
The Growth Performance, Nutrient Digestibility, Gut Bacteria and Bone Strength of Broilers Offered Alternative, Sustainable Diets Varying in Nutrient Specification and Phytase Dose

 , , ,
, , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Declaration
2.2. Diet Treatments
2.3. Performance Trial
2.4. Digestibility Trial
2.4.1. Chemical Analysis
2.4.2. 16S rRNA Profiling
2.4.3. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Bone Strength
3.3. Digestibility Results
3.4. Jejunum Microbiota
3.5. Nitrogen Balance
3.6. Phosphorus Excretion
3.7. Phytate, Lower Phytate Esters and Inositol
4. Discussion
4.1. Bird Growth Performance
4.2. Phytase, Phytate Degradation and Nutrient Digestibility
4.3. Phytase and N and P Excretion
4.4. Bone Strength and Body Composition
4.5. Jejunum Microbiota Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ravindran, V.; Cowieson, A.J.; Selle, P.H. Influence of dietary electrolyte balance and microbial phytase on growth performance, nutrient utilization, and excreta quality of broiler chickens. Poult. Sci. 2008, 87, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Walk, C.L.; Rao, S.R. Dietary phytate has a greater anti-nutrient effect on feed conversion ratio compared to body weight gain and greater doses of phytase are required to alleviate this effect as evidenced by prediction equations on growth performance, bone ash and phytate degradation in broilers. Poult. Sci. 2020, 99, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, F.; Fakhur-un-Nisa, T.; Liu, J.; Luo, X.; Abbasi, I.H.R. Low digestibility of phytate phosphorus, their impacts on the environment, and phytase opportunity in the poultry industry. Environ. Sci. Pollut. Res. 2019, 26, 9469–9479. [Google Scholar] [CrossRef]
- Selle, P.H.; Ravindran, V.; Caldwell, A.; Bryden, W.L. Phytate and phytase: Consequences for protein utilisation. Nutr. Res. Rev. 2000, 13, 255–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, F.R.; Hruby, M.; Pierson, E.E.M.; Remus, J.C.; Sakomura, N.K. Effect of phytase supplementation in diets on nutrient digestibility and performance in broiler chicks. J. Appl. Poult. Res. 2008, 17, 191–201. [Google Scholar] [CrossRef]
- Attia, Y.A.; Bovera, F.; Al-Harthi, M.A.; El-Din, A.E.R.E.; Said Selim, W. Supplementation of microbial and fungal phytases to low protein and energy diets: Effects on productive performance, nutrient digestibility, and blood profiles of broilers. Agriculture 2021, 11, 414. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; Dusel, G. Increasing the dosing of a Buttiauxella phytase improves phytate degradation, mineral, energy, and amino acid digestibility in weaned pigs fed a complex diet based on wheat, corn, soybean meal, barley, and rapeseed meal. J. Anim. Sci. 2019, 97, 2524–2533. [Google Scholar] [CrossRef] [Green Version]
- Dersjant-Li, Y.; Davin, R.; Christensen, T.; Kwakernaak, C. Effect of two phytases at two doses on performance and phytate degradation in broilers during 1–21 days of age. PLoS ONE 2021, 16, e0247420. [Google Scholar] [CrossRef]
- Walters, H.G.; Coelho, M.; Coufal, C.D.; Lee, J.T. Effects of increasing phytase inclusion levels on broiler performance, nutrient digestibility, and bone mineralization in low-phosphorus diets. J. Appl. Poult. Res. 2019, 28, 1210–1225. [Google Scholar] [CrossRef]
- Powell, S.; Bidner, T.D.; Southern, L.L. Phytase supplementation improved growth performance and bone characteristics in broilers fed varying levels of dietary calcium. Poult. Sci. 2011, 90, 604–608. [Google Scholar] [CrossRef]
