
Frequency of β-Casein Gene Polymorphisms in Jersey Cows in Western Japan


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
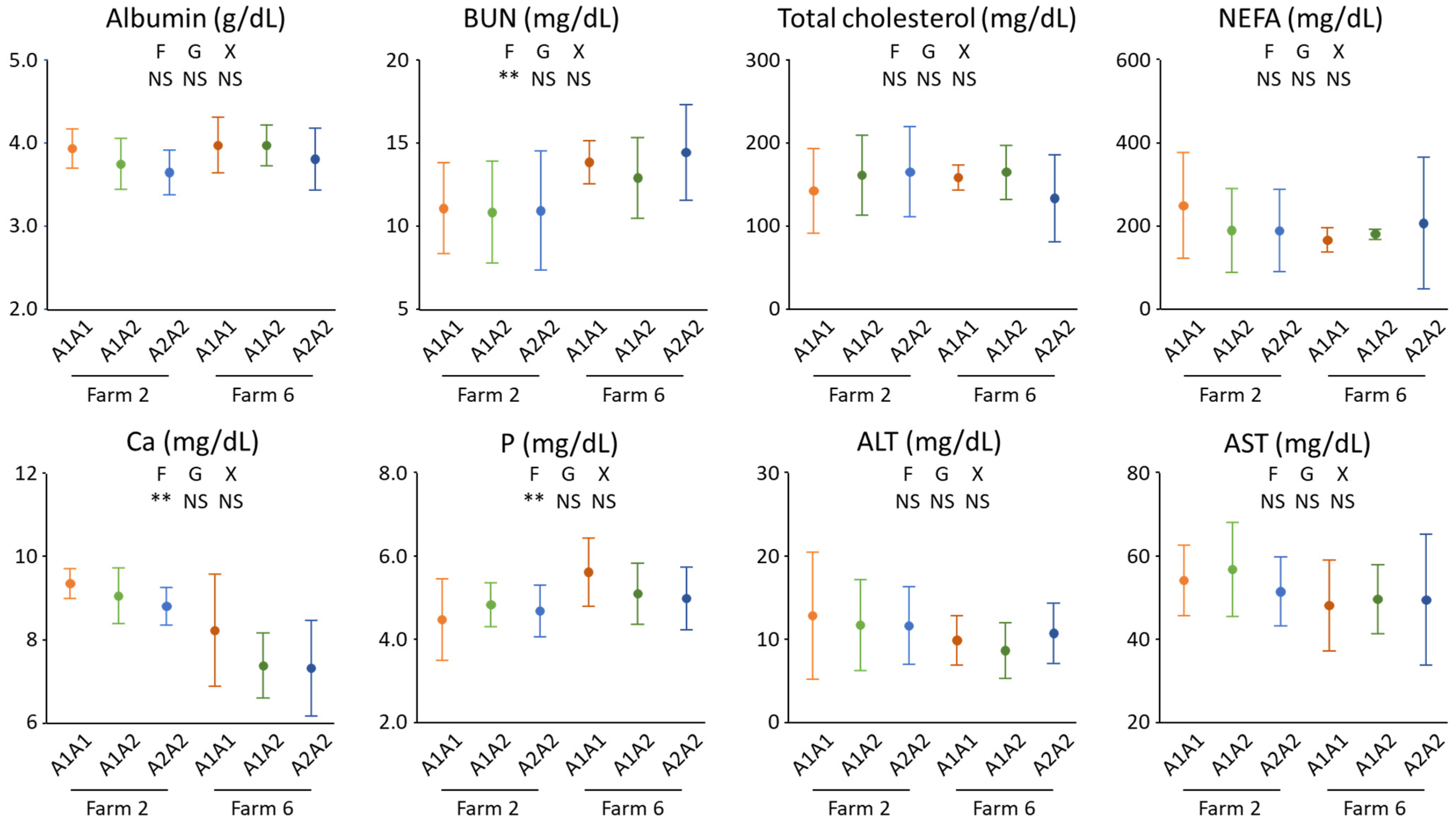
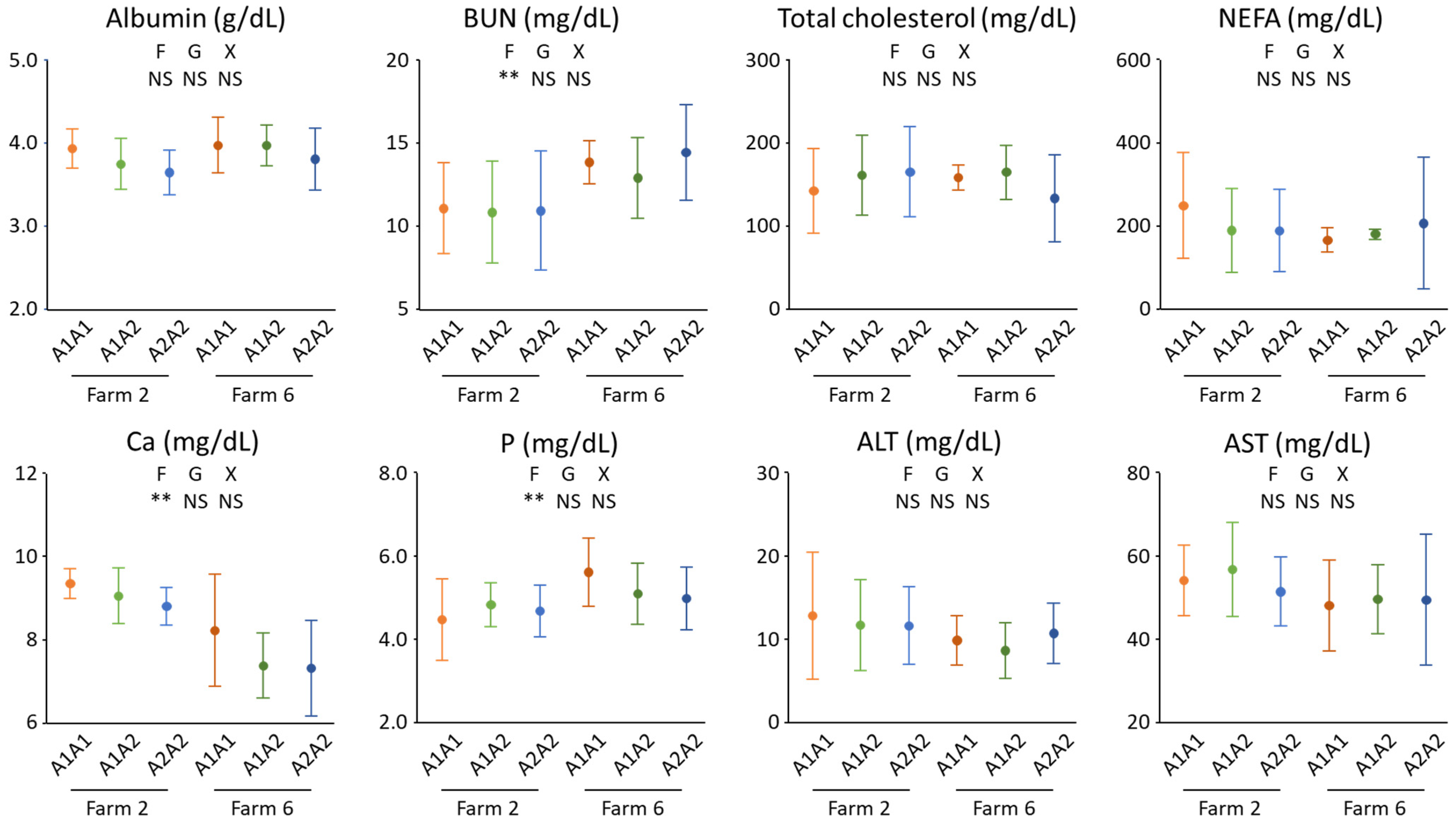
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaminski, S.; Cieslinska, A.; Kostyra, E. Polymorphism of bovine beta-casein and its potential effect on human health. J. Appl. Genet. 2007, 48, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Elferink, A.J.W.; Entiriwaa, D.; Bulgarelli, P.; Smits, N.G.E.; Peters, J. Development of a microsphere-based immunoassay authenticating A2 milk and species purity in the milk production chain. Molecules 2022, 27, 3199. [Google Scholar] [CrossRef] [PubMed]
- Sebastiani, C.; Argangeli, C.; Ciullo, M.; Torricelli, M.; Cinti, G.; Fisichella, S.; Biagetti, M. Frequencies evaluation of β-casein gene polymorphisms in dairy cows reared in central Italy. Animals 2020, 10, 252. [Google Scholar] [CrossRef] [PubMed]
- Caroli, A.M.; Savino, S.; Bulgari, O.; Monti, E. Detecting β-casein gene variation in bovine milk. Molecules 2016, 21, 141. [Google Scholar] [CrossRef] [PubMed]
- de Gaudry, D.K.; Lohner, S.; Schmucker, C.; Kapp, P.; Motschall, E.; Horrlein, S.; Roger, C.; Meerpohl, J.J. Milk A1 β-casein and health-related outcomes in humans: A systematic review. Nutr. Rev. 2019, 77, 278–306. [Google Scholar] [CrossRef] [PubMed]
- de Gaudry, D.K.; Lohner, S.; Bischoff, K.; Schmucker, C.; Hoerrlein, S.; Roeger, C.; Schwingshackl, L.; Meerpohl, J.J. A1 and A2 beta-casein on health-related outcomes: A scoping review of animal studies. Eur. J. Nutr. 2022, 61, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Yamada, A.; Sugimura, M.; Kuramoto, T. Genetic polymorphism of bovine beta-casein gene in Japanese dairy farm herds. Anim. Sci. J. 2021, 92, e13644. [Google Scholar] [CrossRef] [PubMed]
