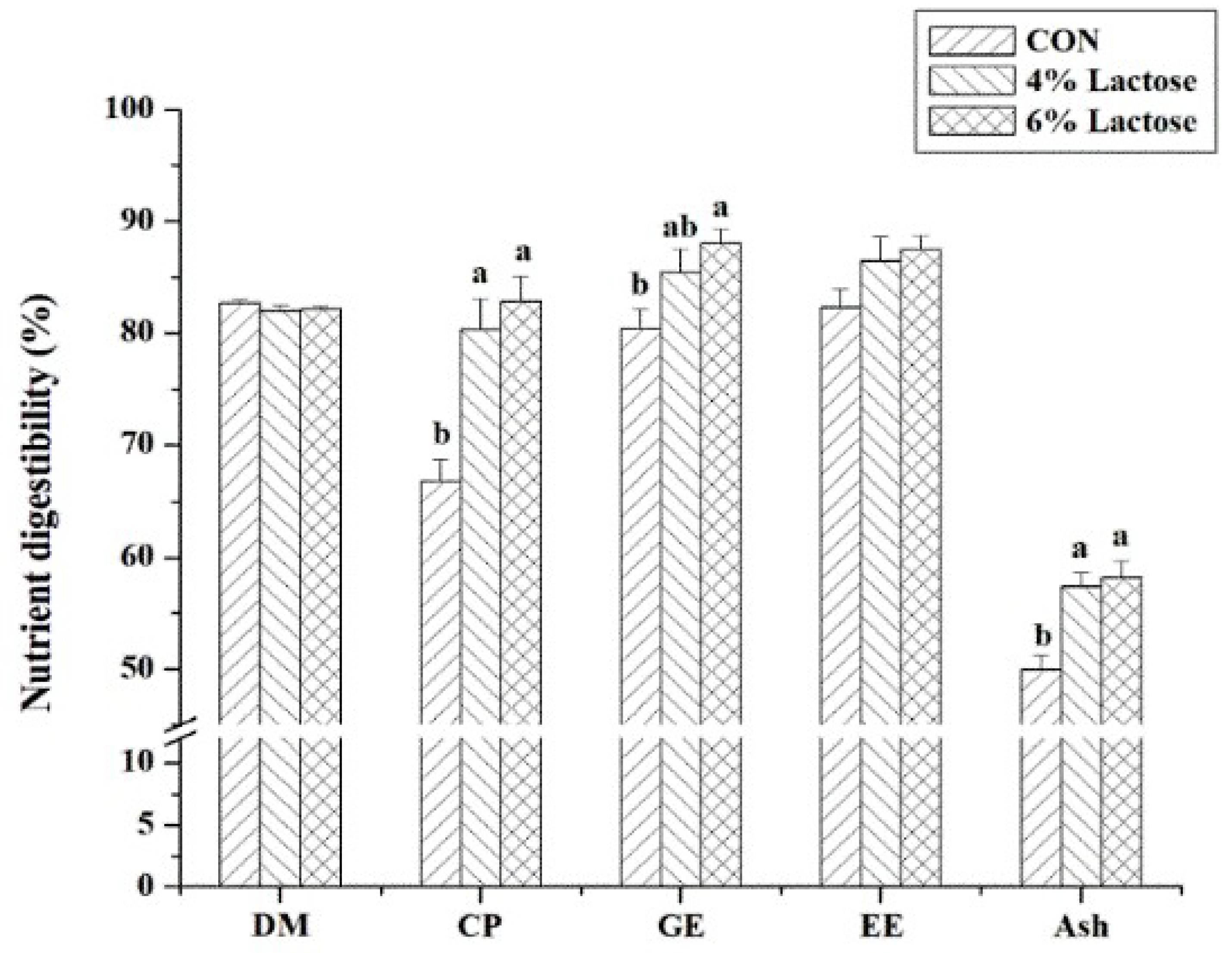
2.4. Nutrient Digestibility
In the digestion trial, ash insoluble in hydrochloric acid was the nonabsorbable digestion marker. Dry matter (DM), crude protein (CP), gross energy (GE), ether extract (EE), ash, and ash insoluble in hydrochloric acid in feeds and feces were determined according to the previous description [
16]. The digestibility of these nutrients was calculated as previously described [
18].
2.5. Serum Urea Nitrogen (UN) and D-Xylose Levels
Serum UN (C013-2-1) and D-xylose (A035-1-1) concentrations were analyzed by using the kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer’s instructions.
2.6. Rotavirus Nonstructural Protein 4 (NSP4), Secreting Immunoglobulin A (sIgA), and Antioxidant Capacity in Jejunal Mucosa
Approximately 100 mg of jejunal mucosa was added into ice-cold PBS, shattered at 4 °C, and then centrifuged at 5000× g for 15 min at 4 °C. The supernatants were collected and were used to measure the related index.
The NSP4 concentration in jejunal mucosa was measured with an ELISA kit (TSZ ELISA, Framingham, MA, USA). The sIgA concentration in jejunal mucosa was detected with ELISA kits from Nuoyuan Co. Ltd. (Shanghai, China). Malondialdehyde (MDA) level, total antioxidant capacity (T-AOC), and total protein level from jejunal mucosa were detected by the kit of Nanjing Jiancheng Bioengineering Institute (Nanjing, China).