- Walk, C.L.; Bedford, M.R.; Santos, T.S.; Paiva, D.; Bradley, J.R.; Wladecki, H.; Honaker, C.; McElroy, A.P. Extra-phosphoric effects of superdoses of a novel microbial phytase. Poult. Sci. 2013, 92, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Leyva-Jimenez, H.; Alsadwi, A.M.; Gardner, K.; Voltura, E.; Bailey, C.A. Evaluation of high dietary phytase supplementation on performance, bone mineralization, and apparent ileal digestible energy of growing broilers. Poult. Sci. 2019, 98, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Al-Qahtani, M.; Ahiwe, E.U.; Abdallh, M.E.; Chang’a, E.P.; Gausi, H.; Bedford, M.R.; Iji, P.A. Endogenous enzyme activities and tibia bone development of broiler chickens fed wheat-based diets supplemented with xylanase, β-glucanase and phytase. Anim. Biosci. 2021, 34, 1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukumar, D.; Jalaludeen, A.; Cyriac, S. Effect of mitigating environmental pollution by incorporation of phytase enzyme in feed of (Athulya) layer chicken. Wayamba J. Anim. Sci. 2013, 5, 569–573. [Google Scholar]
- Musapuor, A.; Afsharmanesh, M.; Shahrbabak, H.M. Use of microbial phytase for decrease of pollutant due to environmental poultry excreta phosphorus. Int. J. Agric. Biol. 2006, 8, 35–37. [Google Scholar]
- Bougouin, A.; Appuhamy, J.A.D.R.N.; Kebreab, E.; Dijkstra, J.; Kwakkel, R.P.; France, J. Effects of phytase supplementation on phosphorus retention in broilers and layers: A meta-analysis. Poult. Sci. 2014, 93, 1981–1992. [Google Scholar] [CrossRef]
- Lalpanmawia, H.; Elangovan, A.V.; Sridhar, M.; Shet, D.; Ajith, S.; Pal, D.T. Efficacy of phytase on growth performance, nutrient utilization and bone mineralization in broiler chicken. Anim. Feed Sci. Technol. 2014, 192, 81–89. [Google Scholar] [CrossRef]
- Puppala, K.R.; Buddhiwant, P.G.; Agawane, S.B.; Kadam, A.S.; Mote, C.S.; Lonkar, V.D.; Khire, J.M.; Dharne, M.S. Performance of Aspergillus niger (NCIM 563) phytase based feed supplement for broiler growth and phosphorus excretion. Biocatal. Agric. Biotechnol. 2021, 31, 101887. [Google Scholar] [CrossRef]
- Humer, E.; Schwarz, C.; Schedle, K. Phytate in pig and poultry nutrition. J. Anim. Physiol. Anim. Nutr. 2015, 99, 605–625. [Google Scholar] [CrossRef]
- Scholey, D.V.; Morgan, N.K.; Riemensperger, A.; Hardy, R.; Burton, E.J. Effect of supplementation of phytase to diets low in inorganic phosphorus on growth performance and mineralization of broilers. Poult. Sci. 2018, 97, 2435–2440. [Google Scholar] [CrossRef]
- Babatunde, O.O.; Bello, A.; Dersjant-Li, Y.; Adeola, O. Evaluation of the responses of broiler chickens to varying concentrations of phytate phosphorus and phytase. Ⅰ. Starter phase (day 1–11 post hatching). Poult. Sci. 2021, 100, 101396. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.I.M.; Horn, D.; Ronconi, E.J.; Buzim, R.; Lima, F.K.; Pazdiora, D.A. Effects of Phytase Superdosing on Digestibility and Bone Integrity of Broilers. J. Appl. Poult. Res. 2019, 28, 390–398. [Google Scholar] [CrossRef]
- Walk, C.L.; Santos, T.T.; Bedford, M.R. Influence of superdoses of a novel microbial phytase on growth performance, tibia ash, and gizzard phytate and inositol in young broilers. Poult. Sci. 2014, 93, 1172–1177. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.A.; Nagalakshmi, D.; Raju, M.V.; Rao, S.V.R.; Bedford, M.R. Effect of phytase superdosing, myo-inositol and available phosphorus concentrations on performance and bone mineralisation in broilers. Anim. Nutr. 2017, 3, 247–251. [Google Scholar] [CrossRef]
