
Post-Mortem Dental Profile as a Powerful Tool in Animal Forensic Investigations—A Review


Abstract
Simple Summary
Abstract
1. Animals in Forensic Sciences
- Assessment of animal welfare (includes survival-related factors: nutrition, environment, health; situation-related factor: behaviour; emotional-related factor: mental state). Includes giving an opinion as to whether an animal may be experiencing or has experienced pain, discomfort, or distress in the past. In some instances, this evaluation may incorporate experts from other scientific disciplines, such as nutritionists and animal behaviourists.
- Determination of the time, the cause, and the circumstances of death of an animal, as well as other related investigations, such as the identification and interpretation of changes in the different body tissues, recognition of parasites and detection of signs of poisoning.
- Verification of the history and provenance of live/dead animals and animal derivatives. Most of the cases are related to domestic species; however, other cases fall under the category of ‘wildlife crime’. In this latter case, this field is related to whether national or international conservation legislation has been breached, such as the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES). This is a large field that includes offences and trapping of wild animals for sale, malicious poisoning, poaching, and the importation of endangered species or derivatives thereof.
- Performing clinical or post-mortem examinations when animal abuse appears to be related to acts of violence towards humans.
- Food safety. Ranges from animal welfare during slaughter (including religious slaughter) to meat inspection standards and the detection of contaminants, import, export, and correct identification of food.
- Human welfare. Covers public health issues such as zoonotic infections, health and safety (both in the workplace and in care institutions), and personal injury cases (e.g., kicks, bites, venomous bites, scratches, etc.).
- Miscellaneous. Miscellaneous cases relating to legal responsibility, negligence, nuisance, fraud, environmental pollution, damage to Crown property (e.g., the swans of Queen Elizabeth II in the UK), as well as other miscellaneous legislation and situations.
1.1. The Animal as the Victim
- To control an animal. Excessive and cruel physical punishment may be employed to exert control or shape an animal’s undesirable behaviours.
- To retaliate against an animal. Extreme punishment or retaliation may be inflicted for suspected misbehaviour on the part of an animal.
- To satisfy a prejudice against a species or breed. On numerous occasions, people designate certain groups of animals as good or bad. These beliefs may be related to cultural values, such as prejudice in our society against spiders or snakes.
- To express aggression through an animal. Inculcating violent tendencies in the animal in order to express violent, aggressive behaviours toward other people or animals.
- To strengthen one’s own aggressiveness. Killing and abusing animals may be a way to enhance one’s aggressive aptitudes or to impress others with the capacity for violence.
- To impress people for amusement. Cruelty toward animals sometimes occurs as a means of creating amusement and ‘entertaining’ friends.
- To retaliate against another person. Sometimes cruelty toward animals occurs as revenge against other people.
- To divert hostility from a person to an animal. This deviated aggression usually involves authority figures whom the subject hates or fears but who they are afraid to aggress against. For example, in childhood, it is often easier to be violent to an animal than against a parent or adult
- Non-specific sadism. This violence is related to the desire to inflict harm, suffering, or death on an animal in the absence of any particular provocation or especially hostile feelings toward an animal.
1.2. The Animal as the Perpetrator
2. Role of the Veterinary Pathologist in Veterinary Forensic Investigations
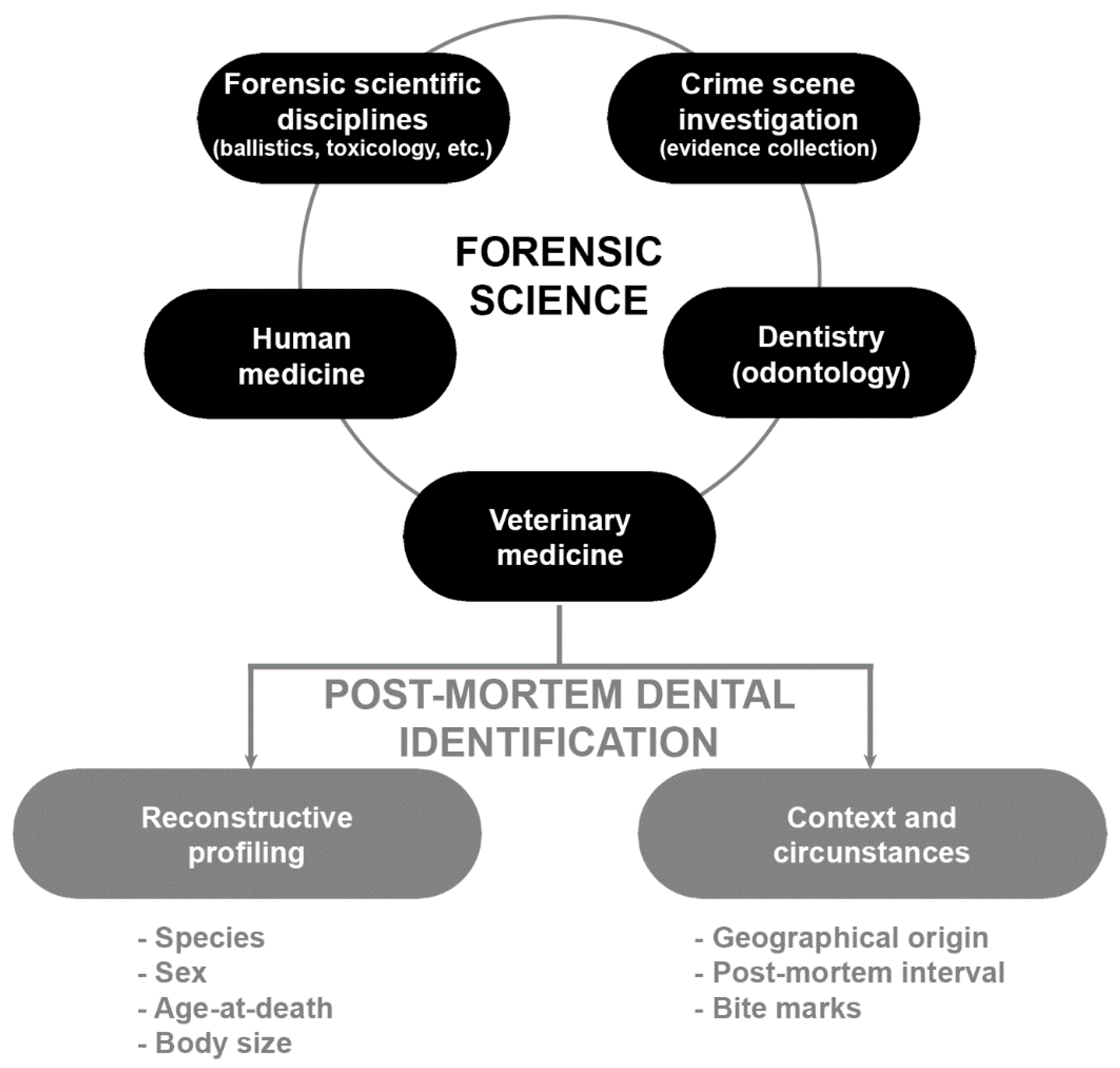
3. Teeth as a Biological Source for Forensic Identification in Animal Remains
3.1. Biological Profile
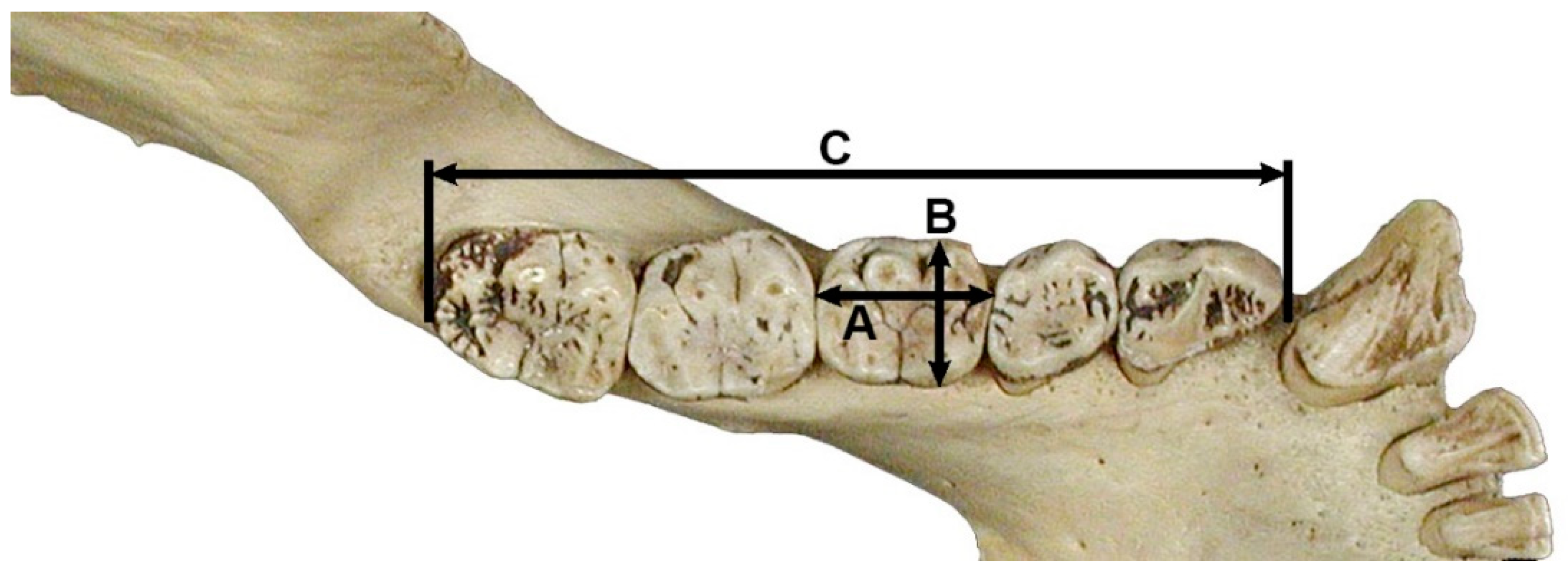
3.1.1. Species Identification
3.1.2. Sex Estimation
3.1.3. Age-at-Death Estimation
3.1.4. Body Size Estimation
3.2. Geographical Origin (Provenance) Identification
3.3. Post-Mortem Interval Estimation
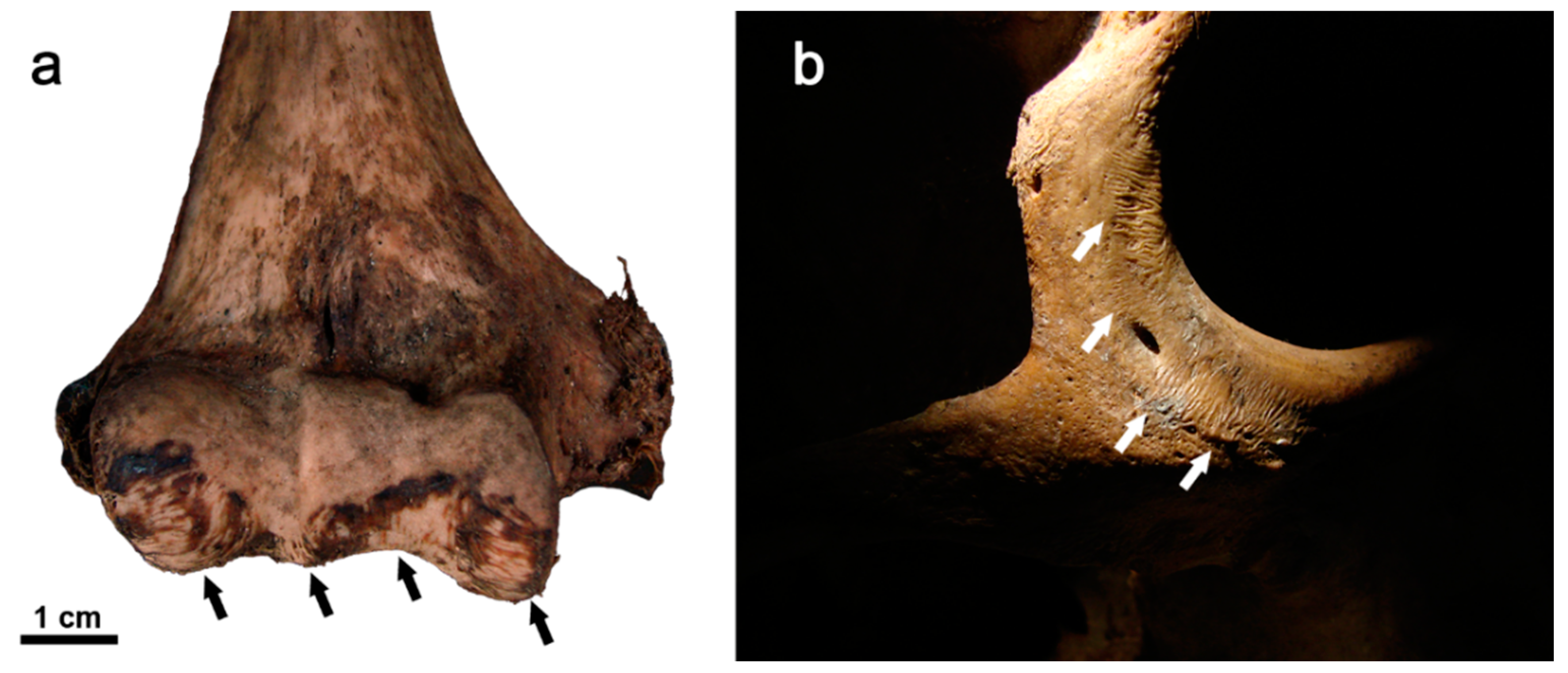
3.4. Bite Marks
3.4.1. Animal Scavenging
3.4.2. Human Deaths from Animal Bites
4. Application Potentialities in Other Contexts
4.1. Archaeological and Palaeoanthropological Contexts
4.2. Ecological Contexts: Study of the Biodiversity of Wild Animal Species
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bailey, D. (Ed.) Introduction—What is the veterinary forensics? In Practical Veterinary Forensics; CAB International: Oxfordshire, UK, 2016; pp. 1–11. [Google Scholar]
- Cooper, J.E.; Cooper, M.E. Special features of veterinary and comparative forensic medicine. In Introduction to Veterinary and Comparative Forensic Medicine; Cooper, J.E., Cooper, M.E., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2007; pp. 168–224. [Google Scholar]
- De Siqueira, A.; Cuevas, S.E.C.; Salvagni, F.A.; Maiorka, P.C. Forensic veterinary pathology: Sharp injuries in animals. Vet. Pathol. 2016, 53, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Forbes, N. An exacting science: The veterinary surgeon as expert witness. In Practice 2004, 26, 503–506. [Google Scholar] [CrossRef]
- Cooper, J.E.; Cooper, M.E. Forensic veterinary medicine: A rapidly evolving discipline. Forensic Sci. Med. Pathol. 2008, 4, 75–82. [Google Scholar] [CrossRef]
- Ledger, R.A.; Mellor, D.J. Forensic use of the five domains model for assessing suffering in cases of animal cruelty. Animals 2018, 8, 101. [Google Scholar] [CrossRef] [PubMed]
- Arkow, P.; Munro, H. The veterinary profession’s roles in recognizing and preventing family violence: The experiences of the human medicine field and the development of diagnostic indicators of non-accidental injury. In The International Handbook of Animal Abuse and Cruelty: Theory, Research, and Application; Ascione, F., Ed.; Purdue University Press: West Lafayette, IN, USA, 2008; pp. 31–58. [Google Scholar]
- McMillan, F.D.; Duffy, D.L.; Serpell, J.A. Mental health of dogs formerly used as ‘breeding stock’ in commercial breeding establishments. Appl. Anim. Behav. Sci. 2011, 135, 86–94. [Google Scholar] [CrossRef]
- Munro, H.M.; Thrusfield, M.V. ‘Battered pets’: Non-accidental physical injuries found in dogs and cats. J. Small Anim. Pract. 2001, 42, 279–290. [Google Scholar] [CrossRef]
- Rebollada-Merino, A.; Bárcena, C.; Mayoral-Alegre, F.J.; García-Real, I.; Domínguez, L.; Rodríguez-Bertos, A. Forensic cases of suspected dog and cat abuse in the Community of Madrid (Spain), 2014–2019. Forensic Sci. Int. 2020, 316, 110522. [Google Scholar] [CrossRef] [PubMed]
- Proulx, G.; Rodtka, D. Killing traps and snares in North America: The need for stricter checking time periods. Animals 2019, 9, 570. [Google Scholar] [CrossRef]
- White, P.A.; Valkenburgh, B.V. Low-cost forensics reveal high rates of non-lethal snaring and shotgun injuries in Zambia’s large carnivores. Front. Conserv. Sci. 2022, 3, 803381. [Google Scholar] [CrossRef]
- Grilo, A.; Moreira, A.; Carrapiço, B.; Belas, A.; São Braz, B. Epidemiological study of pesticide poisoning in domestic animals and wildlife in Portugal: 2014–2020. Front. Vet. Sci. 2021, 7, 616293. [Google Scholar] [CrossRef]
- Rial-Berriel, C.; Acosta-Dacal, A.; Zumbado, M.; Henríquez-Hernández, L.A.; Rodríguez-Hernández, Á.; Macías-Montes, A.; Boada, L.D.; Travieso-Aja, M.M.; Martin-Cruz, B.; Suárez-Pérez, A.; et al. Epidemiology of animal poisonings in the Canary Islands (Spain) during the period 2014–2021. Toxics 2021, 9, 267. [Google Scholar] [CrossRef] [PubMed]
- Pankowski, F.; Bogiel, G.; Paśko, S.; Rzepiński, F.; Misiewicz, J.; Staszak, A.; Bonecka, J.; Dzierzęcka, M.; Bartyzel, B.J. Fatal gunshot injuries in the common buzzard Buteo buteo L. 1758—Imaging and ballistic findings. Forensic Sci. Med. Pathol. 2018, 14, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Alleyne, E.; Parfitt, C. Adult-perpetrated animal abuse: A systematic literature review. Trauma Violence Abuse 2019, 20, 344–357. [Google Scholar] [CrossRef]
