
Effects of Maternal Nutrient Restriction and Melatonin Supplementation on Cardiomyocyte Cell Development Parameters Using Machine Learning Techniques


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal and Experimental Design
2.2. Gestational Day 240 Measurements
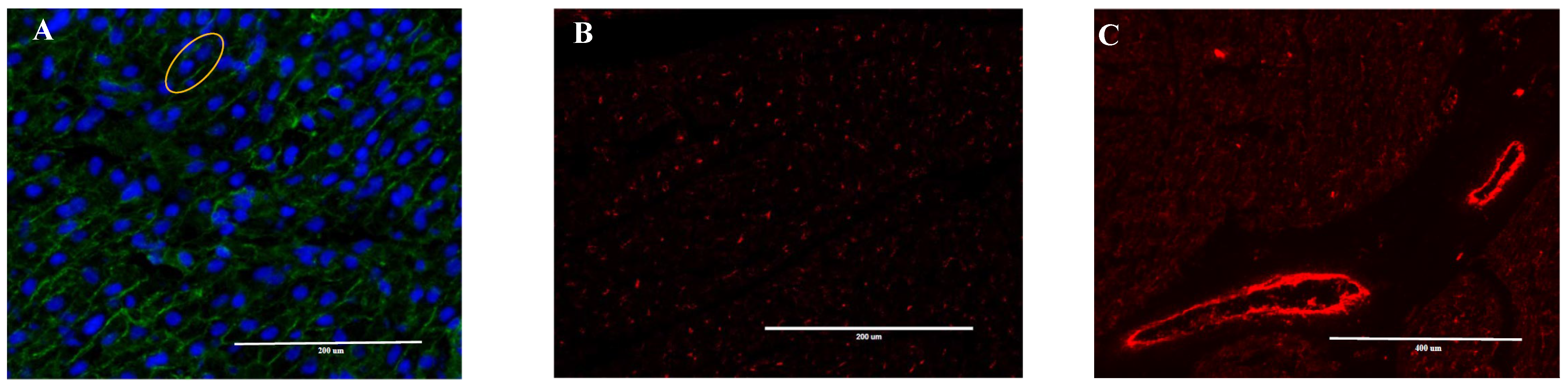
2.3. Heart Immunofluorescence Imaging
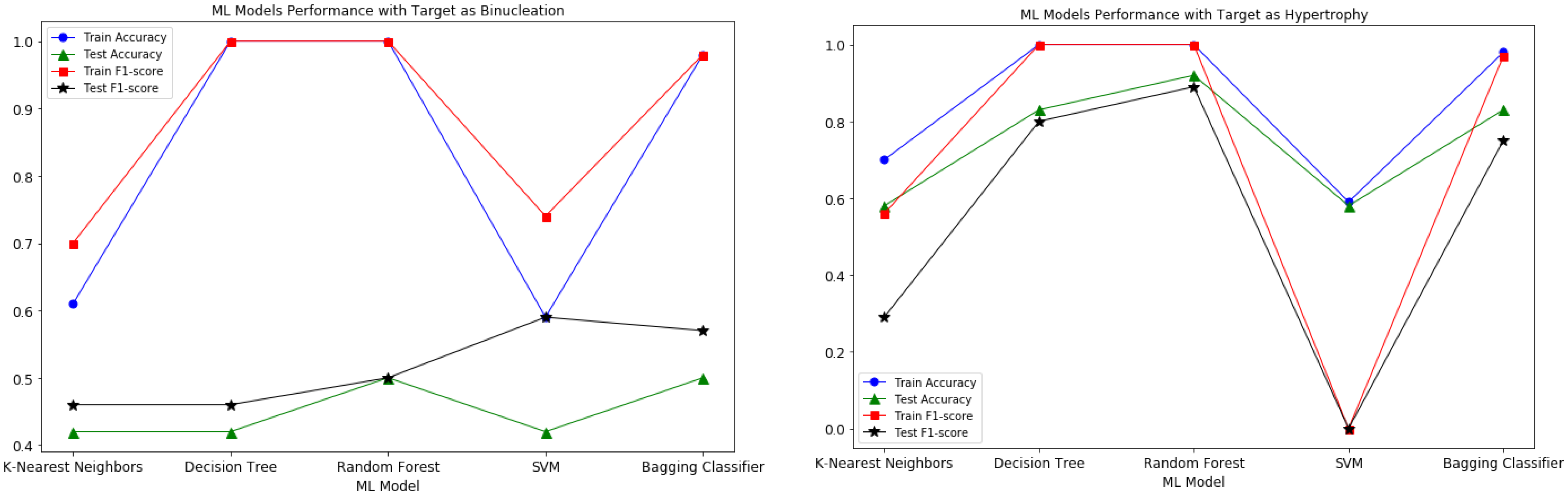
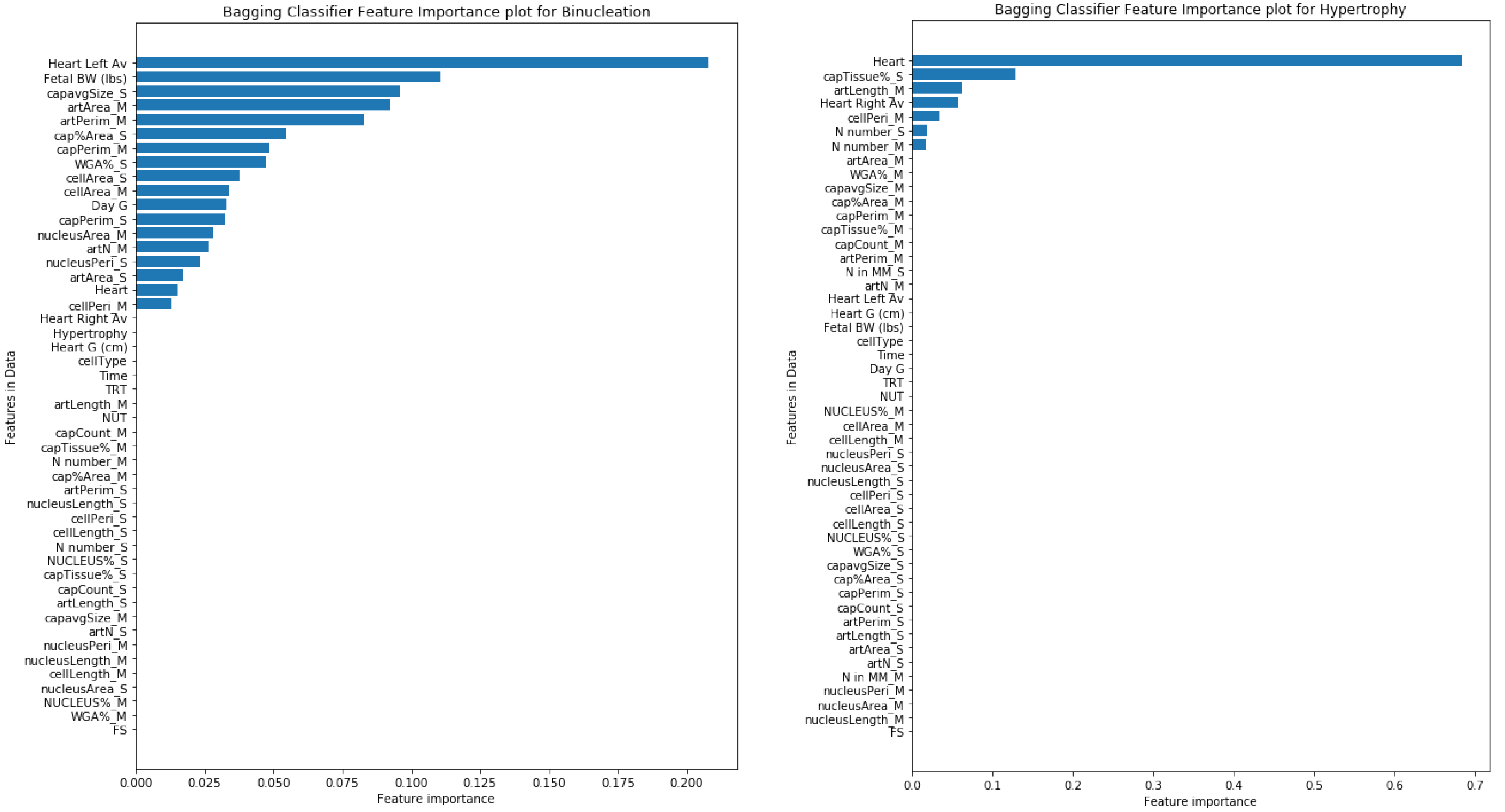
2.4. Training Machine Learning Models
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Godfrey, K.M.; Barker, D.J. Fetal nutrition and adult disease. Am. J. Clin. Nutr. 2000, 71, 1344S–1352S. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, K.M. The role of the placenta in fetal programming—A review. Placenta 2002, 23, 20–27. [Google Scholar] [CrossRef]
- Redmer, D.A.; Wallace, J.M.; Reynolds, L.P. Effect of nutrient intake during pregnancy on fetal and placental growth and vascular development. Domest. Anim. Endocrinol. 2004, 27, 199–217. [Google Scholar] [CrossRef] [PubMed]
- Kenny, D.; Murphy, C.P.; Sleator, R.D.; Judge, M.M.; Evans, R.D.; Berry, D.P. Animal-level factors associated with the achievement of desirable specifications in Irish beef carcasses graded using the EUROP classification system. J. Anim. Sci. 2020, 98, skaa191. [Google Scholar] [CrossRef] [PubMed]
- Ford, S.P.; Hess, B.W.; Schwope, M.M.; Nijland, M.J.; Gilbert, J.S.; Vonnahme, K.A.; Means, W.J.; Han, H.; Nathanielsz, P.W. Maternal undernutrition during early to mid-gestation in the ewe results in altered growth, adiposity, and glucose tolerance in male offspring1. J. Anim. Sci. 2007, 85, 1285–1294. [Google Scholar] [CrossRef] [PubMed]
- Sonberg, E. Gestational Weight Gain by Class of Obesity 2020. Master’s Thesis, University of Oslo, Oslo, Norway, 7 February 2020. [Google Scholar]
- Bensley, J.G.; De Matteo, R.; Harding, R.; Black, M.J. Three-dimensional direct measurement of cardiomyocyte volume, nuclearity, and ploidy in thick histological sections. Sci. Rep. 2016, 6, 23756. [Google Scholar] [CrossRef] [Green Version]
- Lemley, C.O.; Meyer, A.M.; Camacho, L.E.; Neville, T.L.; Newman, D.J.; Caton, J.S.; Vonnahme, K.A. Melatonin supplementation alters uteroplacental hemodynamics and fetal development in an ovine model of intrauterine growth restriction. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, 454–467. [Google Scholar] [CrossRef] [Green Version]