- Gustavsson, F.; Buitenhuis, A.J.; Johansson, M.; Bertelsen, H.P.; Glantz, M.; Poulsen, A.; Lindmark Mansson, H.; Stalhammar, H.; Larsen, L.B.; Bendixen, C.; et al. Effects of breed and casein genetic variants on protein profile in milk from Swedish Red, Danish Holstein, and Danish Jersey cows. J. Dairy Sci. 2014, 97, 3866–3877. [Google Scholar] [CrossRef] [PubMed]
- Dai, R.; Fang, Y.; Zhao, W.; Liu, S.; Ding, J.; Xu, K.; Yang, L.; He, C.; Ding, F.; Meng, H. Identification of alleles and genotypes of beta-casein with DNA sequencing analysis in Chinese-Holstein cow. J. Dairy Res. 2016, 83, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Zepeda-Batista, J.L.; Alarcon-Zuniga, B.; Ruiz-Flores, A.; Nunez-Dominguez, R.; Ramirez-Valverde, R. Polymorphism of three milk protein genes in Mexican Jersey cattle. Electron. J. Biotechnol. 2015, 18, 1–4. [Google Scholar] [CrossRef]
- Antonopoulos, D.; Vougiouklaki, D.; Laliotis, G.P.; Tsironi, T.; Valasi, I.; Chatzilazarou, A.; Halvatsiotis, P.; Houhoula, D. Identification of polymorphisms of the CSN2 gene encoding beta-casein in Greek local breeds of cattle. Vet. Sci. 2021, 8, 257. [Google Scholar] [CrossRef] [PubMed]
- Ivankovic, A.; Pecina, M.; Ramljak, J.; Pasic, V. Genetic polymorphism and effect on milk production of CSN2 gene in conventional and local cattle breeds in Croatia. J. Dairy Prod. Process Improv. 2021, 71, 3–12. [Google Scholar] [CrossRef]
- Massella, E.; Piva, S.; Giacometti, F.; Liuzzo, G.; Zambrini, A.V.; Serraino, A. Evaluation of bovine beta casein polymorphism in two dairy farms located in northern Italy. Ital. J. Food Saf. 2017, 6, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Mohan, G.; Kumar, A.; Khan, S.H.; Kumar, N.A.; Kapila, S.; Lathwal, S.S.; Sodhi, M.; Niranjan, S.K. Casein (CSN) gene variants and parity affect the milk protein traits in crossbred (Bos Taurus X Bos indicus) cows in sub-tropical climate. Trop. Anim. Health Prod. 2021, 53, 289. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, R.V.; Chauhan, A.; Ilayakumar, K.; Kumar, S.; Kumar, A.; Sonwane, A.; Kumar, S.; Panigrahi, M.; Bhushan, B. Genetic association analysis reveals significant effect of β-casein A1/A2 loci on production and reproduction traits in Frieswal crossbred cows. Biol. Rhythm Res. 2020, 51, 1259–1272. [Google Scholar] [CrossRef]
- Molee, A.; Boonek, L.; Rungsakinnin, N. The effect of beta and kappa casein genes on milk yield and milk composition in different percentages of Holstein in crossbred dairy cattle. Anim. Sci. J. 2011, 82, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Rangel, A.H.N.; Zaros, L.G.; Lima, T.C.; Borba, L.H.F.; Novaes, L.P.; Mota, L.F.M.; Silva, M.S. Polymorphism in the beta casein gene and analysis of milk characteristics in Gir and Guzera dairy cattle. Genet. Mol. Res. 2017, 16, gmr16029592. [Google Scholar] [CrossRef] [PubMed]
- Cieslinska, A.; Fiedorowicz, E.; Zwiezchowski, G.; Kordulewska, N.; Jarmolowska, B.; Kostyra, E. Genetic polymorphism of β-casein gene in Polish Red cattle-Preliminary study of A1 and A2 frequency in genetic conservation herd. Animals 2019, 9, 377. [Google Scholar] [CrossRef] [PubMed]
- Olenski, K.; Kaminski, S.; Szyda, J.; Cieslinska, A. Polymorphism of the beta-casein gene and its associations with breeding value for production traits of Holstein-Friesian bulls. Live Sci. 2010, 131, 137–140. [Google Scholar] [CrossRef]
- Visker, M.H.P.W.; Dibbits, B.W.; Kinders, S.M.; van Valenberg, H.J.F.; van Arendonk, J.A.M.; Bovenhuis, H. Association of bovine β-casein protein variant I with milk production and milk protein composition. Anim. Genet. 2010, 42, 212–218. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Variant | Exon 6 | Exon 7 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 18 | 25 | 35 | 36 | 37 | 67 | 72 | 88 | 93 | 106 | 122 | 138 | |