- Infanzón, B.; Herrmann, K.R.; Hofmann, I.; Willbold, S.; Ruff, A.J.; Schwaneberg, U. Phytase blends for enhanced phosphorous mobilization of deoiled seeds. Enzyme Microb. Technol. 2022, 153, 109953. [Google Scholar] [CrossRef]
- Zanu, H.K.; Keerqin, C.; Kheravii, S.K.; Morgan, N.; Wu, S.B.; Bedford, M.R.; Swick, R.A. Influence of meat and bone meal, phytase, and antibiotics on broiler chickens challenged with subclinical necrotic enteritis: 2. intestinal permeability, organ weights, hematology, intestinal morphology, and jejunal gene expression. Poult. Sci. 2020, 99, 2581–2594. [Google Scholar] [CrossRef]
- Moita, V.H.C.; Duarte, M.E.; Kim, S.W. Supplemental Effects of Phytase on Modulation of Mucosa-Associated Microbiota in the Jejunum and the Impacts on Nutrient Digestibility, Intestinal Morphology, and Bone Parameters in Broiler Chickens. Animals 2021, 11, 3351. [Google Scholar] [CrossRef]
- Ptak, A.; Bedford, M.R.; Świątkiewicz, S.; Żyła, K.; Józefiak, D. Phytase modulates ileal microbiota and enhances growth performance of the broiler chickens. PLoS ONE 2015, 10, e0119770. [Google Scholar] [CrossRef] [Green Version]
- Ball, M.E.; Beattie, V.; Hayes, H.; Lawlor, P.; Magowan, E. The use of enzymes in pigs and broiler diets containing low soya and varying levels of energy and amino acids. In Proceedings of the 67th EAAP Annual Meeting, Belfast, UK, 29 August–2 September 2016. [Google Scholar]
- Hollands, C. The animals (scientific procedures) act 1986. Lancet Lond. 1986, 2, 32–33. [Google Scholar] [CrossRef]
- Crenshaw, T.D.; Peo, E.R., Jr.; Lewis, A.J.; Moser, B.D. Bone strength as a trait for assessing mineralization in swine: A critical review of techniques involved. J. Anim. Sci. 1981, 53, 827–835. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1995. [Google Scholar]
- Van Soest, P.J. The use of detergent in the analysis of fibrous feeds. II. A rapid method for the determination of fiber and lignin. J. Assoc. Off. Agric. Chem. 1963, 46, 829–835. [Google Scholar] [CrossRef]
- Leone, J.L. Collaborative study of the quantitative determination of titanium dioxide in cheese. AOAC 1973, 56, 535–537. [Google Scholar] [CrossRef]
- Peddie, J.; Dewar, W.A.; Gilbert, A.B.; Waddington, D. The use of titanium dioxide for determining apparent ileal digestibility in mature domestic fowls (Gallus domesticus). J. Agric. Sci. 1982, 99, 233–236. [Google Scholar] [CrossRef]
- European Commission Commission regulation (EC). No 152/2009 of 27 January 2009 laying down the methods of sampling and analysis for the official control of feed. Off. J. Eur. Union. 2009, 54, 1–130. [Google Scholar]
- Lu, H.; Kühn, I.; Bedford, M.R.; Whitfield, H.; Brearley, C.; Adeola, O.; Ajuwon, K.M. Effect of phytase on intestinal phytate breakdown, plasma inositol concentrations, and glucose transporter type 4 abundance in muscle membranes of weanling pigs. J. Anim. Sci. 2019, 97, 3907–3919. [Google Scholar] [CrossRef]
- Whitfield, H.; White, G.; Sprigg, C.; Riley, A.M.; Potter, B.V.; Hemmings, A.M.; Brearley, C.A. An ATP-responsive metabolic cassette comprised of inositol tris/tetrakisphosphate kinase 1 (ITPK1) and inositol pentakisphosphate 2-kinase (IPK1) buffers diphosphosphoinositol phosphate levels. Biochem. J. 2020, 477, 2621–2638. [Google Scholar] [CrossRef]
- Madsen, C.K.; Brearley, C.A.; Brinch-Pedersen, H. Lab-scale preparation and QC of phytase assay substrate from rice bran. Anal. Biochem. 2019, 578, 7–12. [Google Scholar] [CrossRef]