2.7. Morphology of Jejunal Mucosa
The morphology in jejunal mucosa was determined as described previously [
4]. In brief, following fixing in 4% paraformaldehyde, the jejunal segment was embedded in paraffin. Then, consecutive sections (5 µm) were stained with hematoxylin–eosin. In each sample, a total of 10 intact villi and crypts were randomly selected, and villus height and crypt depth were measured at 40× magnification with an Olympus CK 40 microscope.
2.8. mRNA Expression of Some Genes in Jejunal Mucosa and/or Mesenteric Lymph Nodes
Total RNA in jejunal mucosa and mesenteric lymph nodes was extracted with TRIZOL reagent (TaKaRa Biotechnology (Dalian) Co., Ltd., Dalian, China) according to the manufacturer’s instructions. RNA concentrations were determined with DU 640 UV spectrophotometer detection (Beckman Coulter Inc., Fullerton, CA, USA). The OD
260/OD
280 ratio was 1.8 to 2.0. RNA quality of samples was evaluated with 1% agarose gel electrophoresis. Then, in all samples, the cDNA was synthesized by using RT Reagents (TaKaRa Biotechnology (Dalian) Co., Ltd., Dalian, China) according to the manufacturer’s instructions. All genes’ primers of this study are listed in
Table 2 and obtained from TaKaRa Biotechnology (Dalian) Co., Ltd. (Dalian, China). The mRNA expression of mucin 1 (MUC1), MUC2, zonula occludens-1 (ZO-1), occludin, B-cell lymphoma/leukemia-2-associated X protein (Bax), B-cell lymphoma/leukemia-2 (Bcl-2), interleukin 2 (IL-2), IL-4, interferon γ (IFN-γ), IFN-β, porcine β-defensin 1 (pBD1), pBD2, pBD3, and β-actin in jejunal mucosa and/or mesenteric lymph nodes was determined by real-time quantitative PCR with SYBR Premix Ex Taq reagents (TaKaRa Biotechnology (Dalian) Co., Ltd., Dalian, China) and CFX-96 Real-Time PCR Detection System (Bio-Rad Laboratories, Richmond, CA, USA) as described previously [
18]. To verify the variance in the amount of RNA input of reaction, β-actin was utilized as the reference gene. Then, the relative mRNA expression was obtained by using the previous method [
19].
2.9. Populations of Some Bacteria in the Cecal Digesta
Bacterial DNA of cecal digesta was extracted with the Stool DNA Kit (Omega BioTek, Doraville, CA, USA) according to the manufacturer’s instructions. The bacterial real-time quantitative PCR and related result transformation were executed as described previously [
4]. All primers and probes, listed in
Table 3, were purchased from TaKaRa Biotechnology (Dalian) Co., Ltd. (Dalian, China).
2.10. Short-Chain Fatty Acids (SCFAs) in the Cecal Digesta
SCFAs (including acetic acid, propionic acid, isobutyric acid, butyric acid, isovaleric acid, and valeric acid) levels were analyzed with the Varian CP-3800 gas chromatograph (Agilent Technologies, Santa Clara, CA, USA) as described previously [
4].
2.11. Statistical Analysis
The index of growth and nutrient digestibility of piglets before RV challenge was analyzed by using one-way ANOVA, followed by Duncan’s multiple range test. The data on diarrhea rate of piglets were analyzed through the chi-square test. The other data were analyzed as a 2 × 3 factorial with the general linear model procedures of the Statistical Analysis Package. The model factors were involved in dietary lactose supplementation, rotavirus challenge, and their interaction. These analyses were executed with SPSS (Version 21.0, IBM, Armonk, NY, USA). All data were embodied as means and their SEs. p < 0.05 was considered statistically significant, whereas p < 0.10 was considered a statistical tendency.
4. Discussion
Pathogen-induced diarrhea is the main cause that negatively affects health and/or growth in humans and animals, especially infants, children, and young animals [
3]. Rotavirus (RV) is one of the major pathogens that induce severe diarrhea in young humans and animals by damaging gut health [
1,
2,
20]. Our previous studies have reported that RV infection causes diarrhea, a decrease in growth performance, dysfunction of the gut barrier, inflammation, and changes in the gut bacteria of weaned pigs [
4,
5,
6,
17,
18,
20,
21]), similar to the results of this study. Besides these, we also found that RV challenge increased RV NSP4 levels of jejunal mucosa and decreased serum D-xylose consistency in weaned piglets. Thus, these results illustrated that RV infection in weaned piglets was successfully implemented.