- Vonnahme, K.A.; Hess, B.W.; Hansen, T.R.; McCormick, R.J.; Rule, D.C.; Moss, G.E.; Murdoch, W.J.; Nijland, M.J.; Skinner, D.C.; Nathanielsz, P.W.; et al. Maternal Undernutrition from Early- to Mid-Gestation Leads to Growth Retardation, Cardiac Ventricular Hypertrophy, and Increased Liver Weight in the Fetal Sheep. Biol. Reprod. 2003, 69, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Giussani, D.A.; Davidge, S.T. Developmental programming of cardiovascular disease by prenatal hypoxia. J. Dev. Orig. Health Dis. 2013, 4, 328–337. [Google Scholar] [CrossRef]
- Rizzo, P.; Raffone, E.; Benedetto, V. Effect of the treatment with myo-inositol plus folic acid plus melatonin in comparison with a treatment with myo-inositol plus folic acid on oocyte quality and pregnancy outcome in IVF cycles. A prospective, clinical trial. Eur. Rev. Med. Pharmacol. Sci. 2010, 14, 555–561. [Google Scholar]
- Reiter, R.J.; Tan, D.X.; Galano, A. Melatonin: Exceeding Expectations. Physiology 2014, 29, 325–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voiculescu, S.E.; Zygouropoulos, N.; Zahiu, C.D.; Zagrean, A.M. Role of melatonin in embryo fetal development. J. Med. Life 2014, 7, 488–492. [Google Scholar] [PubMed]
- Unfer, V.; Raffone, E.; Rizzo, P.; Buffo, S. Effect of a supplementation with myo-inositol plus melatonin on oocyte quality in women who failed to conceive in previous in vitro fertilization cycles for poor oocyte quality: A prospective, longitudinal, cohort study. Gynecol. Endocrinol. 2011, 27, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Okatani, Y.; Okamoto, K.; Hayashi, K.; Wakatsuki, A.; Tamura, S.; Sagara, Y. Maternal-fetal transfer of melatonin in pregnant women near term. J. Pineal Res. 1998, 25, 129–134. [Google Scholar] [CrossRef]
- Kivelä, A.; Kauppila, A.; Leppäluoto, J.; Vakkuri, O. Serum and Amniotic Fluid Melatonin during Human Labor. J. Clin. Endocrinol. Metab. 1989, 69, 1065–1068. [Google Scholar] [CrossRef]
- Supramaniam, V.; Jenkin, G.; Loose, J.; Wallace, E.; Miller, S. Basic science: Chronic fetal hypoxia increases activin A concentrations in the late-pregnant sheep. BJOG An Int. J. Obstet. Gynaecol. 2005, 113, 102–109. [Google Scholar] [CrossRef]