- Schiavone, A.; Guo, K.; Tassone, S.; Gasco, L.; Hernandez, E.; Denti, R.; Zoccarato, I. Effects of a natural extract of chestnut wood on digestibility, performance traits, and nitrogen balance of broiler chicks. Poult. Sci. 2008, 87, 521–527. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Hugerth, L.W.; Wefer, H.A.; Lundin, S.; Jakobsson, H.E.; Lindberg, M.; Rodin, S.; Engstrand, L.; Andersson, A.F. DegePrime, a program for degenerate primer design for broad-taxonomic-range PCR in microbial ecology studies. Appl. Environ. Microbiol. 2014, 80, 5116–5123. [Google Scholar] [CrossRef] [Green Version]
- Akter, M.; Graham, H.; Iji, P.A. Response of broiler chickens to diets containing different levels of sodium with or without microbial phytase supplementation. J. Anim. Sci. Technol. 2019, 61, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faridi, A.; Gitoee, A.; France, J. A meta-analysis of the effects of nonphytate phosphorus on broiler performance and tibia ash concentration. Poult. Sci. 2015, 94, 2753–2762. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; He, Z.Z.; Ao, X.; Sun, W.L.; Xiao, X.; Zeng, F.K.; Wang, Y.C.; He, J. Effects of residual superdoses of phytase on growth performance, tibia mineralization, and relative organ weight in ducks fed phosphorus-deficient diets. Poult. Sci. 2019, 98, 3926–3936. [Google Scholar] [CrossRef] [PubMed]
- Lima, G.S.; Lima, M.R.; Gomes, G.A.; Cavalcante, D.T.; Guerra, R.R.; da Silva, J.H.V.; Cardoso, A.S.; Kaneko, I.N.; Costa, F.G.P. Superdosing of bacterial phytase (EC 3.1. 3.26) in broiler diets with reduced levels of digestible amino acids. Livest. Sci. 2021, 253, 104714. [Google Scholar] [CrossRef]
- Pieniazek, J.; Smith, K.A.; Williams, M.P.; Manangi, M.K.; Vazquez-Anon, M.; Solbak, A.; Miller, M.; Lee, J.T. Evaluation of increasing levels of a microbial phytase in phosphorus deficient broiler diets via live broiler performance, tibia bone ash, apparent metabolizable energy, and amino acid digestibility. Poult. Sci. 2017, 96, 370–382. [Google Scholar] [CrossRef]
- Manobhavan, M.; Sridhar, M.; Ajith, S.; Shet, D.; Pal, D.T.; Gowda, N.K.S.; Elangovan, A.V. Efficacy of fungal phytase on growth performance and bone mineralization in broiler chicken. Anim. Nutr. Feed Technol. 2015, 15, 129–136. [Google Scholar] [CrossRef]
- Wyss, M.; Brugger, R.; Kronenberger, A.; Rémy, R.; Fimbel, R.; Oesterhelt, G.; Lehmann, M.; Van Loon, A.P. Biochemical characterization of fungal phytases (myo-inositol hexakisphosphate phosphohydrolases): Catalytic properties. Appl. Environ. Microbiol. 1999, 65, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Cowieson, A.J.; Wilcock, P.; Bedford, M.R. Super-dosing effects of phytase in poultry and other monogastrics. World’s Poult. Sci. J. 2011, 67, 225–236. [Google Scholar] [CrossRef]
- Gautier, A.E.; Walk, C.L.; Dilger, R.N. Effects of a high level of phytase on broiler performance, bone ash, phosphorus utilization, and phytate dephosphorylation to inositol. Poult. Sci. 2018, 97, 211–218. [Google Scholar] [CrossRef]
- Beeson, L.A.; Walk, C.L.; Bedford, M.R.; Olukosi, O.A. Hydrolysis of phytate to its lower esters can influence the growth performance and nutrient utilization of broilers with regular or super doses of phytase. Poult. Sci. 2017, 96, 2243–2253. [Google Scholar] [CrossRef]
- Walk, C.L.; Bedford, M.R.; Olukosi, O.A. Effect of phytase on growth performance, phytate degradation and gene expression of myo-inositol transporters in the small intestine, liver and kidney of 21 day old broilers. Poult. Sci. 2018, 97, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Bedford, M.; Rousseau, X. Recent findings regarding calcium and phytase in poultry nutrition. Anim. Prod. Sci. 2017, 57, 2311–2316. [Google Scholar] [CrossRef]