- Kellert, S.R.; Felthous, A.R. Childhood cruelty toward animals among criminals and noncriminals. Hum. Relat. 1985, 38, 1113–1129. [Google Scholar] [CrossRef]
- Ahmad, H.I.; Ahmad, M.J.; Jabbir, F.; Ahmar, S.; Ahmad, N.; Elokil, A.A.; Chen, J. The domestication makeup: Evolution, survival, and challenges. Front. Ecol. Evol. 2020, 8, 103. [Google Scholar] [CrossRef]
- Cucchi, T.; Arbuckle, B. Animal domestication: From distant past to current development and issues. Anim. Front. 2021, 11, 6–9. [Google Scholar] [CrossRef]
- Tanga, C.; Remigio, M.; Viciano, J. Transmission of zoonotic diseases in the daily life of ancient Pompeii and Herculaneum (79 CE, Italy): A review of animal-human-environment interactions through biological, historical and archaeological sources. Animals 2022, 12, 213. [Google Scholar] [CrossRef]
- Errickson, D.; Thompson, T.J.U. Animal attacks and injuries: Anthropological findings. In Encyclopedia of Forensic and Legal Medicine, 2nd ed.; Payne-James, J., Byard, R.W., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 143–147. [Google Scholar] [CrossRef]
- Fonseca, G.M.; Palacios, R. An unusual case of predation: Dog pack or cougar attack? J. Forensic Sci. 2013, 58, 224–227. [Google Scholar] [CrossRef]
- Butler, L.; Dale, B.; Beckmen, K.; Farley, S. Findings Related to the March 2010 Fatal Wolf Attack Near Chignik Lake, Alaska; ADF&G/DWC/WSP-2011-2; Alaska Department of Fish and Game, Division of Wildlife Conservation, Wildlife Special Publication: Juneau, AK, USA, 2011. [Google Scholar]
- Ďatko, M.; Vojtíšek, T.; Hejna, P. A fatal lion attack. Forensic Sci. Med. Pathol. 2015, 11, 300–302. [Google Scholar] [CrossRef]
- Kanchan, T.; Shekhawat, R.S.; Shetty, B.S.K.; Jayaram, L.; Meshram, V.P. Fatal captive tiger attack—A case report with review of literature. J. Forensic Leg. Med. 2021, 78, 102100. [Google Scholar] [CrossRef]
- Floyd, T. Bear-inflicted human injury and fatality. Wilderness Environ. Med. 1999, 10, 75–87. [Google Scholar] [CrossRef]
- Yilmaz, A.; Hosukler, E.; Erkol, Z.; Samurcu, H.; Gundogdu, V.; Celme, H.A.; Turan, T.; Dincer, G. Old female case who died as a result of bear attack. Med. Sci. 2021, 10, 623–626. [Google Scholar] [CrossRef]
- Janda, D.H.; Ringler, D.H.; Hilliard, J.K.; Hankin, R.C.; Hankin, F.M. Nonhuman primate bites. J. Orthop. Res. 1990, 8, 146–150. [Google Scholar] [CrossRef]
- Behera, C.; Chauhan, M.; Sikary, A.; Millo, T.; Byard, R.W. Injuries and deaths caused by non-human primate attacks: Paediatric vulnerability. Med. Sci. Law 2021, 61, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Manipady, S.; Menezes, R.G.; Bastia, B.K. Death by attack from a wild boar. J. Clin. Forensic Med. 2006, 13, 89–91. [Google Scholar] [CrossRef]
- Gitto, L.; Bonaccorso, L.; Bryant, S.M.; Serinelli, S. Death caused by a domestic pig attack. Forensic Sci. Med. Pathol. 2021, 17, 469–474. [Google Scholar] [CrossRef]
- Kovalenko, F.; Vassiliu, P.; Degiannis, K.; Doll, D. Complex femoral injuries after hippopotamus bite: A case report. Albanian J. Trauma Emerg. Surg. 2021, 5, 879–882. [Google Scholar] [CrossRef]
- Yanai, O.; Goldin, L.; Hiss, J. Fatal rat bites. Harefuah 1999, 136, 611–613. [Google Scholar]
- Skarentzos, S.K.; Papadopulos, N.; Deftereos, S.P.; Thomaidis, S.; Kambouri, K. Serious rodent bites to an 8-month-old infant due to child neglect. Cureus 2021, 13, e18493. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Shee, B.; Sukul, B. Fatal crocodile attack. J. Forensic Leg. Med. 2013, 20, 1139–1141. [Google Scholar] [CrossRef]
- Boyd, B.S.; Colon, F.; Doty, J.F.; Sanders, K.C. Beware of the dragon: A case report of a Komodo dragon attack. Foot Ankle Orthop. 2021, 6, 24730114211015623. [Google Scholar] [CrossRef] [PubMed]
- Idani, M.; N’guessan, E.K.Z.; Millogo, M.; Badini, S.P.A.; Zango, A.; Ili, B.V.; Crezoit, G.E.; Konsem, T. Cranio-cervico-facial injuries due to crocodile bite: A rare observation in Yalgado Ouedraogo University Hospital (Burkina Faso). J. Oral Med. Oral Surg. 2022, 28, 12. [Google Scholar] [CrossRef]
- Byard, R.W.; James, R.A.; Heath, K.J. Recovery of human remains after shark attack. Am. J. Forensic Med. Pathol. 2006, 27, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Clua, E.; Reid, D. Features and motivation of a fatal attack by a juvenile white shark, Carcharodon carcharias, on a young male surfer in New Caledonia (South Pacific). J. Forensic Leg. Med. 2013, 20, 551–554. [Google Scholar] [CrossRef] [PubMed]
- Clua, E.; Bescond, P.M.; Reid, D. Fatal attack by a juvenile tiger shark, Galeocerdo cuvier, on a kitesurfer in New Caledonia (South Pacific). J. Forensic Leg. Med. 2014, 25, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Indra, L.; Errickson, D.; Young, A.; Lösch, S. Uncovering forensic taphonomic agents: Animal scavenging in the European context. Biology 2022, 11, 601. [Google Scholar] [CrossRef]
- Cattaneo, C.; Gibelli, D. Animal effects on bones. In Forensic Anthropology; Houck, M.M., Ed.; Academic Press: London, UK, 2017; pp. 31–35. [Google Scholar]
- Young, A. The effects of terrestrial mammalian scavenging and avian scavenging on the body. In Taphonomy of Human Remains: Forensic Analysis of the Dead and the Depositional Environment; Schotsmans, E.M.J., Márquez-Grant, N., Forbes, S.L., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2017; pp. 212–234. [Google Scholar]
- Pokines, J.T. Fauna dispersal, reconcentration, and gnawing damage to bone in terrestrial environments. In Manual of Forensic Taphonomy, 2nd ed.; Pokines, J.T., L’Abbé, E.N., Symes, S.A., Eds.; CRC Press: Boca Raton, FL, USA, 2022; pp. 295–359. [Google Scholar] [CrossRef]
- Munro, R.; Munro, H.M.C. Some challenges in forensic veterinary pathology: A review. J. Comp. Pathol. 2013, 149, 57–73. [Google Scholar] [CrossRef]
- Kles, M.; Sutton, L. Forensic veterinary osteology. In Veterinary Forensic Medicine and Forensic Sciences; Byrd, J.H., Norris, P., Bradley-Siemens, N., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 199–224. [Google Scholar]
- O’Connor, T. The Archaeology of Animal Bones; Texas A&M University Press: College Station, TX, USA, 2008. [Google Scholar]
- Gifford-Gonzalez, D. Bone’s intrinsic traits: Inferring species, sex, and age. In An Introduction to Zooarchaeology; Gifford-Gonzalez, D., Ed.; Springer: New York, NY, USA, 2018; pp. 103–123. [Google Scholar]
- Ungar, P.S. How teeth work. In Evolution’s Bite: A Story of Teeth, Diet, and Human Origins; Ungar, P.S., Ed.; Princeton University Press: Princeton, NJ, USA, 2017; pp. 5–33. [Google Scholar]
- Sims, M.E. Cranial morphology of five felids: Acinonyx jubatus, Panthera onca, Panthera pardus, Puma concolor, Uncia uncia. Russ. J. Theriol. 2012, 11, 157–170. [Google Scholar] [CrossRef]
- Roy, J.; Singh, A.; Rohith, M.M.; Sharma, L.K.; Johnson, A.; Joshi, H.; Chinnadurai, V.; Chandra, K.; Thakur, M. Cranio-dental signature of three big cats of India: Implications in wildlife forensics. Proc. Zool. Soc. 2022, 75, 57–64. [Google Scholar] [CrossRef]
- Hillson, S. Teeth, 2nd ed.; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Halstead, P.; Collins, P.; Isaakidou, V. Sorting the sheep from the goats: Morphological distinctions between the mandibles and mandibular teeth of adult Ovis and Capra. J. Archaeol. Sci. 2002, 29, 545–553. [Google Scholar] [CrossRef]
- Whitenack, L.B.; Gottfried, M.D. A morphometric approach for addressing tooth-based species delimitation in fossil mako sharks, Isurus (Elasmobranchii: Lamniformes). J. Vertebr. Paleontol. 2010, 30, 17–25. [Google Scholar] [CrossRef]
- Boroni, N.L.; Lobo, L.S.; Romano, P.S.R.; Lessa, G. Taxonomic identification using geometric morphometric approach and limited data: An example using the upper molars of two sympatric species of Calomys (Cricetidae: Rodentia). Zoologia 2017, 34, e19864. [Google Scholar] [CrossRef]
- Chuang, R.; Bonhomme, V. Rethinking the dental morphological differences between domestic equids. J. Archaeol. Sci. 2019, 101, 140–148. [Google Scholar] [CrossRef]