- Liakos, K.G.; Busato, P.; Moshou, D.; Pearson, S.; Bochtis, D. Machine Learning in Agriculture: A Review. Sensors 2018, 18, 2674. [Google Scholar] [CrossRef] [Green Version]
- Valletta, J.J.; Torney, C.; Kings, M.; Thornton, A.; Madden, J. Applications of machine learning in animal behaviour studies. Anim. Behav. 2017, 124, 203–220. [Google Scholar] [CrossRef]
- Contreras-Correa, Z.E.; Messman, R.D.; Sidelinger, D.R.; Heath King, E.; Sánchez-Rodríguez, H.L.; Burnett, D.D.; Lemley, C.O. Melatonin alters bovine uterine artery hemodynamics, vaginal temperatures, and fetal morphometrics during late gestational nutrient restriction in a season-dependent manner. J. Anim. Sci. 2021, 99, skab242. [Google Scholar] [CrossRef]
- Brockus, K.E.; Hart, C.G.; Gilfeather, C.L.; Fleming, B.O.; Lemley, C.O. Dietary melatonin alters uterine artery hemodynamics in pregnant Holstein heifers. Domest. Anim. Endocrinol. 2016, 55, 1–10. [Google Scholar] [CrossRef]
- Vujic, A.; Lerchenmüller, C.; Di Wu, T.; Guillermier, C.; Rabolli, C.P.; Gonzalez, E.; Senyo, S.E.; Liu, X.; Guerquin-Kern, J.L.; Steinhauser, M.L.; et al. Exercise induces new cardiomyocyte generation in the adult mammalian heart. Nat. Commun. 2018, 9, 1659. [Google Scholar] [CrossRef]
- McCarty, K.J.; Owen, M.P.T.; Hart, C.G.; Thompson, R.C.; Burnett, D.D.; King, E.H.; Hopper, R.M.; Lemley, C.O. Effect of chronic melatonin supplementation during mid to late gestation on maternal uterine artery blood flow and subsequent development of male offspring in beef cattle. J. Anim. Sci. 2018, 96, 5100–5111. [Google Scholar] [CrossRef] [PubMed]
- Hemati, K.; Pourhanifeh, M.H.; Dehdashtian, E.; Fatemi, I.; Mehrzadi, S.; Reiter, R.J.; Hosseinzadeh, A. Melatonin and morphine: Potential beneficial effects of co-use. Fundam. Clin. Pharmacol. 2021, 35, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Golombek, D.A.; Pévet, P.; Cardinali, D.P. Melatonin Effects on Behavior: Possible Mediation by the Central GABAergic System. Neurosci. Biobehav. Rev. 1996, 20, 403–412. [Google Scholar] [CrossRef]
- Long, J.M.; Trubenbach, L.A.; Pryor, J.H.; Long, C.R.; Wickersham, T.A.; Sawyer, J.E.; Satterfield, M.C. Maternal nutrient restriction alters endocrine pancreas development in fetal heifers. Domest. Anim. Endocrinol. 2021, 74, 106580. [Google Scholar] [CrossRef] [PubMed]