| A1 | His | |||||||||||
| A2 | Ser-P | Arg | Ser-P | Glu | Glu | Pro | Gln | Leu | Met | His | Ser | Pro |
| A3 | Gln | |||||||||||
| B | His | Arg | ||||||||||
| C | Ser | Lys | His | |||||||||
| D | Lys | |||||||||||
| E | Lys | |||||||||||
| F | His | Leu | ||||||||||
| G | His | Leu | ||||||||||
| H1 | Cys | Ileu | Glu | |||||||||
| H2 | Glu | Leu | ||||||||||
| I | Leu | |||||||||||
| Date of Blood Sampling | Yield (kg/day/cow) | Protein (%) | Fat (%) | SNF (%) | Lactose (%) | SCC (X103 cells/mL) | MUN (mg/mL) | |
|---|---|---|---|---|---|---|---|---|
| F1 (115) | 29 October 2019 | 21.6 ± 0.42 | 4.07 ± 0.12 | 5.06 ± 0.23 | 9.54 ± 0.07 | 4.55 ± 0.05 | 205 ± 30.5 | 8.14 ± 0.88 |
| F2 (82) | 24 November 2020 | 23.6 ± 0.72 | 3.98 ± 0.18 | 5.20 ± 0.20 | 9.42 ± 0.16 | 4.51 ± 0.02 | 115 ± 22.0 | 7.59 ± 1.04 |
| F3 (39) | 20 October 2020 | 16.0 ± 0.51 | 4.17 ± 0.20 | 4.96 ± 0.29 | 9.59 ± 0.21 | 4.45 ± 0.02 | 351 ± 104 | 11.9 ± 1.31 |
| F4 (98) | 25 November 2020 | 23.7 ± 0.65 | 4.02 ± 0.13 | 5.01 ± 0.20 | 9.49 ± 0.14 | 4.55 ± 0.03 | 148 ± 62.6 | 8.90 ± 1.12 |
| F5 (43) | 19 October 2020 | 18.9 ± 0.67 | 4.10 ± 0.09 | 4.95 ± 0.25 | 9.48 ± 0.07 | 4.45 ± 0.01 | 265 ± 22.0 | 11.1 ± 0.47 |
| F6 (60) | 24 November 2020 | 21.7 ± 0.98 | 4.19 ± 0.11 | 5.27 ± 0.20 | 9.68 ± 0.12 | 4.54 ± 0.03 | 172 ± 22.9 | 9.60 ± 0.83 |
| F7 (94) | 11 November 2020 | 24.4 ± 0.75 | 3.96 ± 0.17 | 4.98 ± 0.11 | 9.40 ± 0.15 | 4.51 ± 0.03 | 207 ± 17.6 | 9.86 ± 1.34 |
| F8 (59) | 25 November 2020 | 18.8 ± 1.35 | 4.03 ± 0.14 | 4.66 ± 0.23 | 9.38 ± 0.17 | 4.43 ± 0.06 | 327 ± 43.6 | 10.9 ± 2.61 |
| Genotype Frequency | Allele Frequency | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A1A1 | A1B | BB | A1A2 | A1I | A2B | BI | A2A2 | A2I | II | A1 | A2 | B | I | |
| F1 (115) | 0.009 | 0.017 | 0 | 0.069 | 0.009 | 0.243 | 0.061 | 0.496 | 0.096 | 0 | 0.057 | 0.700 | 0.161 | 0.082 |