- Hulme-Beaman, A.; Claude, J.; Chaval, Y.; Evin, A.; Morand, S.; Vigne, J.D.; Dobney, K.; Cucchi, T. Dental shape variation and phylogenetic signal in the Rattini tribe species of Mainland Southeast Asia. J. Mammal. Evol. 2019, 26, 435–446. [Google Scholar] [CrossRef]
- Miele, V.; Dussert, G.; Cucchi, T.; Renaud, S. Deep learning for species identification of modern and fossil rodent molars. BioRxiv 2020. [Google Scholar] [CrossRef]
- Toledo-González, V.; Ortega-Ojeda, F.; Fonseca, G.M.; García-Ruiz, C.; Navarro-Cáceres, P.; Pérez-Lloret, P.; Marín-García, M.D.P. A morphological and morphometric dental analysis as a forensic tool to identify the Iberian wolf (Canis lupus signatus). Animals 2020, 10, 975. [Google Scholar] [CrossRef]
- Plavcan, J.M.; Cope, D.A. Metric variation and species recognition in the fossil record. Evol. Anthropol. 2001, 10, 204–222. [Google Scholar] [CrossRef]
- Simpson, G.G.; Roe, A.; Lewontin, R.C. Quantitative Zoology; Harcourt, Brace and Company: New York, NY, USA, 1960. [Google Scholar]
- Murmann, D.C.; Brumit, P.C.; Schrader, B.A.; Senn, D.R. A comparison of animal jaws and bite mark patterns. J. Forensic. Sci. 2006, 51, 846–860. [Google Scholar] [CrossRef]
- Cucchi, T.; Mohaseb, A.; Peigné, S.; Debue, K.; Orlando, L.; Mashkour, M. Detecting taxonomic and phylogenetic signals in equid cheek teeth: Towards new palaeontological and archaeological proxies. R. Soc. Open Sci. 2017, 4, 160997. [Google Scholar] [CrossRef]
- Berthaume, M.A.; Lazzari, V.; Guy, F. The landscape of tooth shape: Over 20 years of dental topography in primates. Evol. Anthropol. 2020, 29, 245–262. [Google Scholar] [CrossRef]
- Da Silva, R.F.; Mendes, S.D.S.C.; Marinho, D.E.A.; Júnior, A.F.R.; Guimarães, M.A. Importance of the comparative anatomy in Forensic Anthropology—Case report. RSBO 2013, 10, 193–197. [Google Scholar]
- Courtenay, L.A.; Herranz-Rodrigo, D.; González-Aguilera, D.; Yravedra, J. Developments in data science solutions for carnivore tooth pit classification. Sci. Rep. 2021, 11, 10209. [Google Scholar] [CrossRef] [PubMed]
- Yravedra, J.; Maté-González, M.Á.; Courtenay, L.A.; González-Aguilera, D.; Fernández-Fernández, M. The use of canid tooth marks on bone for the identification of livestock predation. Sci. Rep. 2019, 9, 16301. [Google Scholar] [CrossRef] [PubMed]
- Courtenay, L.A.; Yravedra, J.; Huguet, R.; Aramendi, J.; Maté-González, M.Á.; González-Aguilera, D.; Arriaza, M.C. Combining machine learning algorithms and geometric morphometrics: A study of carnivore tooth marks. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 522, 28–39. [Google Scholar] [CrossRef]
- Whittaker, D.K.; Rawle, L.W. The effect of conditions of putrefaction on species determination in human and animal teeth. Forensic Sci. Int. 1987, 35, 209–212. [Google Scholar] [CrossRef]
- Balasse, M.; Ambrose, S.H. Distinguishing sheep and goats using dental morphology and stable carbon isotopes in C4 grassland environments. J. Archaeol. Sci. 2005, 32, 691–702. [Google Scholar] [CrossRef]
- Kim, T.W.; Lee, H.J.; Kim, Y.K.; Oh, H.S.; Han, S.H. Genetic identification of prey species from teeth in faeces from the Endangered leopard cat Prionailurus bengalensis using mitochondrial cytochrome b gene sequence. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2018, 29, 170–174. [Google Scholar] [CrossRef]
- Minakshi, P.; Shafiq, M.; Alisha, A.; Ranjan, K.; Kumar, P.; Anshul, B.; Singh, R.; Basanti, S.; Prasad, G. Animal forensics and applications. In Biotechnology: Prospects and Applications; Salar, R.K., Gahlawat, S.K., Siwach, P., Duhan, J.S., Eds.; Springer: New Delhi, India, 2013; pp. 265–286. [Google Scholar] [CrossRef]
- Buddhachat, K.; Klinhom, S.; Siengdee, P.; Brown, J.L.; Nomsiri, R.; Kaewmong, P.; Thitaram, C.; Mahakkanukrauh, P.; Nganvongpanit, K. Elemental analysis of bone, teeth, horn and antler in different animal species using non-invasive handheld X-ray fluorescence. PLoS ONE 2016, 11, e0155458. [Google Scholar] [CrossRef]
- Nganvongpanit, K.; Brown, J.L.; Buddhachat, K.; Somgird, C.; Thitaram, C. Elemental analysis of Asian elephant (Elephas maximus) teeth using X-ray fluorescence and a comparison to other species. Biol. Trace Elem. Res. 2016, 170, 94–105. [Google Scholar] [CrossRef]
- Nganvongpanit, K.; Buddhachat, K.; Klinhom, S.; Kaewmong, P.; Thitaram, C.; Mahakkanukrauh, P. Determining comparative elemental profile using handheld X-ray fluorescence in humans, elephants, dogs, and dolphins: Preliminary study for species identification. Forensic Sci. Int. 2016, 263, 101–106. [Google Scholar] [CrossRef]
- Irfanita, N.; Jaswir, I.; Mirghani, M.E.S.; Sukmasari, S.; Ardini, Y.D.; Lestari, W. Rapid detection of gelatin in dental materials using Attenuated Total Reflection Fourier Transform Infrared spectroscopy (ATR-FTIR). J. Phys. Conf. Ser. 2017, 884, 012090. [Google Scholar] [CrossRef]
- Wei, X.; Yu, K.; Wu, H.; Shen, C.; Li, H.; Liu, R.; Sun, Q.; Wang, Z. Species identification of teeth of human and non-human. Forensic Sci. Int. 2022, 333, 111205. [Google Scholar] [CrossRef] [PubMed]
- Caniglia, R.; Fabbri, F.; Mastrogiuseppe, L.; Randia, E. Who is who? Identification of livestock predators using forensic genetic approaches. Forensic Sci. Int. Genet. 2013, 7, 397–404. [Google Scholar] [CrossRef]
- Toledo-González, V.; Ortega-Ojeda, F.; Fonseca, G.M.; García-Ruiz, C.; Pérez-Lloret, P. Analysis of tooth mark patterns on bone remains caused by wolves (Canis lupus) and domestic dogs (Canis lupus familiaris) for taxonomic identification: A scoping review focused on their value as a forensic tool. Appl. Anim. Behav. Sci. 2021, 240, 105356. [Google Scholar] [CrossRef]
- McPherson, F.J.; Chenoweth, P.J. Mammalian sexual dimorphism. Anim. Reprod. Sci. 2012, 131, 109–122. [Google Scholar] [CrossRef]
- Lough-Stevens, M.; Schultz, N.G.; Dean, M.D. The baubellum is more developmentally and evolutionarily labile than the baculum. Ecol. Evol. 2017, 8, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Szuma, E. Geography of sexual dimorphism in the tooth size of the red fox Vulpes vulpes (Mammalia, Carnivora). J. Zool. Syst. Evol. Res. 2008, 46, 73–81. [Google Scholar] [CrossRef]
- Weckerly, F.W. Sexual-size dimorphism: Influence of mass and mating systems in the most dimorphic mammals. J. Mammal. 1998, 79, 33–52. [Google Scholar] [CrossRef]
- Gittleman, J.L.; Valkenburgh, B.V. Sexual dimorphism in the canines and skulls of carnivores: Effects of size, phylogeny, and behavioural ecology. J. Zool. 1997, 242, 97–117. [Google Scholar] [CrossRef]
- Gingerich, P.D. Variation, sexual dimorphism, and social structure in the early Eocene horse Hyracotherium (Mammalia, Perissodactyla). Paleobiology 1981, 7, 443–455. [Google Scholar] [CrossRef]
- Lüps, P.; Roper, T.J. Tooth size in the European badger (Meles meles) with special reference to sexual dimorphism, diet and intraspecific aggression. Acta Theriol. 1988, 33, 21–33. [Google Scholar] [CrossRef]
- Mayer, J.J.; Brisbin, I.L., Jr. Sex identification of Sus scrofa based on canine morphology. J. Mammal. 1988, 69, 408–412. [Google Scholar] [CrossRef]
- Lowry, M.S.; Folk, R.L. Sex determination of the California sea lion (Zalophus californianus) from canine teeth. Mar. Mammal Sci. 1990, 6, 25–31. [Google Scholar] [CrossRef]
- Valkenburgh, B.V.; Ruff, C.B. Canine tooth strength and killing behaviour in large carnivores. J. Zool. 1987, 212, 379–397. [Google Scholar] [CrossRef]
- Turner, A. Dental sex dimorphism in European lions (Panthera leo L.) of the Upper Pleistocene: Palaeoecological and palaeoethological implications. Ann. Zool. Fenn. 1984, 21, 1–8. [Google Scholar]
- Christiansen, P.; Harris, J.M. Variation in craniomandibular morphology and sexual dimorphism in pantherines and the sabercat Smilodon fatalis. PLoS ONE 2012, 7, e48352. [Google Scholar] [CrossRef]
- Szuma, E. Variation and correlation patterns in the dentition of the red fox from Poland. Ann. Zool. Fenn. 2000, 37, 113–127. [Google Scholar]
- Kurtén, B. Sex dimorphism and size trends in the cave bear, Ursus spelaeus Rosenmüller and Heinroth. Acta Zool. Fenn. 1955, 90, 1–48. [Google Scholar]
- Larsen, T. Sexual dimorphism in the molar rows of the polar bear. J. Wildl. Manag. 1971, 35, 374–377. [Google Scholar] [CrossRef]
- Harvey, P.H.; Kavanagh, M.; Clutton-Brock, T.H. Sexual dimorphism in primate teeth. J. Zool. Lond. 1978, 186, 474–485. [Google Scholar] [CrossRef]