- Broadhead, D.; Mulliniks, J.T.; Funston, R.N. Developmental Programming in a Beef Production System. Vet. Clin. N. Am. Food Anim. Pract. 2019, 35, 379–390. [Google Scholar] [CrossRef]
- Greenwood, P.L.; Cafe, L.M. Prenatal and pre-weaning growth and nutrition of cattle: Long-term consequences for beef production. Animal 2007, 1, 1283–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trubenbach, L.A.; Wickersham, T.A.; Bierschwale, L.N.; Morrill, J.C.; Baber, J.R.; Sawyer, J.E. Limit feeding as a strategy to increase energy efficiency in intensified cow–calf production systems1. Transl. Anim. Sci. 2019, 3, 796–810. [Google Scholar] [CrossRef] [Green Version]
- Hales, C.N.; Barker, D.J.P. Type 2 (non-insulin-dependent) diabetes mellitus: The thrifty phenotype hypothesis. Diabetologia 1992, 35, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Underwood, K.R.; Tong, J.F.; Price, P.L.; Roberts, A.J.; Grings, E.E.; Hess, B.W.; Means, W.J.; Du, M. Nutrition during mid to late gestation affects growth, adipose tissue deposition, and tenderness in cross-bred beef steers. Meat Sci. 2010, 86, 588–593. [Google Scholar] [CrossRef]
- Long, N.M.; Tousley, C.B.; Underwood, K.R.; Paisley, S.I.; Means, W.J.; Hess, B.W.; Du, M.; Ford, S.P. Effects of early- to mid-gestational undernutrition with or without protein supplementation on offspring growth, carcass characteristics, and adipocyte size in beef cattle1. J. Anim. Sci. 2012, 90, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Daniel, Z.C.T.R.; Brameld, J.M.; Craigon, J.; Scollan, N.D.; Buttery, P.J. Effect of maternal dietary restriction during pregnancy on lamb carcass characteristics and muscle fiber composition1. J. Anim. Sci. 2007, 85, 1565–1576. [Google Scholar] [CrossRef] [PubMed]
- Dedkov, E.I.; Zheng, W.; Christensen, L.P.; Weiss, R.M.; Mahlberg-Gaudin, F.; Tomanek, R.J. Preservation of coronary reserve by ivabradine-induced reduction in heart rate in infarcted rats is associated with decrease in perivascular collagen. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, 590–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Phillips, E.; Riggins, T.; Sangha, G.; Chakraborty, S.; Lee, J.; Lycke, R.; Hernandez, C.; Soepriatna, A.; Thorne, B.; et al. Imaging of Small Animal Peripheral Artery Disease Models: Recent Advancements and Translational Potential. Int. J. Mol. Sci. 2015, 16, 11131–11177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bubb, K.J.; Cock, M.L.; Black, M.J.; Dodic, M.; Boon, W.M.; Parkington, H.C.; Harding, R.; Tare, M. Intrauterine growth restriction delays cardiomyocyte maturation and alters coronary artery function in the fetal sheep. J. Physiol. 2007, 578, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.J.; Campbell, M.E.; McMillen, I.C.; Davidge, S.T. Differential effects of maternal hypoxia or nutrient restriction on carotid and femoral vascular function in neonatal rats. Am. J. Physiol. Integr. Comp. Physiol. 2005, 288, R360–R367. [Google Scholar] [CrossRef] [Green Version]
- Dong, F.; Ford, S.P.; Nijland, M.J.; Nathanielsz, P.W.; Ren, J. Influence of maternal undernutrition and overfeeding on cardiac ciliary neurotrophic factor receptor and ventricular size in fetal sheep. J. Nutr. Biochem. 2008, 19, 409–414. [Google Scholar] [CrossRef] [Green Version]
- Colavitti, R.; Pani, G.; Bedogni, B.; Anzevino, R.; Borrello, S.; Waltenberger, J.; Galeotti, T. Reactive Oxygen Species as Downstream Mediators of Angiogenic Signaling by Vascular Endothelial Growth Factor Receptor-2/KDR. J. Biol. Chem. 2002, 277, 3101–3108. [Google Scholar] [CrossRef] [Green Version]
- Gacche, R.N.; Meshram, R.J. Targeting tumor micro-environment for design and development of novel anti-angiogenic agents arresting tumor growth. Prog. Biophys. Mol. Biol. 2013, 113, 333–354. [Google Scholar] [CrossRef]
- Wang, R.; Liu, H.; Xu, L.; Zhang, H.; Zhou, R. Melatonin downregulates nuclear receptor RZR/RORγ expression causing growth-inhibitory and anti-angiogenesis activity in human gastric cancer cells in vitro and in vivo. Oncol. Lett. 2016, 12, 897–903. [Google Scholar] [CrossRef] [Green Version]