| F2 (82) | 0 | 0.061 | 0.012 | 0.098 | 0 | 0.244 | 0.000 | 0.512 | 0.073 | 0 | 0.079 | 0.719 | 0.165 | 0.037 |
| F3 (39) | 0 | 0 | 0 | 0.077 | 0 | 0.102 | 0.026 | 0.641 | 0.154 | 0 | 0.038 | 0.808 | 0.064 | 0.090 |
| F4 (98) | 0 | 0 | 0 | 0.092 | 0 | 0.092 | 0.020 | 0.704 | 0.092 | 0 | 0.046 | 0.842 | 0.056 | 0.056 |
| F5 (43) | 0.023 | 0.023 | 0.023 | 0.070 | 0.023 | 0.140 | 0.023 | 0.442 | 0.210 | 0.023 | 0.082 | 0.651 | 0.116 | 0.151 |
| F6 (60) | 0.017 | 0.067 | 0.017 | 0.116 | 0 | 0.266 | 0.017 | 0.483 | 0.017 | 0 | 0.108 | 0.683 | 0.192 | 0.017 |
| F7 (94) | 0 | 0.011 | 0 | 0.074 | 0 | 0.160 | 0.021 | 0.606 | 0.128 | 0 | 0.043 | 0.787 | 0.096 | 0.074 |
| F8 (59) | 0 | 0 | 0 | 0.068 | 0 | 0.237 | 0.034 | 0.525 | 0.119 | 0.017 | 0.034 | 0.737 | 0.136 | 0.093 |
| Total (590) | 0.005 | 0.022 | 0.005 | 0.083 | 0.003 | 0.190 | 0.027 | 0.558 | 0.104 | 0.003 | 0.059 | 0.746 | 0.125 | 0.070 |
| References | Year | Country | Breed | Genotype Frequency | Allele Frequency | |||
|---|---|---|---|---|---|---|---|---|
| A1A1 | A1A2 | A2A2 | A1 | A2 | ||||
| This study | 2022 | Japan | Jersey (590) | 0.032 | 0.304 | 0.664 | 0.184 | 0.816 |
| Antonopoulos et al. | 2021 | Greece | Holstein (780) | 0.033 | 0.445 | 0.522 | 0.256 | 0.744 |
| Vrachykeratiki (46) | 0.000 | 0.609 | 0.391 | 0.304 | 0.696 | |||
| Ivankovic et al. | 2021 | Croatia | Simmental (60) | 0.033 | 0.417 | 0.550 | 0.242 | 0.758 |
| Holstein (60) | 0.134 | 0.433 | 0.433 | 0.350 | 0.650 | |||
| Brown Swiss (60) | 0.067 | 0.517 | 0.416 | 0.325 | 0.675 | |||
| Mohan et al. | 2021 | India | Karan Fries (Holstein Friesian × Tharparkar crossbreed) (100) | 0.090 | 0.620 | 0.290 | 0.400 | 0.600 |
| Rangel et al. | 2017 | Brazil | Gir (68) | 0.000 | 0.044 | 0.956 | 0.022 | 0.978 |
| Guzera (88) | 0.000 | 0.068 | 0.932 | 0.034 | 0.966 | |||