- Beauchamp, G. Canine tooth size variability in primates. Folia Primatol. 1989, 52, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Lucas, P.W.; Corlett, R.T.; Luke, D.A. Sexual dimorphism of tooth size in anthropoids. Hum. Evol. 1986, 1, 23–39. [Google Scholar] [CrossRef]
- Plavcan, J.M. Sexual dimorphism in primate evolution. Am. J. Phys. Anthropol. 2001, 116 (Suppl. S33), 25–53. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, G.T.; Dean, C. Ontogeny of canine dimorphism in extant hominoids. Am. J. Phys. Anthropol. 2001, 115, 269–283. [Google Scholar] [CrossRef]
- Plavcan, J.M. Sexual size dimorphism, canine dimorphism, and male-male competition in primates: Where do humans fit in? Hum. Nat. 2012, 23, 45–67. [Google Scholar] [CrossRef]
- Garn, S.; Kerewsky, R.; Swindler, D. Canine “field” in sexual dimorphism of tooth size. Nature 1966, 212, 1501–1502. [Google Scholar] [CrossRef]
- Gingerich, P.D.; Ryan, A.S. Dental and cranial variation in living Indriidae. Primates 1979, 20, 141–159. [Google Scholar] [CrossRef][Green Version]
- Gingerich, P.D.; Schoeninger, M.J. Patterns of tooth size variability in the dentition of primates. Am. J. Phys. Anthropol. 1979, 51, 457–465. [Google Scholar] [CrossRef]
- Plavcan, J.M.; van Schaik, C.P. Intrasexual competition and canine dimorphism in anthropoid primates. Am. J. Phys. Anthropol. 1992, 87, 461–477. [Google Scholar] [CrossRef]
- Swindler, D.R. Appendix 1: Odontometry. In Primate Dentition: An Introduction to the Teeth of Non-Human Primates; Swindler, D.R., Ed.; Cambridge University Press: Cambridge, UK, 2002; pp. 165–219. [Google Scholar]
- Sriraman, P.K. Forensic necropsy. In Wildlife Necropsy and Forensics; Sriraman, P.K., Ed.; CRC Press: Oxon, UK, 2021; pp. 152–199. [Google Scholar]
- Cooper, J.E.; Cooper, M.E. Clinical work. In Introduction to Veterinary and Comparative Forensic Medicine; Cooper, J.E., Cooper, M.E., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2007; pp. 117–167. [Google Scholar]
- Mendes-Oliveira, A.C.; Maria, S.L.S.; Lima, R.C.S.; Fernandes, A.S.; Almeida, P.C.R.; Montag, L.F.; Cavalho, O., Jr. Testing simple criteria for de age estimation of six hunted mammal species in Brazilian Amazon. Mastozool. Neotrop. 2012, 19, 105–116. [Google Scholar]
- Cooper, J.E.; Cooper, M.E. Importance and application of animal law. In Introduction to Veterinary and Comparative Forensic Medicine; Cooper, J.E., Cooper, M.E., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2007; pp. 42–60. [Google Scholar]
- Modina, S.C.; Andreis, M.E.; Moioli, M.; Di Giancamillo, M. Age assessment in puppies: Coming to terms with forensic requests. Forensic Sci. Int. 2019, 297, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Ahn, J.; Kang, S.; Lee, E.; Kim, S.; Park, S.; Park, S.; Noh, H.; Seo, K. Determining the age of cats by pulp cavity/tooth width ratio using dental radiography. J. Vet. Med. Sci. 2014, 15, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Roccaro, M.; Peli, A. Age determination in dog puppies by teeth examination: Legal, health and welfare implications, review of the literature and practical considerations. Vet. Ital. 2020, 56, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.E.; Cooper, M.E. Pathology and post-mortem examinations. In Introduction to Veterinary and Comparative Forensic Medicine; Cooper, J.E., Cooper, M.E., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2007; pp. 168–224. [Google Scholar]
- Mbizah, M.M.; Steenkamp, G.; Groom, R.J. Evaluation of the applicability of different age determination methods for estimating age of the endangered African wild dog (Lycaon pictus). PLoS ONE 2016, 11, e0164676. [Google Scholar] [CrossRef]
- Sutton, L.K.; Byrd, J.H.; Brooks, J.W. Age determination in dogs and cats. In Veterinary Forensic Pathology; Brooks, J.W., Ed.; Springer: Cham, Switzerland, 2018; Volume 2, pp. 151–163. [Google Scholar]
- Adserias-Garriga, J. Evolution of methods and state-of-the-art in dental age estimation. In Age Estimation; Adserias-Garriga, J., Ed.; Academic Press: London, UK, 2019; pp. 77–87. [Google Scholar]
- O’Connor, T.P. Vertebrate demography by numbers: Age, sex, and zooarchaeological practice. In Recent Advances in Ageing and Sexing Animal Bones; Ruscillo, D., Ed.; Oxbow Books: Oxford, UK, 2006; pp. 1–8. [Google Scholar]
- Lemons, M.; Beebe, D. Oral anatomy and physiology. In Wiggs’s Veterinary Dentistry: Principles and Practice, 2nd ed.; Lobprise, H.B., Dodd, J.R., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2019; pp. 1–24. [Google Scholar]
- Spinage, C.A. A review of age determination of mammals by means of teeth, with special reference to Africa. East Afr. Wildl. J. 1973, 11, 165–187. [Google Scholar] [CrossRef]
- Silver, I.A. The ageing of domestic animals. In Science in Archaeology: A Comprehensive Survey of Progress and Research, 2nd ed.; Brothwell, D., Higgs, E., Eds.; Basic Books: New York, NY, USA, 1963; pp. 250–268. [Google Scholar]
- Lorászkó, G.; Rácz, B.; Ózsvári, L. Changes in the dentition of small dogs up to 4 months of age. Animals 2022, 12, 1417. [Google Scholar] [CrossRef]
- Kelley, J.; Schwartz, G.T.; Smith, T.M. Age at first molar emergence in Pan troglodytes verus and variation in the timing of molar emergence among free-living chimpanzees. J. Hum. Evol. 2020, 145, 102823. [Google Scholar] [CrossRef]
- Mahoney, P. Root growth and dental eruption in modern human deciduous teeth with preliminary observations on great apes. J. Hum. Evol. 2019, 129, 46–53. [Google Scholar] [CrossRef]
- Holly Smith, B.; Crummett, T.L.; Brandt, K.L. Ages of eruption of primate teeth: A compendium for aging individuals and comparing life histories. Am. J. Phys. Anthropol. 1994, 37, 177–231. [Google Scholar] [CrossRef]
- Smith, T.D.; Muchlinski, M.N.; Jankord, K.D.; Progar, A.J.; Bonar, C.J.; Evans, S.; Williams, L.; Vinyard, C.J.; Deleon, V.B. Dental maturation, eruption, and gingival emergence in the upper jaw of newborn primates. Anat. Rec. 2015, 298, 2098–2131. [Google Scholar] [CrossRef]
- Greenfield, H.J.; Arnold, E.R. Absolute age and tooth eruption and wear sequences in sheep and goat: Determining age-at-death in zooarchaeology using a modern control sample. J. Archaeol. Sci. 2008, 35, 836–849. [Google Scholar] [CrossRef]
- Moran, N.C.; O’Connor, T.P. Age attribution in domestic sheep by skeletal and dental maturation: A pilot study of available sources. Int. J. Osteoarchaeol. 1994, 4, 267–285. [Google Scholar] [CrossRef]
- Andrews, A.H. The relationship of bovine mandibular cheek tooth development to age determined by post-mortem radiographic examination of cattle aged between 12 and 24 months. J. Agric. Sci. 1982, 98, 109–117. [Google Scholar] [CrossRef]
- Oroian, T.E.; Oroian, R.G.; Pasca, I.; Oroian, E.; Covrig, L. Methods of age estimation by dentition in Sus scrofa ferus sp. Bull. Univ. Agric. Sci. Vet. Med. 2010, 67, 291–295. [Google Scholar] [CrossRef]
- Payne, S. Kill-off patterns in sheep and goats: The mandibles from Aşvan Kale. Anatol. Stud. 1973, 23, 281–303. [Google Scholar] [CrossRef]
- Grant, A. The use of tooth wear as a guide to the age of domestic ungulates. In Ageing and Sexing Animal Bones from Archaeological Sites; Wilson, B., Grigson, C., Payne, S., Eds.; British Archaeological Reports: Oxford, UK, 1982; pp. 91–108. [Google Scholar]
- Spinage, C.A. Age estimation of zebra. East Afr. Wildl. J. 1972, 10, 273–277. [Google Scholar] [CrossRef]
- Levine, M.A. The use of crown height measurements and eruption-wear sequences to age horse teeth. In Ageing and Sexing Animal Bones from Archaeological Sites; Wilson, B., Grigson, C., Payne, S., Eds.; British Archaeological Reports: Oxford, UK, 1982; pp. 223–250. [Google Scholar]
- Klein, R.G.; Wolf, C.; Freeman, L.G.; Allwarden, K. The use of dental crown heights for constructing age profiles of red deer and similar species in archaeological samples. J. Archaeol. Sci. 1981, 8, 1–31. [Google Scholar] [CrossRef]
- Lieberman, D.E. The biological basis for seasonal increments in dental cementum and their application to archaeological research. J. Archaeol. Sci. 1994, 21, 525–539. [Google Scholar] [CrossRef]