- Reiter, R.J.; Tan, D.X. Melatonin: A novel protective agent against oxidative injury of the ischemic/reperfused heart. Cardiovasc. Res. 2003, 58, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Yang, H.-L.; Gu, C.-J.; Liu, Y.-K.; Shao, J.; Zhu, R.; He, Y.-Y.; Zhu, X.-Y.; Li, M.-Q. Melatonin restricts the viability and angiogenesis of vascular endothelial cells by suppressing HIF-1α/ROS/VEGF. Int. J. Mol. Med. 2019, 43, 945–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoppeler, H.; Kayar, S. Capillarity and Oxidative Capacity of Muscles. Physiology 1988, 3, 113–116. [Google Scholar] [CrossRef]
- Lim, K.; Zimanyi, M.A.; Black, M.J. Effect of maternal protein restriction in rats on cardiac fibrosis and capillarization in adulthood. Pediatr. Res. 2006, 60, 83–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Yaniz-Galende, E.; Kagan, H.J.; Liang, L.; Hekmaty, S.; Giannarelli, C.; Hajjar, R.J. Abnormalities of capillary microarchitecture in a rat model of coronary ischemic congestive heart failure. Am. J. Physiol. Heart Circ. Physiol. 2015, 308, H830–H840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roseboom, T.J.; van der Meulen, J.H.P.; Ravelli, A.C.J.; Osmond, C.; Barker, D.J.P.; Bleker, O.P. Perceived health of adults after prenatal exposure to the Dutch famine. Paediatr. Perinat. Epidemiol. 2003, 17, 391–397. [Google Scholar] [CrossRef]
- Reynolds, L.P.; Borowicz, P.P.; Caton, J.S.; Vonnahme, K.A.; Luther, J.S.; Hammer, C.J.; Maddock Carlin, K.R.; Grazul-Bilska, A.T.; Redmer, D.A. Developmental programming: The concept, large animal models, and the key role of uteroplacental vascular development1,2. J. Anim. Sci. 2010, 88, E61–E72. [Google Scholar] [CrossRef]
- Reiter, R.J.; Calvo, J.R.; Karbownik, M.; Qi, W.; Tan, D.X. Melatonin and Its Relation to the Immune System and Inflammation. Ann. N. Y. Acad. Sci. 2006, 917, 376–386. [Google Scholar] [CrossRef]
- Corstius, H.B.; Zimanyi, M.A.; Maka, N.; Herath, T.; Thomas, W.; Van Der Laarse, A.; Wreford, N.G.; Black, M.J. Effect of Intrauterine Growth Restriction on the Number of Cardiomyocytes in Rat Hearts. Pediatr. Res. 2005, 57, 796–800. [Google Scholar] [CrossRef] [Green Version]
- McMullen, J.R.; Jennings, G.L. Differences between pathological and physiological cardiac hypertrophy: Novel therapeutic strategies to treat heart failure. Clin. Exp. Pharmacol. Physiol. 2007, 34, 255–262. [Google Scholar] [CrossRef]
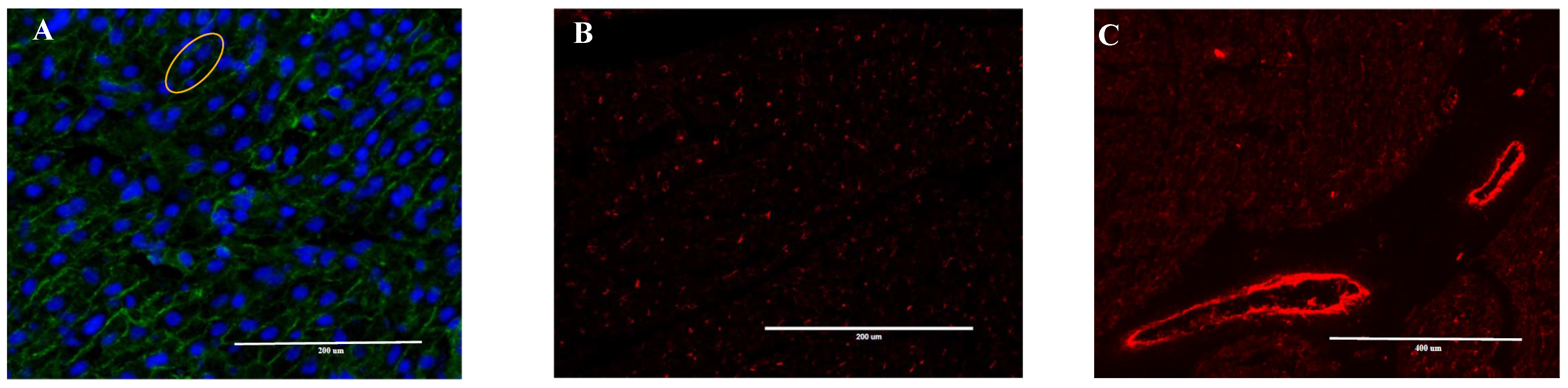


{kind=link}
{kind=link}
{kind=link}
{kind=link}
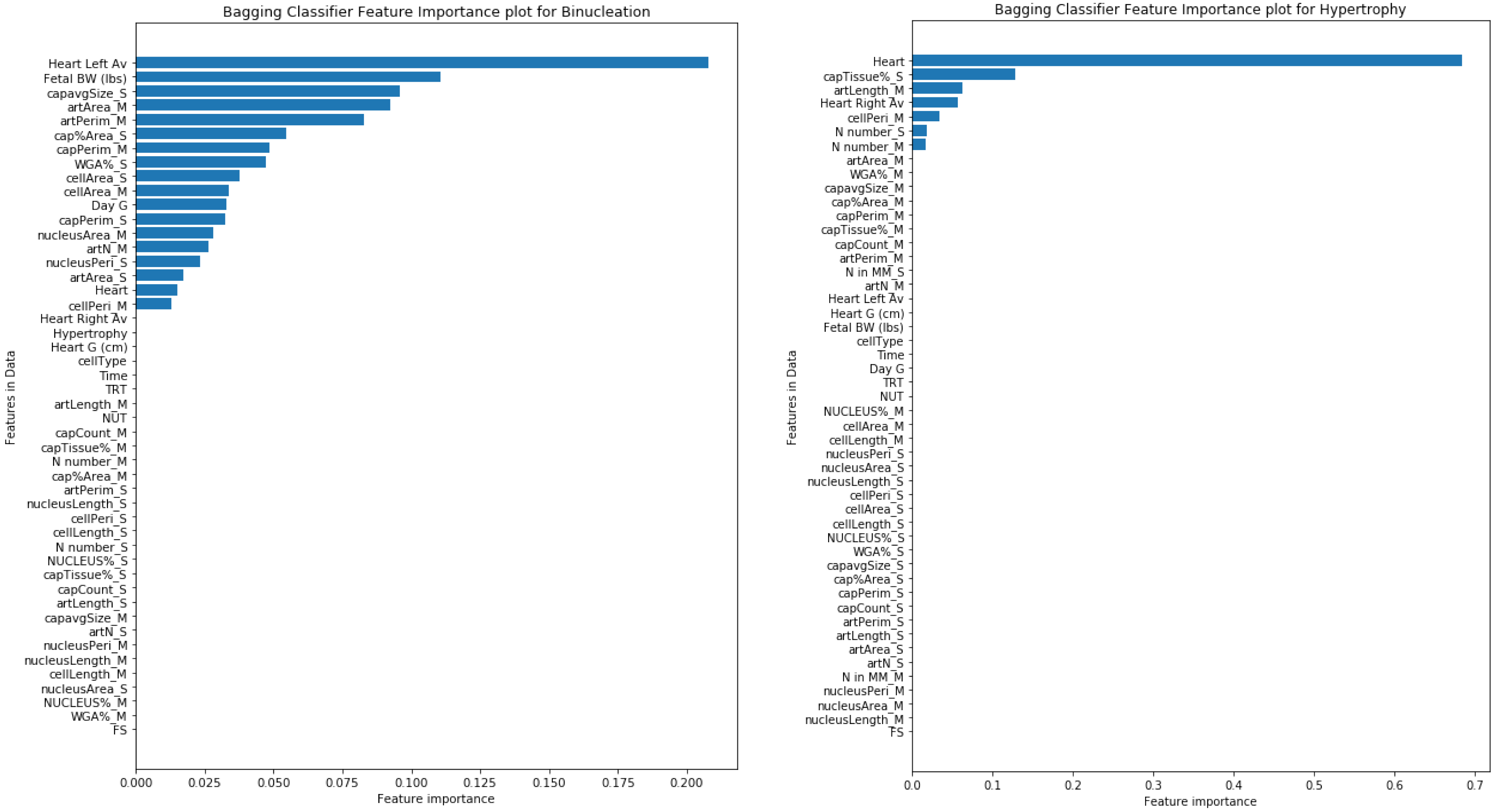{kind=link}