Previous research has shown that supplementing lactose in diets might increase the growth performance of weaned piglets and nursery pigs, and the influence of lactose administration on growth performance is dose-dependent [
22,
23,
24]. The present research achieved similar results, showing that supplementing lactose in the diets improved the F/G of weaned piglets and tended to improve the ADG of weaned piglets and the F/G of RV-infected piglets. These results also embodied the better effect of high-dose lactose. It is well known that the digestion, absorption, and utilization of nutrients play a vital role in the growth of humans and animals. In this research, nutrient digestibility and serum UN in weaned piglets were measured, and we discovered that lactose administration raised the digestibility of CP, GE, and ash and decreased serum UN, known as a potential marker of nitrogen (especially protein and amino acid) utilization, in weaned piglets. These illustrated that supplementing lactose in the diets improving the growth performance of piglets was associated with an increase in nutrient digestion, absorption, and utilization. Moreover, our results showed that lactose administration increasing growth performance on Days 1 to 14 was more efficient than that on Days 15 to 20. Therefore, lactose should be supplemented in the feed of piglets during the initial stage of weaning.
Weaning is often coupled with intestinal morphology damage, which has been widely applied to evaluate the intestinal health or functions of piglets [
25]. Gut epithelial integrity is one of the important causes that maintain digestion and absorption functions, and the surface area of villi in the intestinal mucosa is related to nutrient digestion and absorption. In the current research, lactose administration did not affect villus height and crypt depth but, to some extent, improved villus height/crypt depth in the jejunal mucosa of normal and/or RV-infected piglets. Despite this, we also found that dietary lactose supplementation could increase serum D-xylose levels in piglets, which could reflect the positive effect of lactose on gut absorption function and epithelial integrity.
NSP4 is a nonstructural protein of RV, which is a vital factor that infects gut epithelial cells and induces severe diarrhea in humans and animals. NSP4 is often regarded as an important marker of RV infection [
26]. The results of our research showed that RV challenge elevated NSP4 concentrations, but lactose administration decreased NSP4 concentrations in the jejunal mucosa of weaned piglets. Thus, it is possible that supplementing lactose in the diets inhibited RV invasion and infection in the intestine of piglets.
Intestinal barrier function is vital for protecting gut health from pathogen invasion [
27]. Epithelial tight junctions (TJs) are a critical physical barrier against the permeation of many molecules (such as pathogens, toxins, and antigens) from the luminal environment into the mucosal tissues and circulatory system [
28]. Lactose administration potentially improved the intestinal barrier function in RV-infected piglets. Thus, except for mucosal epithelial integrity (such as mucosal morphology and serum D-xylose levels), we analyzed the gut-barrier-related indexes. The intercellular junctions between epithelial cells of the mucosa are mainly associated with transmembrane and no membrane proteins, such as ZO-1 and occludin [
29]. The main component of the mucus gel layer is mucins generated by the goblet and epithelial cells of the mucosa [
30]. In addition, immunity is also pivotal to preventing pathogen (especially virus) invasion in the gut, which includes some immune macromolecules, including sIgA, interferons, and host-defense peptides [
31,
32,
33]. In our research, supplementing lactose in the diets alleviated the influence of RV infection on sIgA level and MUC1, MUC2, ZO-1, occludin, pBD1, and pBD3 mRNA expression of the jejunal mucosa, and further promoted IFN-γ and IFN-β mRNA expression of jejunal mucosa and/or mesenteric lymph nodes in weaned piglets. These illustrated that supplementing lactose in the diets protecting the gut from RV infection was related to the improvement of barrier function.
In the process of maintaining gut barrier function, the activity of mucosal cells plays an important role. Bax and Bcl-2 are apoptosis-related proteins [
34]. The results of our study showed that supplementing lactose in the diets relieved the negative influence of RV infection on the mRNA expression of Bax and Bcl-2 in the jejunal mucosa of weaned piglets. Impairing intracellular redox status and increasing inflammation are two important ways to increase RV-induced apoptosis of gut mucosal cells (especially in the late period of RV infection) [
5]. In this study, supplementing lactose in the diets did not regulate the decreasing antioxidant capacity of the jejunal mucosa, but alleviated the increasing proinflammatory cytokine (IL-2) and decreasing anti-inflammatory cytokine (IL-4) mRNA expression of jejunal mucosa and/or mesenteric lymph nodes in RV-infected piglets. Thus, lactose administration improving the gut barrier function of RV-infected piglets could result from the regulated apoptosis of mucosal cells possibly via the downregulated inflammation.
Gut microbiota and their metabolites (including SCFAs) also play a critical role in gut barrier function [
35,
36]. Previous researches have reported that lactose administration can increase the number of beneficial bacteria in the intestine and SCFA levels in humans and animals [
11,
12,
13], which is similar to the results of our research. It was possible that supplementing high-dose lactose in the diets attenuating diarrhea induced by RV infection, at least partially, should be in contact with the improvement of gut microbiota in piglets.
In addition, we also found that high-dose lactose administration had a more significant effect on improving growth performance (especially in the first two weeks) and reducing RV-induced diarrhea. After analyzing all indexes, this could be due to the fact that the improvement of nutrient digestibility (such as GE) and gut barrier functions (including the mRNA expression of mucins, tight junctions, and immunity) in piglets with a diet supplemented with 6% lactose is more efficient than that in piglets with a diet supplemented with 4% lactose.

 ,
, 



{kind=link}