- Pontoppidan, K.; Glitsoe, V.; Guggenbuhl, P.; Quintana, A.P.; Nunes, C.S.; Pettersson, D.; Sandberg, A.S. In vitro and in vivo degradation of myo-inositol hexakisphosphate by a phytase from Citrobacter braakii. Arch. Anim. Nutr. 2012, 66, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Carré, B. Causes for variation in digestibility of starch among feedstuffs. World’s. Poult. Sci. J. 2004, 60, 76–89. [Google Scholar] [CrossRef]
- Zeller, E.; Schollenberger, M.; Witzig, M.; Shastak, Y.; Kühn, I.; Hoelzle, L.E.; Rodehutscord, M. Interactions between supplemented mineral phosphorus and phytase on phytate hydrolysis and inositol phosphates in the small intestine of broilers. Poult. Sci. 2015, 94, 1018–1029. [Google Scholar] [CrossRef] [PubMed]
- Cowieson, A.J.; Aureli, R.; Guggenbuhl, P.; Fru-Nji, F. Possible involvement of myo-inositol in the physiological response of broilers to high doses of microbial phytase. Anim. Poult. Sci. 2014, 55, 710–719. [Google Scholar] [CrossRef]
- Vieira, S.L.; Anschau, D.L.; Serafini, N.C.; Kindlein, L.; Cowieson, A.J.; Sorbara, J.O.B. Phosphorus equivalency of a Citrobracter braakii phytase in broilers. J. Appl. Poult. Res. 2015, 24, 335–342. [Google Scholar] [CrossRef]
- Manobhavan, M.; Elangovan, A.V.; Sridhar, M.; Shet, D.; Ajith, S.; Pal, D.T.; Gowda, N.K.S. Effect of super dosing of phytase on growth performance, ileal digestibility and bone characteristics in broilers fed corn–soya-based diets. J. Anim. Physiol. Anim. Nutr. 2016, 100, 93–100. [Google Scholar] [CrossRef]
- Woyengo, T.A.; Wilson, J. Enhancing nutrient utilization in maize for broiler chickens by superdosing phytase. Anim. Feed Sci. Technol. 2019, 252, 34–40. [Google Scholar] [CrossRef]
- Poulsen, H.D.; Carlson, D.; Nørgaard, J.V.; Blaabjerg, K. Phosphorus digestibility is highly influenced by phytase but slightly by calcium in growing pigs. Livest. Sci. 2010, 134, 100–102. [Google Scholar] [CrossRef]
- Srikanthithasan, K.; Macelline, S.P.; Wickramasuriya, S.S.; Tharangani, H.; Jayasena, D.D.; Heo, J.M. Effects of adding phytase from aspergillus niger to a low phosphorus diet on growth performance, tibia characteristics, phosphorus excretion, and meat quality of broilers 35 days after hatching. Poult. Sci. J. 2019, 57, 28–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, T.C.; Dove, R.; Bedford, M.R.; Azain, M.J. Effect of phytase on phosphorous balance in 20-kg barrows fed low or adequate phosphorous diets. Anim. Nutr. 2020, 6, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Bento, M.H.L.; Pedersen, C.; Plumstead, P.W.; Salmon, L.; Nyachoti, C.M.; Bikker, P. Dose response of a new phytase on dry matter, calcium, and phosphorus digestibility in weaned piglets. J. Anim. Sci. 2012, 90, 245–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adedokun, S.A.; Owusu-Asiedu, A.; Ragland, D.; Plumstead, P.; Adeola, O. The efficacy of a new 6-phytase obtained from Buttiauxella spp. expressed in Trichoderma reesei on digestibility of amino acids, energy, and nutrients in pigs fed a diet based on corn, soybean meal, wheat middlings, and corn distillers’ dried grains with solubles. Sci. J. Anim. Sci. 2015, 93, 168–175. [Google Scholar] [CrossRef]
- Zeng, Z.; Li, Q.; Tian, Q.; Zhao, P.; Xu, X.; Yu, S.; Piao, X. Super high dosing with a novel Buttiauxella phytase continuously improves growth performance, nutrient digestibility, and mineral status of weaned pigs. Biol. Trace Elem. Res. 2015, 168, 103–109. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; Wealleans, A.L.; Barnard, L.P.; Lane, S. Effect of increasing Buttiauxella phytase dose on nutrient digestibility and performance in weaned piglets fed corn or wheat based diets. Anim. Feed Sci. Technol. 2017, 234, 101–109. [Google Scholar] [CrossRef]
