1. Introduction
Reducing the levels of dietary crude protein (CP) has been traditionally considered to favor the reduction in feed cost, nitrogen excretion, and the risk of gut disorders, without impairing the growth performance of pigs. Interestingly, besides these beneficial effects, accumulating evidence also shows that low-protein diets greatly increase the intramuscular fat content and improve meat quality [
1,
2,
3]. In this sense, studies on the application of low-protein diets in animal production have been gaining attention. In the literature, information on the application of low-protein diets in non-Chinese commercial breeds is available. However, the effects of low-protein diets on the meat quality of native Chinese pig breeds are less well-known.
Non-Chinese commercial breeds and native Chinese pig breeds display distinctive differences in terms of meat quality, prolificacy, and growth. Specifically, native Chinese pig breeds have better meat quality (such as the cherry-red color, high levels of marbling, soft texture and superior flavor), lower growth rates, and lower lean meat rates when compared with non-Chinese commercial breeds [
4,
5,
6]. With the improvement in people’s living standards, a growing demand for healthier, tastier, and more nutritious pork has emerged. In this context, native Chinese pig breeds are returning as focus of studies, mainly since they are a resource that could meet the diverse needs of consumers [
7].
The Shaziling pig is a traditional fat-type native pig breed mainly reared in Hunan Province, China. It possesses excellent meat quality, strong disease resistance, and slow growth rates like most native Chinese pig breeds [
8]. However, studies of Shaziling pigs are very limited, and previous studies have mainly focused on the comparison of carcass traits, meat quality, serum metabolome, and lipid metabolic and microbial profiles between Shaziling and Yorkshire pigs at different ages [
9,
10], and have discussed the differences in genetics, involving epigenetics, quantitative genetics, and metagenomics between Shaziling and Yorkshire pigs [
8,
11]. In view of the information above, our understanding of optimal nutrition interventions to regulate meat quality of Shaziling pigs is at its infancy, which hinders the breeding of Shaziling pigs and the promotion of meat products. Given the importance of nutrition in the quality control of animal meat, further research is required.
Metabolomics is a method that has the ability to detect subtle alterations in the content of low-molecular-weight metabolites in complex biological systems such as tissues and cells, resulting from external or internal factors. The past few decades have witnessed the development of metabolomics, and this emerging analytical platform shows valuable application in assessing meat quality. For example, ultra-high performance liquid chromatography–tandem mass spectrometry (UHPLC-MS/MS) aided in the investigation of serum potential biomarkers to predict meat quality of Shaziling pigs [
10]. Non-targeted liquid chromatography–mass spectrometry has been used to characterize lipid-related metabolites in the muscle of pigs [
12]. Xiao et al. adopted
1H nuclear magnetic resonance spectroscopy to analyze the metabolic composition of chicken meat [
13]. Ultra-performance liquid chromatography–tandem mass spectrometry (UPLC-MS/MS) has been used to investigate the underlying mechanisms responsible for chlorogenic acid-induced improvements in meat quality of broiler breast muscles [
14]. These studies highlighted the effectiveness of metabolomics for the evaluation of meat quality; hence, an approach integrating metabolomics and molecular biology should advance the understanding of the mechanism of meat quality changes in response to low-protein diets.
In this study, we determined the effects of long-term protein restriction for 24 weeks on meat quality of Shaziling pigs and showed muscle metabolite alterations resulting from low-protein diets via analysis of muscular metabolic profiling.
4. Discussion
The role of protein-restriction diets in saving protein resources and decreasing the emission of nitrogen in urea and feces has been well characterized. However, most of the studies related to protein restriction have focused on the short-term effects on non-Chinese commercial breeds [
19]. Moreover, our understanding of the long-term effects of high-protein and low-protein diets on pig production performance (growth performance, carcass characteristics, and meat quality) is incomplete for native Chinese pig breeds. The long-term use of low-protein diet to save protein feed resources cannot be at the expense of pig production performance. The first factor to evaluate low-protein diets is growth performance, which includes ADG, ADFI, and F:G. Decades of studies have well-demonstrated that dietary CP reduction within 3% of the NRC (1998) did not impair the growth performance of growing–finishing pigs when supplemented with the first four limited amino acids (L-lysine, DL-methionine, L-threonine and L-tryptophan) [
20]. However, an inhibitory effect on growth performance was observed when dietary CP levels were reduced by more than 3% with only the first four limited amino acids supplemented in diets [
21]. Further studies have elucidated that compared with pigs fed high-protein diets, a reduction in CP by 4.8% along with the first four limited amino acids supplemented led to significantly reduced growth performance [
22]. However, conflicting results were observed in the current study, which showed that reducing dietary CP level by 20% relative to the control group for 24 weeks resulted in similar growth performance in Shaziling pigs as those of control diets and long-term high-protein diets. These above findings suggest that long-term protein restriction (20% reduction in comparison to the control group) could be applied in Shaziling pigs without affecting their growth performance.