- Veiberg, V.; Nilsen, E.B.; Rolandsen, C.M.; Heim, M.; Andersen, R.; Holmstrøm, F.; Meisingset, E.L.; Solberg, E.J. The accuracy and precision of age determination by dental cementum annuli in four northern cervids. Eur. J. Wildl. Res. 2020, 66, 91. [Google Scholar] [CrossRef]
- Wolfe, M.L. Age determination in moose from cemental layers of molar teeth. J. Wildl. Manag. 1969, 33, 428–431. [Google Scholar] [CrossRef]
- Low, W.A.; Cowan, I.M.T. Age determination of deer by annular structure of dental cementum. J. Wildl. Manag. 1963, 27, 466–471. [Google Scholar] [CrossRef]
- Stoneberg, R.P.; Jonkel, C.J. Age determination of black bears by cementum layers. J. Wildl. Manag. 1966, 30, 411–414. [Google Scholar] [CrossRef]
- Medill, S.; Derocher, A.E.; Stirling, I.; Lunn, N.; Moses, R.A. Estimating cementum annuli width in polar bears: Identifying sources of variation and error. J. Mammal. 2009, 90, 1256–1264. [Google Scholar] [CrossRef]
- Christensen-Dalsgaard, S.N.; Aars, J.; Andersen, M.; Lockyer, C.; Yoccoz, N.G. Accuracy and precision in estimation of age of Norwegian Arctic polar bears (Ursus maritimus) using dental cementum layers from known-age individuals. Polar Biol. 2010, 33, 589–597. [Google Scholar] [CrossRef]
- Mansfield, A.W.; Fisher, H.D. Age determination in the harbour seal, Phoca vitulina L. Nature 1960, 186, 92–93. [Google Scholar] [CrossRef]
- Stewart, R.E.A.; Stewart, B.E.; Stirling, I.; Street, E. Counts of growth layer groups in cementum and dentine in ringed seals (Phoca hispida). Mar. Mammal Sci. 1996, 12, 383–401. [Google Scholar] [CrossRef]
- Crowe, D.M.; Strickland, M.D. Dental annulation in the American badger. J. Mammal. 1975, 56, 269–272. [Google Scholar] [CrossRef]
- Clarke, C.M.H.; Dzieciolowski, R.M.; Batcheler, D.; Frampton, C.M. A comparison of tooth eruption and wear and dental cementum techniques in age determination of New Zealand feral pigs. Wildl. Res. 1992, 19, 769–777. [Google Scholar] [CrossRef]
- Nomokonova, T.; Losey, R.J.; McLachlin, K.; Bachura, O.P.; Gusev, A.V.; Kosintsev, P.A.; Fedorovad, N.V.; Sablin, M.V. Age estimation of archaeological dogs using pulp cavity closure ratios. J. Archaeol. Sci. 2020, 123, 105252. [Google Scholar] [CrossRef]
- Knowlton, F.F.; Whittemore, S.L. Pulp cavity-tooth width ratios from known-age and wild-caught coyotes determined by radiography. Wildl. Soc. Bull. 2001, 29, 239–244. [Google Scholar]
- White, P.A.; Ikanda, D.; Ferrante, L.; Chardonnet, P.; Mesochina, P.; Cameriere, R. Age estimation of African lions Panthera leo by ratio of tooth Areas. PLoS ONE 2016, 11, e0153648. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, S.S.B. Estimation of body size in fossil mammals. In Methods in Paleoecology: Reconstructing Cenozoic Terrestrial Environments and Ecological Communities; Croft, D.A., Su, D.F., Simpson, S.W., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 7–22. [Google Scholar] [CrossRef]
- Christiansen, F.; Sironi, M.; Moore, M.J.; Di Martino, M.; Ricciardi, M.; Warick, H.A.; Irschick, D.J.; Gutierrez, R.; Uhart, M.M. Estimating body mass of free-living whales using aerial photogrammetry and 3D volumetrics. Methods Ecol. Evol. 2019, 10, 2034–2044. [Google Scholar] [CrossRef]
- Iskjaer, C.; Slade, N.A.; Childs, J.E.; Glass, G.E.; Korch, G.W. Body mass as a measure of body size in small mammals. J. Mammal. 1989, 70, 662–667. [Google Scholar] [CrossRef]
- Chrószcz, A.; Janeczek, M.; Onar, V.; Staniorowski, P.; Pospieszny, N. The shoulder height estimation in dogs based on the internal dimension of cranial cavity using mathematical formula. Anat. Histol. Embryol. 2007, 36, 269–271. [Google Scholar] [CrossRef]
- Chrószcz, A.; Janeczek, M.; Pasicka, E.; Klećkowska-Nawrot, J. Height at the withers estimation in the horses based on the internal dimension of cranial cavity. Folia Morphol. 2014, 73, 143–148. [Google Scholar] [CrossRef]
- Gingerich, P.D. Prediction of body mass in mammalian species from long bone lengths and diameters. In Contributions from the Museum of Paleontology; The University of Michigan: Ann Arbor, MI, USA, 1990; Volume 28, pp. 79–92. [Google Scholar]
- Campione, N.E.; Evans, D.C. A universal scaling relationship between body mass and proximal limb bone dimensions in quadrupedal terrestrial tetrapods. BMC Biol. 2012, 10, 60. [Google Scholar] [CrossRef]
- Ruff, C.B. Body mass and hindlimb bone cross-sectional and articular dimensions in anthropoid primates. In Body Size in Mammalian Paleobiology: Estimation and Biological Implications; Damuth, J., MacFadden, B.J., Eds.; Cambridge University Press: Cambridge, UK, 1990; pp. 119–150. [Google Scholar]
- Copes, L.E.; Schwartz, G.T. The scale of it all: Postcanine tooth size, the taxon-level effect, and the universality of Gould’s scaling law. Paleobiology 2010, 36, 188–203. [Google Scholar] [CrossRef]
- Mendoza, M.; Janis, C.M.; Palmqvist, P. Estimating the body mass of extinct ungulates: A study on the use of multiple regression. J. Zool. 2006, 270, 90–101. [Google Scholar] [CrossRef]
- Saarinen, J.; Cirilli, O.; Strani, F.; Meshida, K.; Bernor, R.L. Testing equid body mass estimate equations on modern zebras—With implications to understanding the relationship of body size, diet, and habitats of Equus in the Pleistocene of Europe. Front. Ecol. Evol. 2021, 9, 622412. [Google Scholar] [CrossRef]
- Gould, S.J. On the scaling of tooth size in mammals. Am. Zool. 1975, 15, 351–362. [Google Scholar] [CrossRef]
- McNab, B. Complications inherent in scaling the basal rate of metabolism in mammals. Q. Rev. Biol. 1988, 63, 25–54. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.R. Preliminary body mass estimates for mammalian genera of the Morrison Formation (Upper Jurassic, North America). PaleoBios 2009, 28, 114–122. [Google Scholar]
- Losey, R.J.; Osipov, B.; Sivakumaran, R.; Nomokonova, T.; Kovychev, E.V.; Diatchina, N.G. Estimating body mass in dogs and wolves using cranial and mandibular dimensions: Application to Siberian canids. Int. J. Osteoarchaeol. 2015, 25, 946–959. [Google Scholar] [CrossRef]
- Freudenthal, M.; Martín-Suárez, E. Estimating body mass of fossil rodents. Scr. Geol. 2013, 145, 1–130. [Google Scholar]
- Gingerich, P.D.; Smith, B.H.; Rosenberg, K. Allometric scaling in the dentition of primates and prediction of body weight from tooth size in fossils. Am. J. Phys. Anthropol. 1982, 58, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Valkenburgh, B.V.; Wang, X.; Damuth, J. Cope’s rule, hypercarnivory, and extinction in North American canids. Science 2004, 306, 101–104. [Google Scholar] [CrossRef]
- MacFadden, B. Fossil horses from “Eohippus” (Hyracotherium) to Equus: Scaling, Cope’s Law, and the evolution of body size. Paleobiology 1986, 12, 355–369. [Google Scholar] [CrossRef]
- Gordon, C.L. A first look at estimating body size in dentally conservative marsupials. J. Mamm. Evol. 2003, 10, 1–21. [Google Scholar] [CrossRef]
- Creighton, G.K. Static allometry of mammalian teeth and the correlation of tooth size and body size in contemporary mammals. J. Zool. 1980, 191, 435–443. [Google Scholar] [CrossRef]
- Legendre, S. Analysis of mammalian communities from the late Eocene and Oligocene of southern France. Palaeovertebrata 1986, 16, 191–212. [Google Scholar]
- Legendre, S.; Roth, C. Correlation of carnassial tooth size and body weight in recent carnivores (Mammalia). Hist. Biol. 1988, 1, 85–98. [Google Scholar] [CrossRef]
- Morris, B.; Mead, A.J. Body mass estimates from bone and tooth measurements in white-tailed deer, Odocoileus virginianus. Ga. J. Sci. 2016, 74, 18. [Google Scholar]
- Gingerich, P.D. Size variability of the teeth in living mammals and the diagnosis of closely related sympatric fossil species. J. Paleontol. 1974, 48, 895–903. [Google Scholar]
- Miller, E.H.; Mahoney, S.P.; Kennedy, M.L.; Kennedy, P.K. Variation, sexual dimorphism, and allometry in molar size of the black bear. J. Mammal. 2009, 90, 491–503. [Google Scholar] [CrossRef]
- Hopkins, S.S.B. Reassessing the mass of exceptionally large rodents using toothrow length and area as proxies for body mass. J. Mammal. 2008, 89, 232–243. [Google Scholar] [CrossRef][Green Version]
- Moncunill-Solé, B.; Jordana, X.; Marín-Moratalla, N.; Moyà-Solà, S.; Köhler, M. How large are the extinct giant insular rodents? New body mass estimations from teeth and bones. Integr. Zool. 2014, 9, 197–212. [Google Scholar] [CrossRef]