| Dependent Variable | ADQ-CON | ADQ-MEL | RES-CON | RES-MEL | SE | p-Value | ||
|---|---|---|---|---|---|---|---|---|
| Nut | trt | Nut*trt | ||||||
| Maternal BW (kg) | 575.7 | 580.2 | 531.8 | 536.2 | 18.8 | 0.01 | 0.87 | 0.99 |
| Fetal BW (kg) | 26.7 | 26.9 | 23.3 | 24.6 | 1.0 | 0.01 | 0.23 | 0.62 |
| Thoracic girth (cm) | 64.2 | 64.2 | 60.9 | 62.7 | 0.9 | 0.01 | 0.31 | 0.35 |
| Heart weight (g) | 186.2 | 187.2 | 173.7 | 174.2 | 9.6 | 0.17 | 0.94 | 0.98 |
| Left ventricle thickness (mm) | 10.4 | 11.1 | 11.1 | 10.7 | 0.8 | 0.84 | 0.83 | 0.48 |
| Right ventricle thickness (mm) | 7.2 | 7.8 | 5.8 | 6.4 | 0.4 | 0.001 | 0.12 | 0.96 |
| Heart thickness/heart weight | 0.09 | 0.10 | 0.10 | 0.10 | 0.05 | 0.69 | 0.27 | 0.82 |
| Dependent Variable | ADQ-CON | ADQ-MEL | RES-CON | RES-MEL | SE | p-Value | ||
|---|---|---|---|---|---|---|---|---|
| Nut | Trt | Nut*trt | ||||||
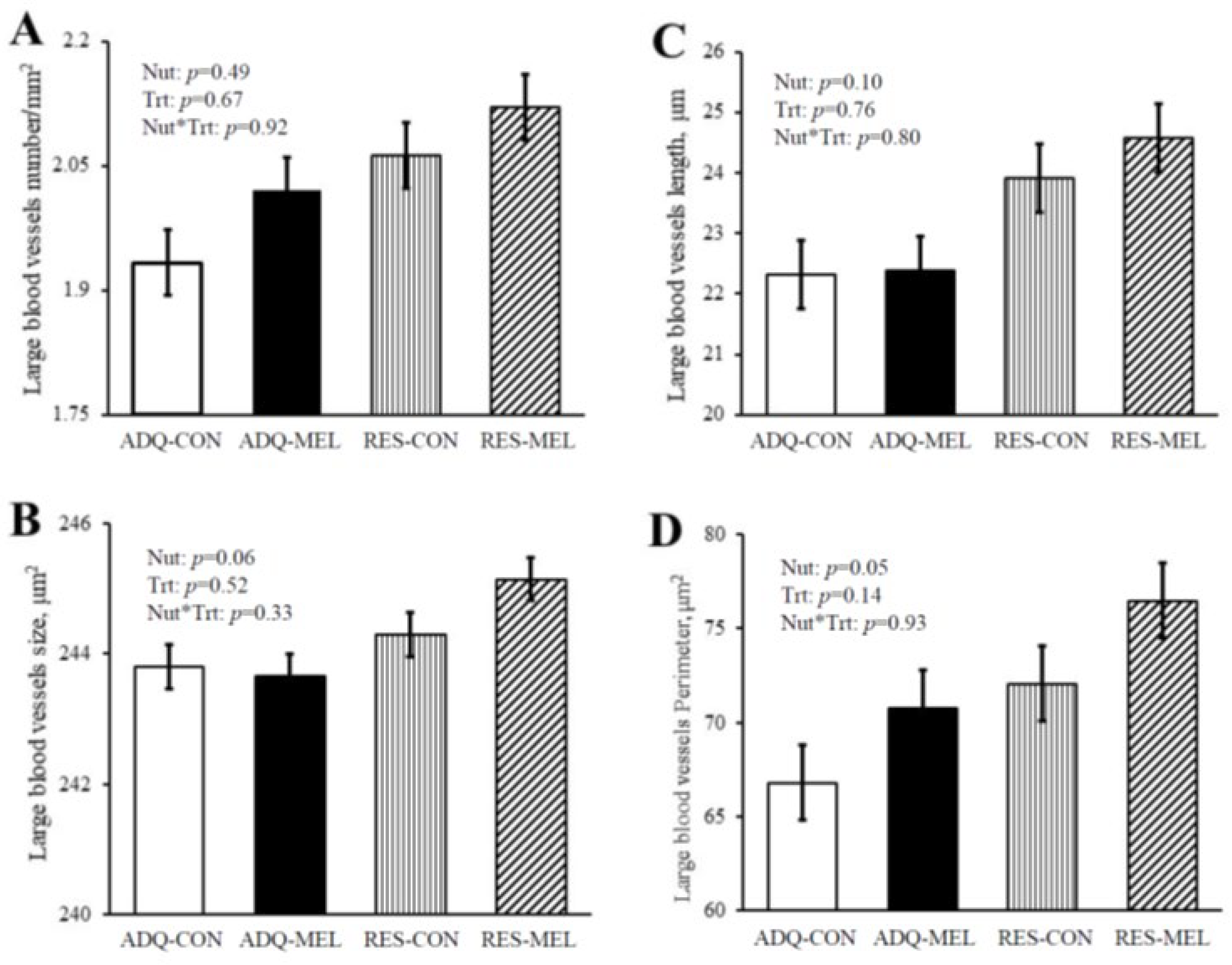
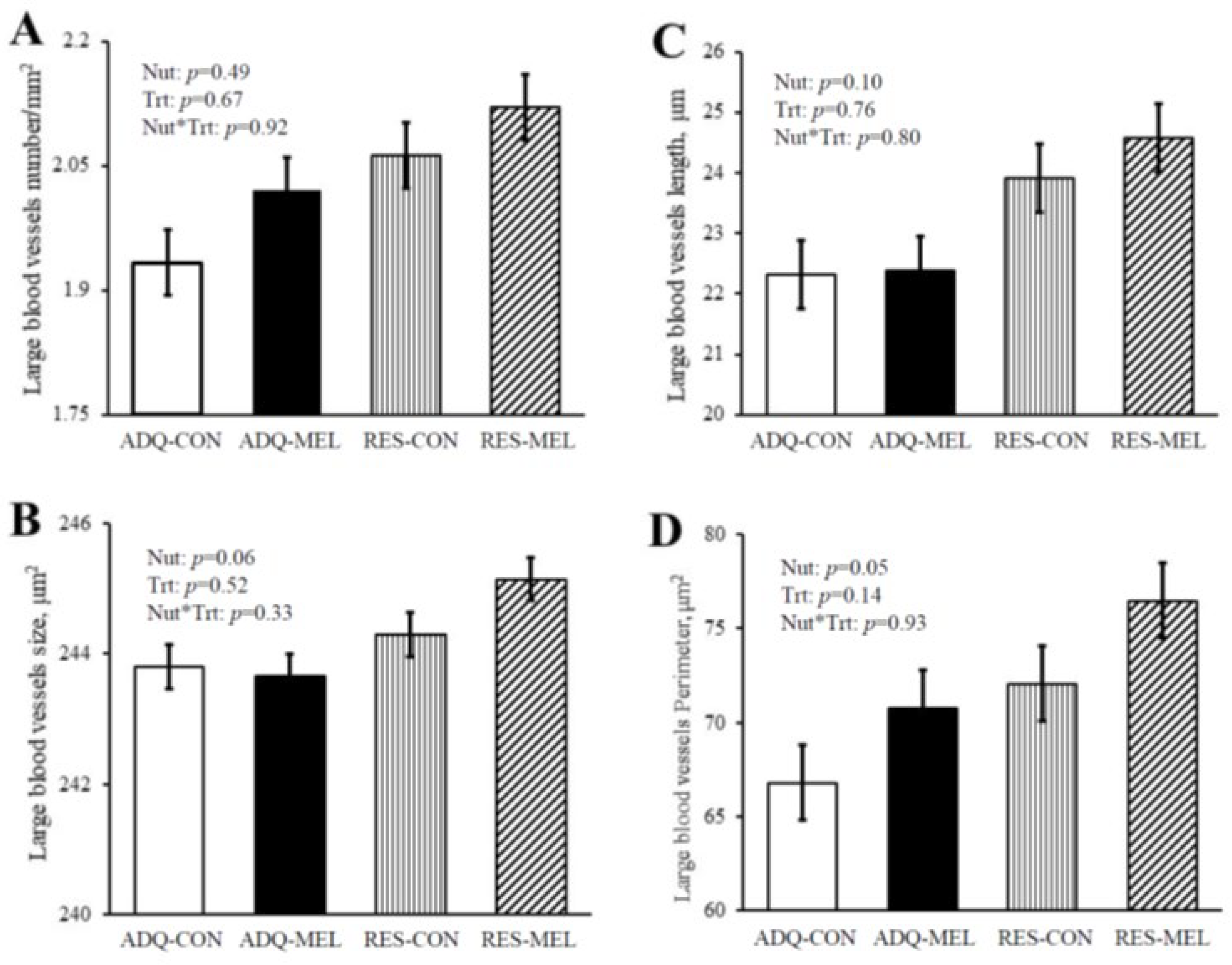
| Capillary number per mm2 | 1142 ab | 1106 b | 1139 ab | 1172 a | 15.47 | 0.03 | 0.93 | 0.01 |
| Percent capillary area, %/mm2 | 7.01 a | 6.08 c | 6.49 b | 6.48 bc | 0.15 | 0.70 | 0.002 | 0.0023 |
| Capillary size, μm2 | 20.77 a | 19.02 b | 18.80 b | 19.17 b | 0.36 | 0.01 | 0.05 | 0.002 |
| Total capillary perimeter (mm−1) | 90.07 a | 79.36 c | 85.04 b | 83.69 b | 1.82 | 0.84 | 0.0007 | 0.0078 |
| Dependent Variable | ADQ-CON | ADQ-MEL | RES-CON | RES-MEL | SE | p-Value | ||
|---|---|---|---|---|---|---|---|---|
| Nut | Trt | Nut*trt | ||||||
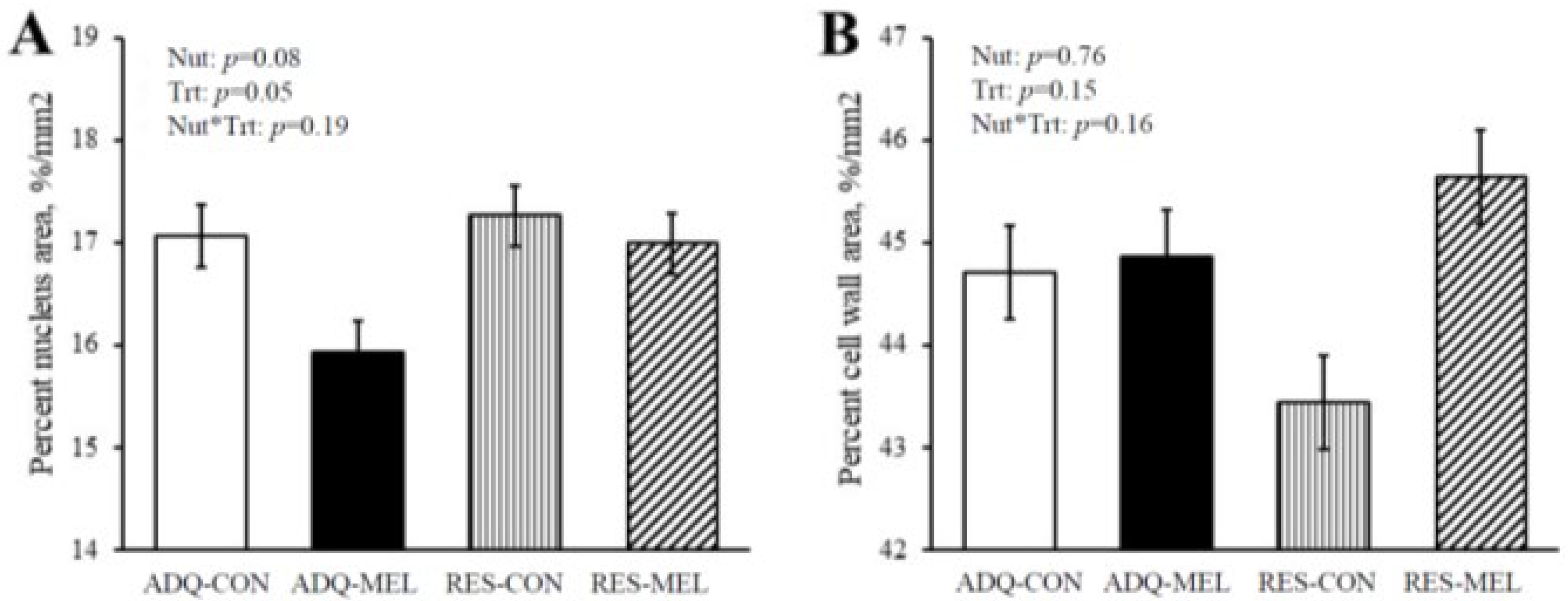
| Binucleated cells number per mm2 | 388 a | 284 b | 281 b | 322 ab | 27.28 | 0.21 | 0.25 | 0.010 |
| Cardiomyocyte binucleation % | 7.66 | 6.33 | 4.78 | 5.46 | 0.59 | 0.001 | 0.58 | 0.054 |
| Binucleated nucleus length (μm) | 5.76 | 5.76 | 4.86 | 4.85 | 0.20 | <0.0001 | 0.98 | 0.98 |
| Mononucleated nucleus length (μm) | 5.77 b | 6.01 a | 5.62 b | 5.28 c | 0.087 | <0.0001 | 0.56 | 0.0005 |
| Binucleated cell length (μm) | 15.96 | 16.98 | 15.98 | 14.16 | 0.81 | 0.061 | 0.58 | 0.07 |
| Mononucleated cell length (μm) | 11.72 | 11.99 | 10.53 | 10.22 | 0.20 | <0.0001 | 0.93 | 0.11 |
| Binucleated nucleus size (μm2) | 18.91 | 20.33 | 15.59 | 14.89 | 0.83 | <0.0001 | 0.66 | 0.14 |