Carcass characteristics, including carcass length, carcass weight, dressing percentage, backfat thickness, loin eye area, lean percentage, and fat percentage, is the second factor to assess low-protein diets. Extensive past research has demonstrated that reducing dietary CP levels did not significantly affect dressing percentage [
23,
24,
25]. These results are well-matched with our research since Shaziling pigs fed protein-restricted diets for 24 weeks had similar dressing percentage as that of control and high-protein diets. However, decreased loin eye area and increased backfat thickness at slaughter were consistently reported in pigs fed low-protein diets [
26,
27,
28,
29,
30]. However, contradictory results are found when looking at long-term protein-restricted diets, which had no detrimental effect on loin eye area and backfat thickness of Shaziling pigs. This discrepancy might be related to response variation in different breeds (non-Chinese commercial breeds in previous studies vs. native Chinese pig breeds in this study), and these controversial data point out the necessity to further explore the mechanisms underlying these effects of long-term protein restriction on Shaziling pigs. These findings suggest a balance of energy to nitrogen in diets of the current study.
Meat quality, the third factor to evaluate low-protein diets, is primarily assessed by the following parameters: pH, color, WHC (drip loss, pressing loss, and cooking loss), and tenderness (shear force) [
31]. Accumulating and emerging lines of evidence have revealed no significant influence of altering dietary CP on pH
24h and WHC of pigs [
32,
33]. In agreement with this, we found no significant difference in meat values of pH
45min, pH
24h, and WHC in Shaziling pigs fed different dietary CP. However, contradictory results are reported when looking at meat color. Several laboratory established that the values of L*, a*, and b* elevated when dietary CP was restricted [
26,
32]. In contrast, other studies demonstrated that altering dietary CP did not significantly affect the L* value [
33,
34]. The current study reports a significant reduction in L* value and an increase in a* value in pigs fed with low-protein diets (20% reduction in comparison to the control group) during the stage from 8.78 to 83.50 kg body weight. This finding suggests that low-protein diets lead to an improvement on meat quality of Shaziling pigs.
Fatty acid composition is an important contributor to various aspects of meat quality and nutritional value of meat and also the human health [
35]. Studies have shown that SFA, such as C12:0, C14:0, C16:0, and C17:0, will raise the risk of cardiovascular disease and type 2 diabetes when over-consumed [
36,
37]. In this study, C12:0 and C17:0 greatly decreased when dietary protein was restricted by 20% relative to the control group. Therefore, the intake of Shaziling pigs fed protein-restricted diets (reducing dietary CP level by 20% relative to the control group) for 24 weeks may reduce the risk of cardiovascular disease. Conversely, unsaturated fatty acids, especially n3 PUFAs, are beneficial to human health and are also related to flavor and overall acceptability of meat [
38]. In the current study, the n3 PUFA decreased with the low-protein diets but did not achieve statistical significance. These data suggest that reducing dietary CP level by 20% relative to the control group for 24 weeks did not significantly impair the flavor and overall acceptability of Shaziling pig meat. In addition, a PUFA:SFA ratio of above 0.4 has been recommended for meat [
39], but in the present study, values were lower than this and were not significantly influenced by dietary CP levels. Moreover, higher values for Hpo/Hper and lower values for AI and TI are regarded to be healthier [
18]. Our current study showed that these parameters were not significantly different between the control and the −20% group. Overall, from the perspective of fatty acid composition, reducing dietary CP level by 20% relative to the control group for 24 weeks could produce healthy pork as the control group.