- Dagosto, M.; Terranova, C.J. Estimating the body size of Eocene primates: A comparison of results from dental and postcranial variables. Int. J. Primatol. 1992, 13, 307. [Google Scholar] [CrossRef]
- Lauer, C. The relationship of tooth size to body size in a population of rhesus monkeys (Macaca mulatta). Am. J. Phys. Anthropol. 1975, 43, 333–339. [Google Scholar] [CrossRef]
- Perez, V.J.; Leder, R.M.; Badaut, T. Body length estimation of Neogene macrophagous lamniform sharks (Carcharodon and Otodus) derived from associated fossil dentitions. Palaeontol. Electron. 2021, 4, a09. [Google Scholar] [CrossRef]
- Rubenstein, D.R.; Hobson, K.A. From birds to butterflies: Animal movement patterns and stable isotopes. Trends Ecol. Evol. 2004, 19, 256–263. [Google Scholar] [CrossRef]
- Hobson, K. Tracing origins and migration of wildlife using stable isotopes: A review. Oecologia 1999, 120, 314–326. [Google Scholar] [CrossRef] [PubMed]
- Reitsema, L.J. Laboratory and field methods for stable isotope analysis in human biology. Am. J. Hum. Biol. 2015, 27, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Schuba, J.; Südekum, K.H.; Pfeffer, E.; Jayanegara, A. Excretion of faecal, urinary urea and urinary non-urea nitrogen by four ruminant species as influenced by dietary nitrogen intake: A meta-analysis. Livest. Sci. 2017, 198, 82–88. [Google Scholar] [CrossRef]
- Lourenço, P.M.; Granadeiro, J.P.; Guilherme, J.L.; Catry, T. Turnover rates of stable isotopes in avian blood and toenails: Implications for dietary and migration studies. J. Exp. Mar. Biol. Ecol. 2015, 472, 89–96. [Google Scholar] [CrossRef]
- Bearhop, S.; Furness, R.W.; Hilton, G.M.; Votier, S.C.; Waldron, S. A forensic approach to understanding diet and habitat use from stable isotope analysis of (avian) claw material. Funct. Ecol. 2003, 17, 270–275. [Google Scholar] [CrossRef]
- Ayliffe, L.K.; Cerling, T.E.; Robinson, T.; West, A.G.; Sponheimer, M.; Passey, B.H.; Hammer, J.; Roeder, B.; Dearing, M.D.; Ehleringer, J.R. Turnover of carbon isotopes in tail hair and breath CO2 of horses fed an isotopically varied diet. Oecologia 2004, 139, 11–22. [Google Scholar] [CrossRef]
- Evans-Ogden, L.J.; Hobson, K.A.; Lank, D.B. Blood isotopic (δ13C and δ15N) turnover and diet-tissue fractionation factors in captive Dunlin (Calidris Alpina Pacifica). Auk 2004, 121, 170–177. [Google Scholar] [CrossRef]
- Bauchinger, U.; McWilliams, S. Carbon turnover in tissues of a passerine bird: Allometry, isotopic clocks, and phenotypic flexibility in organ size. Physiol. Biochem. Zool. 2009, 82, 787–797. [Google Scholar] [CrossRef]
- Madigan, D.J.; Snodgrass, O.E.; Hyde, J.R.; Dewar, H. Stable isotope turnover rates and fractionation in captive California yellowtail (Seriola dorsalis): Insights for application to field studies. Sci. Rep. 2021, 11, 4466. [Google Scholar] [CrossRef]
- Hedges, R.E.; Clement, J.G.; Thomas, C.D.; O’Connell, T.C. Collagen turnover in the adult femoral mid-shaft: Modeled from anthropogenic radiocarbon tracer measurements. Am. J. Phys. Anthropol. 2007, 133, 808–816. [Google Scholar] [CrossRef]
- Hobson, K.A.; Clark, R.G. Assessing avian diets using stable isotopes I: Turnover of 13C in tissues. Condor 1992, 94, 181–188. [Google Scholar] [CrossRef]
- Matsubayashi, J.; Tayasu, I. Collagen turnover and isotopic records in cortical bone. J. Archaeol. Sci. 2019, 106, 37–44. [Google Scholar] [CrossRef]
- Gage, J.P.; Francis, M.J.O.; Triffit, J.T. Collagen and Dental Matrices; Butterworth-Heinemann: London, UK, 1989. [Google Scholar]
- Hoppe, K.A.; Koch, P.L.; Furutani, T.T. Assessing the preservation of biogenic strontium in fossil bones and tooth enamel. Int. J. Osteoarchaeol. 2003, 13, 20–28. [Google Scholar] [CrossRef]
- Meier-Augenstein, W. Chapter III.5: Provenancing people. In Stable Isotope Forensics: Methods and Forensic Applications of Stable Isotope Analysis, 2nd ed.; Meier-Augenstein, W., Ed.; Wiley: Chichester, UK, 2018; pp. 333–400. [Google Scholar]
- Beard, B.L.; Johnson, C.M. Strontium isotope composition of skeletal material can determine the birth place and geographic mobility of humans and animals. J. Forensic Sci. 2000, 45, 1049–1061. [Google Scholar] [CrossRef] [PubMed]
- Hale, C.; Ogden, R.; Ciavaglia, S.A.; Cook, G.T.; Clarke, G.; Ogle, S.; Webster, L.M.I. Investigating the origins of ivory recovered in the United Kingdom. Forensic Sci. Int. Anim. Environ. 2021, 1, 100027. [Google Scholar] [CrossRef]
- Wyatt, T.; Johnson, K.; Hunter, L.; George, R.; Gunter, R. Corruption and wildlife trafficking: Three case studies involving Asia. Asian J. Criminol. 2018, 13, 35–55. [Google Scholar] [CrossRef]
- Alexander, J.; Downs, C.T.; Butler, M.; Woodborne, S.; Symes, C.T. Stable isotope analyses as a forensic tool to monitor illegally traded African grey parrots. Anim. Conserv. 2019, 22, 134–143. [Google Scholar] [CrossRef]
- Ziegler, S.; Merker, S.; Streit, B.; Boner, M.; Jacob, D.E. Towards understanding isotope variability in elephant ivory to establish isotopic profiling and source-area determination. Biol. Conserv. 2016, 197, 154–163. [Google Scholar] [CrossRef]
- Van der Merwe, N.; Lee-Thorp, J.; Thackeray, J.; Hall-Martin, A.; Kruger, F.J.; Coetzee, H.; Bell, R.H.V.; Lindeque, M. Source-area determination of elephant ivory by isotopic analysis. Nature 1990, 346, 744–746. [Google Scholar] [CrossRef]
- Coutu, A.N.; Lee-Thorp, J.; Collins, M.J.; Lane, P.J. Mapping the elephants of the 19th century East African ivory trade with a multi-isotope approach. PLoS ONE 2016, 11, e0163606. [Google Scholar] [CrossRef]
- Coutu, A.N.; Whitelaw, G.; le Roux, P.; Sealy, J. Earliest evidence for the ivory trade in Southern Africa: Isotopic and ZooMS analysis of seventh–tenth century AD ivory from KwaZulu-Natal. Afr. Archaeol. Rev. 2016, 33, 411–435. [Google Scholar] [CrossRef]
- Morcatty, T.Q.; Bausch-Macedo, J.C.; Nekaris, K.A.; Ni, Q.; Durigan, C.C.; Svensson, M.S.; Nijman, V. Illegal trade in wild cats and its link to Chinese-led development in Central and South America. Conserv. Biol. 2020, 34, 1525–1535. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, A.M.; Aliaga-Rossel, E. Jaguar fangs trafficking by Chinese in Bolivia. CAT News 2017, 65, 50–51. [Google Scholar]
- Arias, M.; Hinsley, A.; Nogales-Ascarrunz, P.; Negroes, N.; Glikman, J.A.; Milner-Gulland, E.J. Prevalence and characteristics of illegal jaguar trade in north-western Bolivia. Conserv. Sci. Pract. 2021, 3, e444. [Google Scholar] [CrossRef]
- Charity, S.; Ferreira, M. Wildlife trade in the Brazilian Amazon. In Wildlife Trafficking in Brazil; Charity, S., Ferreira, M., Eds.; TRAFFIC International: Cambridge, UK, 2020; pp. 59–93. [Google Scholar]
- Munro, R.; Munro, H.M.C. (Eds.) Estimation of time since death. In Animal Abuse and Unlawful Killing: Forensic Veterinary Pathology; Saunders Elsevier: London, UK, 2008; pp. 88–93. [Google Scholar]
- Brooks, J.W. Postmortem changes in animal carcasses and estimation of the postmortem interval. Vet. Pathol. 2016, 53, 929–940. [Google Scholar] [CrossRef]
- Brooks, J.W.; Sutton, L. Postmortem changes and estimating the postmortem interval. In Veterinary Forensic Pathology; Brooks, J.W., Ed.; Springer: Cham, Switzerland, 2018; pp. 43–63. [Google Scholar]
- Erlandsson, M.; Munro, R. Estimation of the post-mortem interval in beagle dogs. Sci. Justice 2007, 47, 150–154. [Google Scholar] [CrossRef]
- Brooks, J.W. Postmortem changes and the estimation of time since death. In Veterinary Forensics: Investigation, Evidence Collection, and Expert Testimony; Rogers, E., Stern, A.W., Eds.; CRC Press: Boca Raton, FL, USA, 2018; pp. 225–250. [Google Scholar]
- Merck, M.D.; Miller, D.M. Postmortem changes and the postmortem interval. In Veterinary Forensics: Animal Cruelty Investigations, 2nd ed.; Merck, M.D., Ed.; Wiley-Blackwell: Chichester, UK, 2013; pp. 255–286. [Google Scholar]
- Probst, C.; Gethmann, J.; Amendt, J.; Lutz, L.; Teifke, J.P.; Conraths, F.J. Estimating the postmortem interval of wild boar carcasses. Vet. Sci. 2020, 7, 6. [Google Scholar] [CrossRef]
- Ugrappa, S.; Jain, A. An emergence of dental tissues in the forensic medicine for the postmortem interval estimation: A scoping review. J. Forensic Sci. Med. 2021, 7, 54–60. [Google Scholar] [CrossRef]