| Yamada et al. | 2021 | Japan | Holstein (311) & other three breeds (9) | 0.141 | 0.441 | 0.418 | 0.361 | 0.639 |
| Sebastiani et al. | 2020 | Italy (central) | Holstein Friesian (1629) | 0.132 | 0.458 | 0.410 | 0.361 | 0.639 |
| Kumar et al. | 2020 | India | Frieswal (Friesian × Sahiwal crossbreed) (429) | 0.175 | 0.515 | 0.310 | 0.432 | 0.568 |
| Cieslinska et al. | 2019 | Poland | Polish Red (177) | 0.367 | 0.526 | 0.107 | 0.630 | 0.370 |
| Massella et al. | 2017 | Italy (northern) | Holstein Friesian (1226) & Braunvieh (4) | 0.185 | 0.487 | 0.328 | 0.428 | 0.572 |
| Dai et al. | 2016 | China | Holstein (133) | 0.271 | 0.444 | 0.285 | 0.492 | 0.508 |
| Zepeda-Batista et al. | 2015 | Mexico | Jersey (453) | 0.080 | 0.364 | 0.556 | 0.262 | 0.738 |
| Gustavsson et al. | 2014 | Sweden | Holstein (415) | 0.089 | 0.446 | 0.465 | 0.312 | 0.688 |
| Swedish Red (392) | 0.230 | 0.525 | 0.245 | 0.492 | 0.508 | |||
| Jersey (406) | 0.108 | 0.382 | 0.510 | 0.299 | 0.701 | |||
| Molee et al. | 2011 | Thailand | Holstein × Bos indicus crossbreed (231) | 0.074 | 0.640 | 0.286 | 0.394 | 0.606 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nuomin; Nguyen, Q.D.; Aodaohu; Nishino, N. Frequency of β-Casein Gene Polymorphisms in Jersey Cows in Western Japan. Animals 2022, 12, 2076. https://doi.org/10.3390/ani12162076
Nuomin, Nguyen QD, Aodaohu, Nishino N. Frequency of β-Casein Gene Polymorphisms in Jersey Cows in Western Japan. Animals. 2022; 12(16):2076. https://doi.org/10.3390/ani12162076
Chicago/Turabian StyleNuomin, Qui Dang Nguyen, Aodaohu, and Naoki Nishino. 2022. "Frequency of β-Casein Gene Polymorphisms in Jersey Cows in Western Japan" Animals 12, no. 16: 2076. https://doi.org/10.3390/ani12162076
APA StyleNuomin, Nguyen, Q. D., Aodaohu, & Nishino, N. (2022). Frequency of β-Casein Gene Polymorphisms in Jersey Cows in Western Japan. Animals, 12(16), 2076. https://doi.org/10.3390/ani12162076