| Mononucleated nucleus size (μm2) | 19.75 | 19.40 | 16.46 | 14.67 | 0.45 | <0.0001 | 0.015 | 0.06 |
| Binucleated nucleus perimeter (μm) | 16.54 | 17.05 | 15.27 | 14.75 | 0.37 | <0.0001 | 0.99 | 0.11 |
| Mononucleated nucleus perimeter (μm) | 17.14 | 16.89 | 15.80 | 14.73 | 0.20 | <0.0001 | 0.0010 | 0.019 |
| Binucleated cell size (μm2) | 91.44 b | 121.16 a | 80.79 b | 77.48 b | 6.73 | <0.0001 | 0.05 | 0.005 |
| Mononucleated cell size (μm2) | 60.33 b | 66.52 a | 51.12 cd | 47.53 d | 1.71 | <0.0001 | 0.43 | 0.0008 |
| Binucleated cell perimeter (μm) | 41.62 b | 46.28 a | 38.17 bc | 36.62 c | 1.61 | <0.0001 | 0.34 | 0.02 |
| Mononucleated cell perimeter (μm) | 32.64 a | 33.76 a | 29.84 b | 28.87 b | 0.50 | <0.0001 | 0.88 | 0.015 |
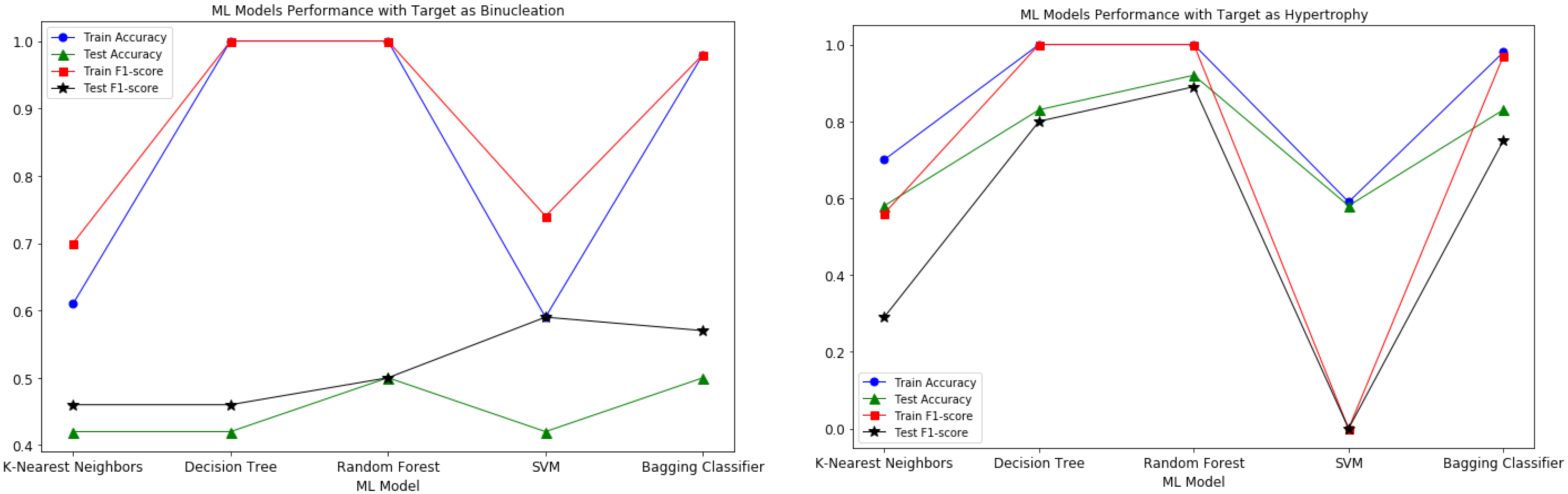
| Target Variable | Accuracy | F1-Score | ROC AUC | |||
|---|---|---|---|---|---|---|
| Training | Test | Training | Test | Training | Test | |
| Hypertrophy | 0.935 | 0.833 | 0.919 | 0.800 | 0.929 | 0.829 |
| N in mm * (Average) | 0.978 | 0.750 | 0.981 | 0.571 | 0.981 | 0.700 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazinani, M.; Contreras-Correa, Z.E.; Behzadan, V.; Gopal, S.; Lemley, C.O. Effects of Maternal Nutrient Restriction and Melatonin Supplementation on Cardiomyocyte Cell Development Parameters Using Machine Learning Techniques. Animals 2022, 12, 1818. https://doi.org/10.3390/ani12141818
Mazinani M, Contreras-Correa ZE, Behzadan V, Gopal S, Lemley CO. Effects of Maternal Nutrient Restriction and Melatonin Supplementation on Cardiomyocyte Cell Development Parameters Using Machine Learning Techniques. Animals. 2022; 12(14):1818. https://doi.org/10.3390/ani12141818
Chicago/Turabian StyleMazinani, Mitra, Zully E. Contreras-Correa, Vahid Behzadan, Shreya Gopal, and Caleb O. Lemley. 2022. "Effects of Maternal Nutrient Restriction and Melatonin Supplementation on Cardiomyocyte Cell Development Parameters Using Machine Learning Techniques" Animals 12, no. 14: 1818. https://doi.org/10.3390/ani12141818
APA StyleMazinani, M., Contreras-Correa, Z. E., Behzadan, V., Gopal, S., & Lemley, C. O. (2022). Effects of Maternal Nutrient Restriction and Melatonin Supplementation on Cardiomyocyte Cell Development Parameters Using Machine Learning Techniques. Animals, 12(14), 1818. https://doi.org/10.3390/ani12141818