- Akbulut, N.; Çetin, S.; Bilecenoğlu, B.; Altan, A.; Akbulut, S.; Ocak, M.; Orhan, K. The micro-CT evaluation of enamel-cement thickness, abrasion, and mineral density in teeth in the postmortem interval (PMI): New parameters for the determination of PMI. Int. J. Leg. Med. 2020, 134, 645–653. [Google Scholar] [CrossRef]
- Granrud, M.A.; Dabbs, G.R. A preliminary study of incisor exfoliation as an estimator of the postmortem interval using accumulated degree days. Forensic Sci. Int. 2012, 220, e29–e32. [Google Scholar] [CrossRef]
- Mehendiratta, M.; Jain, K.; Boaz, K.; Bansal, M.; Manaktala, N. Estimation of time elapsed since the death from identification of morphological and histological time-related changes in dental pulp: An observational study from porcine teeth. J. Forensic Dent. Sci. 2015, 7, 95–100. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Young, S.T.; Wells, J.D.; Hobbs, G.R.; Bishop, C.P. Estimating postmortem interval using RNA degradation and morphological changes in tooth pulp. Forensic Sci. Int. 2013, 229, 163.e1–163.e6. [Google Scholar] [CrossRef] [PubMed]
- Schulz, I.; Schneider, P.M.; Olek, K.; Rothschild, M.A.; Tsokos, M. Examination of postmortem animal interference to human remains using cross-species multiplex PCR. Forensic Sci. Med. Pathol. 2006, 2, 95–101. [Google Scholar] [CrossRef]
- Ubelaker, D.H.; DeGaglia, C.M. The impact of scavenging: Perspective from casework in forensic anthropology. Forensic Sci. Res. 2020, 5, 32–37. [Google Scholar] [CrossRef]
- Bernstein, M. The nature of bitemarks. In Bitemark Evidence: A Color Atlas and Text, 2nd ed.; Dorion, R., Ed.; CRC Press: Boca Raton, FL, USA, 2011; pp. 53–65. [Google Scholar]
- Binford, L.R. Bones: Ancient Men and Modern Myths; Academic Press: New York, NY, USA, 1981. [Google Scholar]
- Tanga, C. Gli Inumati di San Pelino e Sant’Alessandro, Corfinio (AQ): Osteobiografia dei Resti Umani. Bachelor’s Thesis, ‘G. d’Annunzio’ of Chieti-Pescara, Chieti, Italy, December 2016. [Google Scholar]
- Moraitis, K.; Spiliopoulou, C. Forensic implications of carnivore scavenging on human remains recovered from outdoor locations in Greece. J. Forensic Leg. Med. 2010, 17, 298–303. [Google Scholar] [CrossRef]
- Tsokos, M.; Schulz, F. Indoor postmortem animal interference by carnivores and rodents: Report of two cases and review of the literature. Int. J. Leg. Med. 1999, 112, 115–119. [Google Scholar] [CrossRef]
- Keyes, C.A.; Myburgh, J.; Brits, D. Taphonomic bone trauma caused by southern African scavengers. Int. J. Leg. Med. 2020, 134, 1227–1238. [Google Scholar] [CrossRef]
- Colard, T.; Delannoy, Y.; Naji, S.; Gosset, D.; Hartnett, K.; Bécart, A. Specific patterns of canine scavenging in indoor settings. J. Forensic Sci. 2015, 60, 495–500. [Google Scholar] [CrossRef]
- Young, A.; Márquez-Grant, N.; Stillman, R.; Smith, M.J.; Korstjens, A.H. An investigation of red fox (Vulpes vulpes) and Eurasian badger (Meles meles) scavenging, scattering, and removal of deer Remains: Forensic implications and applications. J. Forensic Sci. 2015, 60, S39–S55. [Google Scholar] [CrossRef]
- Young, A.; Stillman, R.; Smith, M.J.; Korstjens, A.H. Scavenger species-typical alteration to bone: Using bite mark dimensions to identify scavengers. J. Forensic Sci. 2015, 60, 1426–1435. [Google Scholar] [CrossRef]
- Young, A.; Stillman, R.; Smith, M.J.; Korstjens, A.H. Applying knowledge of species-typical scavenging behavior to the search and recovery of mammalian skeletal remains. J. Forensic Sci. 2016, 61, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Bellis, C.; Ashton, K.J.; Freney, L.; Blair, B.; Griffiths, L.R. A molecular genetic approach for forensic animal species identification. Forensic Sci. Int. 2003, 134, 99–108. [Google Scholar] [CrossRef]
- Jones, A. Animal Scavengers as Agents of Decomposition: The Postmortem Succession of Louisiana Wildlife. Master’s Thesis, Louisiana State University and Agricultural and Mechanical College, Baton Rouge, LA, USA, 2011. [Google Scholar]
- Mansegosa, D.A.; Marchiori, J.I.; Giannotti, P.S. Dislocation, consumption and marks in human bodies produced by carnivores: A comparated study with forensic cases of the central-west of Argentina. J. Forensic Anthropol. Odont. 2020, 3, 6–17. [Google Scholar]
- Steadman, D.W.; Dautartas, A.; Kenyhercz, M.W.; Jantz, L.M.; Mundorff, A.; Vidoli, G.M. Differential scavenging among pig, rabbit, and human subjects. J. Forensic Sci. 2018, 63, 1684–1691. [Google Scholar] [CrossRef]
- Krenzer, U. Compendio de Métodos Antropológico Forenses Para la Reconstrucción del Perfil Osteo-Biológico; CAFCA, Centro de Análisis Forense y Ciencias Aplicadas: Ciudad de Guatemala, Guatemala, 2006; Volume 7. [Google Scholar]
- Cantu, M. Animal Scavenging on Human Skeletal Remains in the Southwest United States: A Preliminary Model. Master’s Thesis, Louisiana State University and Agricultural and Mechanical College, Baton Rouge, LA, USA, 2014. [Google Scholar]
- Erkol, Z.; Hösükler, E. Postmortem animal attacks on human corpses. In Post Mortem Examination and Autopsy: Current Issues from Death to Laboratory Analysis; Dogan, K.H., Ed.; InTech: London, UK, 2018. [Google Scholar]
- Andrés, M.; Gidna, A.O.; Yravedra, J.; Domínguez-Rodrigo, M. A study of dimensional differences of tooth Marks (pits and scores) on bones modified by small and large carnivores. Archaeol. Anthropol. Sci. 2012, 4, 209–219. [Google Scholar] [CrossRef]
- Klippel, W.E.; Synstelien, J.A. Rodents as taphonomic agents: Bone gnawing by brown rats and gray squirrels. J. Forensic Sci. 2007, 52, 765–773. [Google Scholar] [CrossRef]
- Tsokos, M.; Matschke, J.; Gehl, A.; Koops, E.; Püschel, K. Skin and soft tissue artifacts due to postmortem damage caused by rodents. Forensic Sci. Int. 1999, 104, 47–57. [Google Scholar] [CrossRef]
- Brothwell, D. Further evidence of bone chewing by ungulates: The sheep of North Ronaldsay, Orkney. J. Archaeol. Sci. 1976, 3, 179–182. [Google Scholar] [CrossRef]
- Kierdorf, U. Fork formation and other signs of osteophagia on a long bone swallowed by a red deer stag (Cervus elaphus). Int. J. Osteoarchaeol. 1993, 3, 37–40. [Google Scholar] [CrossRef]
- Meckel, L.A.; McDaneld, C.P.; Wescott, D.J. White-tailed deer as a taphonomic agent: Photographic evidence of white-tailed deer gnawing on human Bone. J. Forensic Sci. 2018, 63, 292–294. [Google Scholar] [CrossRef]
- Tumram, N.K.; Ambade, V.N.; Dixit, P.G. Human fatalities caused by animal attacks: A six-year autopsy study. Med. Leg. J. 2017, 85, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Kiuchi, T.; Matoba, K.; Terazawa, K. Unusual injuries on the right hand and forearm caused by unidentified wild animals. Leg. Med. 2008, 10, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Byard, R.W.; James, R.A.; Gilbert, J.D. Diagnostic problems associated with cadaveric trauma from animal activity. Am. J. Forensic Med. Pathol. 2002, 23, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, G.; Mora, E.; Lucena, J.; Cantin, M. Forensic studies of dog attacks on humans: A focus on bite mark analysis. Res. Rep. Forensic Med. Sci. 2015, 5, 39–51. [Google Scholar] [CrossRef]
- De Munnynck, K.; Van de Voorde, W. Forensic approach of fatal dog attacks: A case report and literature review. Int. J. Leg. Med. 2002, 116, 295–300. [Google Scholar] [CrossRef]
- Pomara, C.; D’Errico, S.; Jarussi, V.; Turillazzi, E.; Fineschi, V. Cave canem: Bite mark analysis in a fatal dog pack attack. Am. J. Forensic Med. Pathol. 2011, 32, 50–54. [Google Scholar] [CrossRef]
- Haj Salem, N.; Belhadj, M.; Aissaoui, A.; Mesrati, M.A.; Chadly, A. Multidisciplinary approach to fatal dog attacks: A forensic case study. J. Forensic Leg. Med. 2013, 20, 763–766. [Google Scholar] [CrossRef]
- Le Garff, E.; Mesli, V.; Delannoy, Y.; Pollard, J.; Becart, A.; Hedouin, V. Domestic predation of an elder: A fatal dog attack case. J. Forensic Sci. 2017, 62, 1379–1382. [Google Scholar] [CrossRef]
- Santoro, V.; Smaldone, G.; Lozito, P.; Smaldone, M.; Introna, F. A forensic approach to fatal dog attacks. A case study and review of the literature. Forensic Sci. Int. 2011, 206, e37–e42. [Google Scholar] [CrossRef]