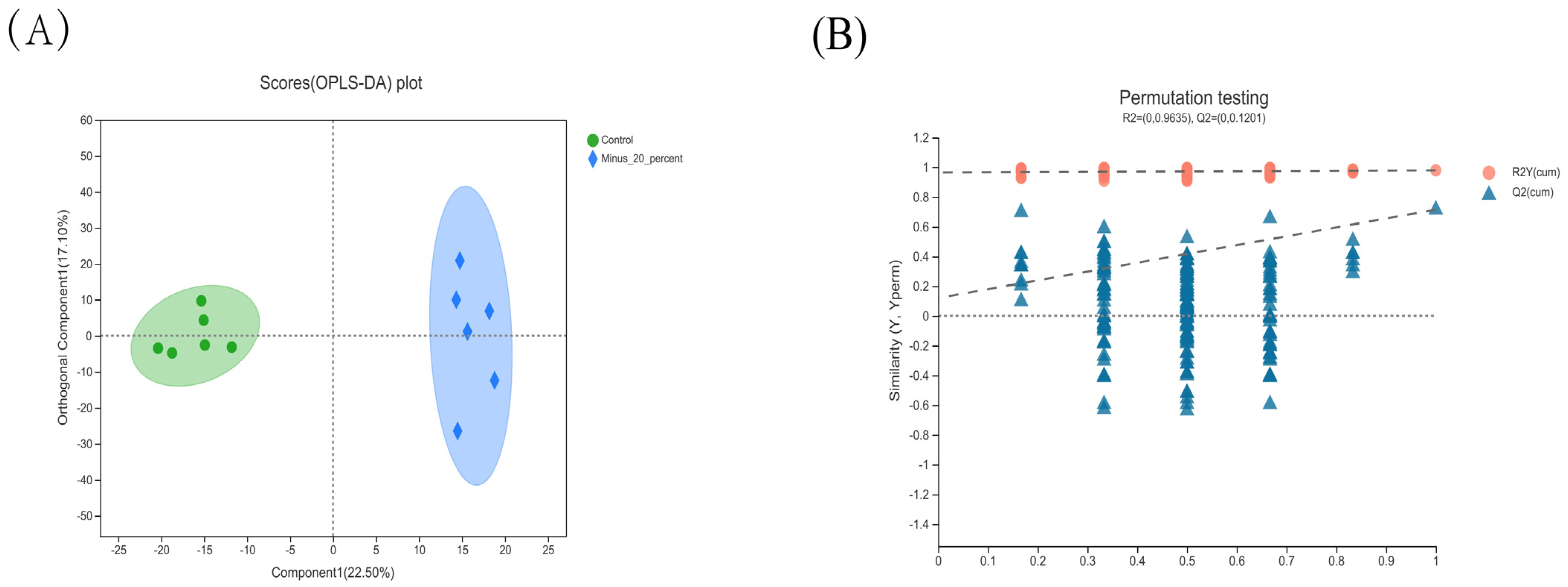
Metabolomics is applied for the investigation of key metabolites contributing to the physico-chemical properties, and hence, it helps to account for meat quality traits [
40]. In the current study, a comparative analysis of muscular metabolome was performed between the control and −20% group, since the −20% group improved meat quality without impairing growth performance and carcass characteristics. We found that the −20% group exhibited significantly lower concentrations of Danazol, N,N-dimethyl-Safingol, and cer(d18:0/14:0) compared with the control group. Moreover, the correlation analysis showed that the three metabolites were negatively associated with the a* value and positively related to the L* value. Therefore, based on these data and the abovementioned data concerning meat quality, it is postulated that Danazol, N,N-dimethyl-Safingol, and cer(d18:0/14:0) might be potential biomarkers of the −20% group and might be implicated in various pathways for improved meat quality upon protein-restricted diets.
In conclusion, the current study suggested that the long-term ingestion of a protein-restricted diet could improve the meat quality of Shaziling pigs without impairing their growth performance and carcass characteristics. Furthermore, long-term protein restriction reduced the metabolites (including Danazol, N,N-dimethyl-Safingol, and cer(d18:0/14:0)) in the muscle of Shaziling pigs, which may help to explain the improvement in meat quality. Taken together, the above findings provide a molecular basis for designing nutritional and effective feeding strategy for Shaziling pigs to improve meat quality and sustain their growth performance.


 ,
, 



{kind=link}
{kind=link}
{kind=link}