- Marko, P.B. Sympatry. In Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; Academic Press: Amsterdam, The Netherlands, 2008; pp. 3450–3458. [Google Scholar]
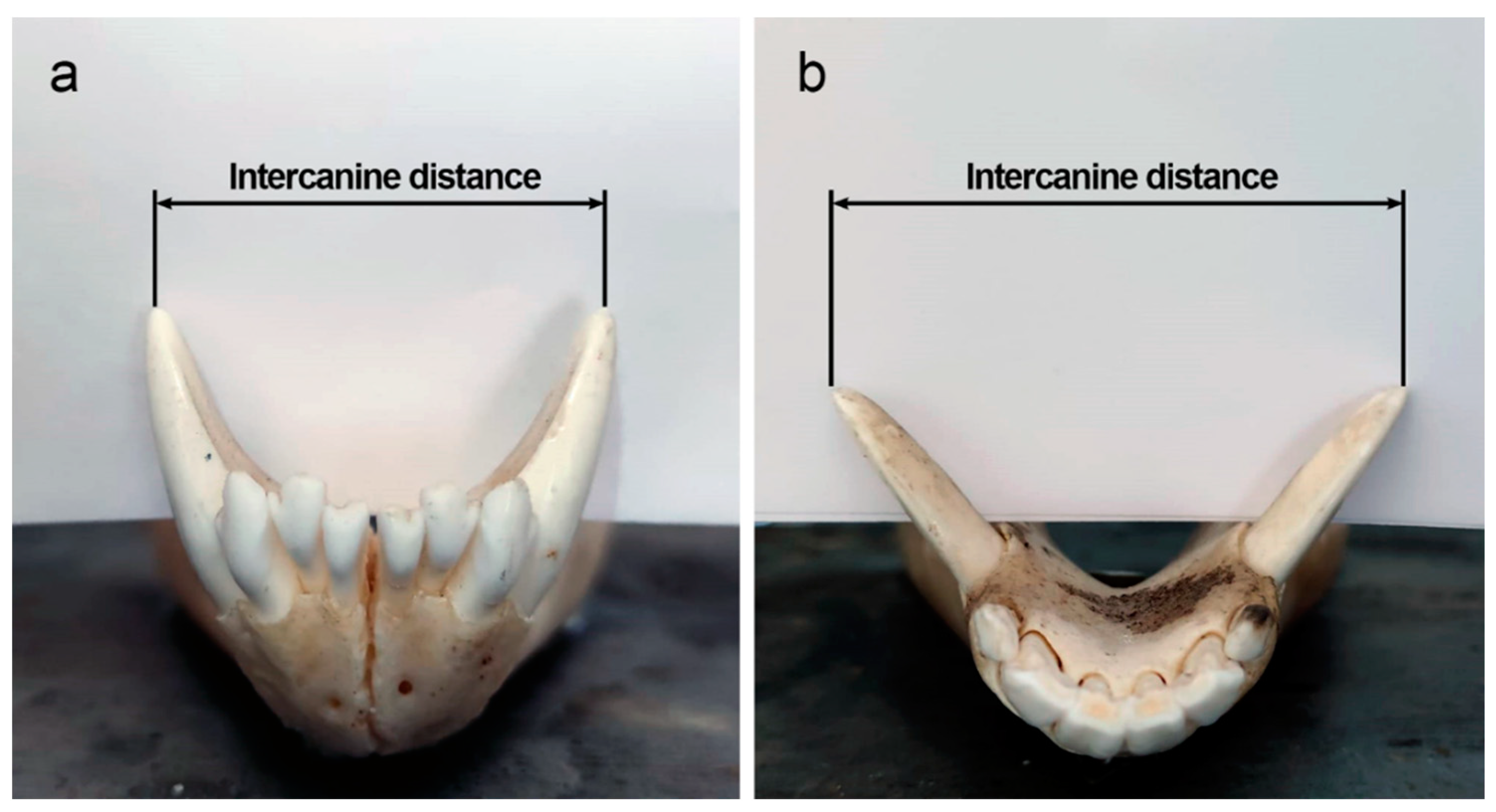
- Ohana, A.J.S. Intercanine Distance Used as a Measure to Rule Out Biters in Bite Mark Forensics. Master’s Thesis, University of Tennessee: Knoxville, TN, USA, 2016. [Google Scholar]
- Põdra, M.; Gómez, A.; Palazón, S. Do American mink kill European mink? Cautionary message for future recovery efforts. Eur. J. Wildl. Res. 2013, 59, 431–440. [Google Scholar] [CrossRef]
- Verzuh, T.; Bergman, D.L.; Bender, S.C.; Dwire, M.; Breck, S.W. Intercanine width measurements to aid predation investigations: A comparison between sympatric native and non-native carnivores in the Mexican wolf recovery area. J. Mammal. 2018, 99, 1405–1410. [Google Scholar] [CrossRef]
- Ratz, H.; Moller, H. Identification of Predators of Royal Albatross Chicks at Taiaroa Head in February 1994; Conservation Advisory Science Notes No. 138; Department of Conservation: Wellington, New Zealand, 1997. [Google Scholar]
- Elbroch, M.; McFarland, C. (Eds.) Interpreting prey remains. In Mammal Tracks and Sign: A Guide to North American Species, 2nd ed.; Stackpole Books: Guilford, CT, USA, 2019; pp. 307–376. [Google Scholar]
- Knüsel, C.; Robb, J. Funerary taphonomy: An overview of goals and methods. J. Archaeol. Sci. Rep. 2016, 10, 655–673. [Google Scholar] [CrossRef]
- Cyriac, V.P.; Kodandaramaiah, U. Digging their own macroevolutionary grave: Fossoriality as an evolutionary dead end in snakes. J. Evol. Biol. 2018, 31, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Morlan, R.E. Dialogue: Rodent bones in archaeological sites. J. Can. D’archéologie 1994, 18, 135–142. [Google Scholar]
- Camarós, E.; Cueto, M.; Lorenzo, C.; Villaverde, V.; Rivals, F. Large carnivore attacks on hominins during the Pleistocene: A forensic approach with a Neanderthal example. Archaeol. Anthropol. Sci. 2016, 8, 635–646. [Google Scholar] [CrossRef]
- Wolverton, S.; Nagaoka, L. Zooarcheology: Investigating past interactions between humans and other Animals. In Ethnozoology: Animals in Our Lives; Nóbrega-Alves, R.R., Albuquerque, U.P., Eds.; Academic Press: London, UK, 2018; pp. 25–43. [Google Scholar] [CrossRef]
- Domínguez-Rodrigo, M.; Piqueras, A. The use of tooth pits to identify carnivore taxa in tooth-marked archaeofaunas and their relevance to reconstruct hominid carcass processing behaviours. J. Archaeol. Sci. 2003, 30, 1385–1391. [Google Scholar] [CrossRef]
- Selvaggio, M.M.; Wilder, J. Identifying the involvement of multiple carnivore taxa with archaeological bone assemblages. J. Archaeol. Sci. 2001, 28, 465–470. [Google Scholar] [CrossRef]
- Brain, C.K. New Finds at the Swartkrans Australopithecine Site. Nature 1970, 225, 1112–1119. [Google Scholar] [CrossRef]
- Pobiner, B. Paleoecological information in predator tooth marks. J. Taphon. 2008, 6, 373–397. [Google Scholar]
- Haynes, G. Evidence of carnivore gnawing on Pleistocene and recent mammalian bones. Paleobiology 1980, 6, 341–351. [Google Scholar] [CrossRef]
- Haynes, G. A guide for differentiating mammalian carnivore taxa responsible for gnaw damage to herbivore limb bones. Paleobiology 1983, 9, 164–172. [Google Scholar] [CrossRef]
- Pante, M.C.; Blumenschine, R.J.; Capaldo, S.D.; Scott, R.S. Validation of bone surface modification models for inferring fossil hominin and carnivore feeding interactions, with reapplication to FLK 22, Olduvai Gorge, Tanzania. J. Hum. Evol. 2012, 63, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Egeland, C.P.; Domínguez-Rodrigo, M. Taphonomic perspectives on hominid site use and foraging strategies during Bed II times at Olduvai Gorge, Tanzania. J. Hum. Evol. 2008, 55, 1031–1052. [Google Scholar] [CrossRef]
- Njau, J.K.; Blumenschine, R.J. Crocodylian and mammalian carnivore feeding traces on hominid fossils from FLK 22 and FLK NN 3, Plio-Pleistocene, Olduvai Gorge, Tanzania. J. Hum. Evol. 2012, 63, 408–417. [Google Scholar] [CrossRef]
- Daujeard, C.; Geraads, D.; Gallotti, R.; Lefèvre, D.; Mohib, A.; Raynal, J.-P.; Hublin, J.-J. Pleistocene hominins as a resource for carnivores: A c. 500,000-year-old human femur bearing tooth-marks in North Africa (Thomas Quarry I, Morocco). PLoS ONE 2016, 11, e0152284. [Google Scholar] [CrossRef] [PubMed]
- Holloway, R.L.; Broadfield, D.C.; Yuan, M.S. The Human Fossil Record; Wiley-Liss: Hoboken, NJ, USA, 2004; Volume 3. [Google Scholar]
- Singh, V.; Shukla, S.; Sing, A. The principal factors responsible for biodiversity loss. Open J. Plant. Sci. 2021, 6, 11–14. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Fluids and Tissues | Time Period Represented | ||||
|---|---|---|---|---|---|
| Hours | Days | Weeks | Months | Years | |
| Urine | ✓ | ✓ | |||
| Faeces | ✓ | ✓ | |||
| Blood plasma | ✓ | ✓ | |||
| Blood cells | ✓ | ✓ | |||
| Hair | ✓ | ✓ | ✓ | ✓ * | |
| Nail | ✓ | ✓ | ✓ | ✓ * | |
| Claw | ✓ | ✓ | ✓ | ✓ * | |
| Bill | ✓ | ✓ | ✓ | ||
| Feather | ✓ | ✓ | ✓ | ||
| Antler | ✓ | ✓ | ✓ | ||
| Muscle | ✓ | ✓ | ✓ | ||
| Bone (collagen) | ✓ | ||||
| Tooth (dentine, enamel) | ✓ | ||||
| Family | Species | Common Name | Intercanine Distance (mm) | |
|---|---|---|---|---|
| Maxilla | Mandible | |||
| Felidae | Felis catus | Domestic cat | 7.0–22.0 | 4.0–18.0 |
| Felis catus | Feral cat | 8.2–21.0 * | ||
| Lynx canadensis | Canada lynx | 22.7–27.6 | 21.9–27.8 | |
| Lynx lynx | Eurasian lynx | 13.0–30.0 | 5.0–25.0 | |
| Lynx rufus | Bobcat | 11.0–31.0 | 5.0–24.0 | |
| Puma concolor | Cougar | 21.0–48.0 | 10.0–44.0 | |
| Canidae | Canis latrans | Coyote | 15.0–39.1 | 7.0–39.0 |
| Canis lupus | Grey wolf | 23.0–51.0 | 11.0–45.0 | |
| Canis lupus baileyi | Mexican wolf | 31.4–49.8 | 27.8–43.7 | |
| Canis lupus familiaris | Domestic dog | 13.0–48.0 | 6.0–49.0 | |
| Canis lupus familiaris | Feral dog | 14.1–53.4 | 10.8–46.2 | |
| Urocyon cinereoargenteus | Gray fox | 9.0–22.0 | 4.0–20.0 | |
| Vulpes lagopus | Arctic fox | 19.4–24.2 | 16.4–20.8 | |
| Vulpes macrotis | Kit fox | 13.8–17.5 | 11.3–15.5 | |
| Vulpes velox | Swift fox | 15.0–17.0 | 13.6–15.2 | |
| Vulpes vulpes | Red fox | 11.0–27.0 | 4.0–25.0 | |
| Mephitidae | Mephitis mephitis | Striped skunk | 12.0–16.4 | 10.3–14.8 |
| Spilogale gracilis | Western spotted skunk | 8.5–11.6 | 7.5–10.5 | |
| Mustelidae | Genetta genetta | Spotted genet | 9.0–13.0 * | |
| Gulo gulo | Wolverine | 18.0–43.0 | 7.0–32.0 | |
| Lontra canadensis | Northern river otter | 16.7–22.6 | 14.6–21.4 | |
| Martes americanus | American marten | 8.6–14.6 | 8.2–12.5 | |
| Martes foina | Stone marten | 11.0–15.0 * | ||
| Mustela erminea | Stoat | 3.5–7.1 | 2.6–5.1 | |
| Mustela frenata | Long-tailed weasel | 5.8–9.0 | 4.6–8.0 | |
| Mustela furo | Ferret | 7.8–13.6 * | ||
| Mustela lutreola | European mink | 8.0–12.0 * | ||
| Mustela nivalis | Least weasel | 3.2–5.3 | 2.7–4.4 | |
| Mustela putorius | Polecat | 8.0–14.0 * | ||
| Neogale vison | American mink | 8.3–12.4 | 6.2–10.1 | |
| Taxidea taxus | American badger | 24.8–33.9 | 23.8–32.3 | |
| Phocidae | Phoca vitulina | Harbour seal | 27.5–36.3 | 23.0–36.1 |
| Procyonidae | Bassariscus astutus | Ringtail | 9.5–12.2 | 8.6–11.8 |
| Nasua narica | White-nosed coati | 19.1–29.3 | 16.4–22.1 | |
| Procyon lotor | Raccoon | 19.4–28.2 | 16.1–25.6 | |
| Ursidae | Ursus americanus | Black bear | 20.0–64.0 | 11.0–52.0 |
| Ursus arctos horribilis | Grizzly bear | 34.0–96.0 | 15.0–91.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viciano, J.; López-Lázaro, S.; Tanga, C. Post-Mortem Dental Profile as a Powerful Tool in Animal Forensic Investigations—A Review. Animals 2022, 12, 2038. https://doi.org/10.3390/ani12162038
Viciano J, López-Lázaro S, Tanga C. Post-Mortem Dental Profile as a Powerful Tool in Animal Forensic Investigations—A Review. Animals. 2022; 12(16):2038. https://doi.org/10.3390/ani12162038
Chicago/Turabian StyleViciano, Joan, Sandra López-Lázaro, and Carmen Tanga. 2022. "Post-Mortem Dental Profile as a Powerful Tool in Animal Forensic Investigations—A Review" Animals 12, no. 16: 2038. https://doi.org/10.3390/ani12162038
APA StyleViciano, J., López-Lázaro, S., & Tanga, C. (2022). Post-Mortem Dental Profile as a Powerful Tool in Animal Forensic Investigations—A Review. Animals, 12(16), 2038. https://doi.org/10.3390/ani12162038