- Afsharmanesh, M.; Scott, T.A.; Silversides, F.G. Effect of wheat type grinding, heat treatment and phytase supplementation on growth efficiency and nutrient utilization of wheat-based diets for broilers. Can. J. Anim. Sci. 2008, 88, 57–64. [Google Scholar] [CrossRef]
- Attia, Y.A.; Bovera, F.; Wang, J.; Al-Harthi, M.A.; Kim, W.K. Multiple amino acid supplementations to low-protein diets: Effect on performance, carcass yield, meat quality and nitrogen excretion of finishing broilers under hot climate conditions. Animals 2020, 10, 973. [Google Scholar] [CrossRef]
- Angel, R.; Saylor, W.W.; Mitchell, A.D.; Powers, W.; Applegate, T.J. Effect of dietary phosphorus, phytase, and 25-hydroxycholecalciferol on broiler chicken bone mineralization, litter phosphorus, and processing yields. Poult. Sci. 2006, 85, 1200–1211. [Google Scholar] [CrossRef]
- Chung, T.K.; Rutherfurd, S.M.; Thomas, D.V.; Moughan, P.J. Effect of two microbial phytases on mineral availability and retention and bone mineral density in low-phosphorus diets for broilers. Br. Poult. Sci. 2013, 54, 362–373. [Google Scholar] [CrossRef]
- Hakami, Z.; Al Sulaiman, A.R.; Alharthi, A.S.; Casserly, R.; Bouwhuis, M.A.; Abudabos, A.M. Growth performance, carcass and meat quality, bone strength, and immune response of broilers fed low-calcium diets supplemented with marine mineral complex and phytase. Poult. Sci. 2022, 101, 101849. [Google Scholar] [CrossRef] [PubMed]
- Diaz Carrasco, J.M.; Casanova, N.A.; Fernández Miyakawa, M.E. Microbiota, gut health and chicken productivity: What is the connection? Microorganisms 2019, 7, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourenco, J.M.; Rothrock Jr, M.J.; Fluharty, F.L.; Callaway, T.R. The successional changes in the gut microbiome of pasture-raised chickens fed soy-containing and soy-free diets. Front. Sustain. Food Syst. 2019, 3, 35. [Google Scholar] [CrossRef]
- Fusco, V.; Quero, G.M.; Cho, G.S.; Kabisch, J.; Meske, D.; Neve, H.; Bockelmann, W.; Franz, C.M. The genus Weissella: Taxonomy, ecology and biotechnological potential. Front. Microbiol. 2015, 6, 155. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Xiang, Y.; Zhou, W.; Chen, J.; Li, K.; Yang, H. Microbial community mapping in intestinal tract of broiler chicken. Poult. Sci. 2017, 96, 1387–1393. [Google Scholar] [CrossRef]
- Qaisrani, S.N.; Van Krimpen, M.M.; Verstegen, M.W.A.; Hendriks, W.H.; Kwakkel, R.P. Effects of three major protein sources on performance, gut morphology and fermentation characteristics in broilers. Br. Poult. Sci. 2020, 61, 43–50. [Google Scholar] [CrossRef]
- Nourmohammadi, R.; Afzali, N. Effect of citric acid and microbial phytase on small intestinal morphology in broiler chicken. Ital. J. Anim. Sci. 2013, 12, e7. [Google Scholar] [CrossRef]

{kind=link}
| Starter/Grower (0–28 d) | Finisher (29–42 d) | |||||||
|---|---|---|---|---|---|---|---|---|
| PC1 | PC2 | NC | NC+ | PC1 | PC2 | NC | NC+ | |
| Wheat, g/kg | 602 | 563 | 604 | 604 | 608 | 476 | 518 | 517 |
| Maize DDGS, g/kg | — | 35 | 35 | 35 | — | 100 | 100 | 100 |
| Soyabean meal, g/kg | 305 | 280 | 260 | 260 | 279 | 197 | 176 | 176 |
| Rapeseed meal extract, g/kg | — | 25 | 25 | 25 | — | 100 | 100 | 100 |
| Soya oil, g/kg | 24 | 27.9 | 11.4 | 11.5 | 52.7 | 68.8 | 52.2 | 52.3 |
| Salt, g/kg | 3.3 | 3 | 1.9 | 1.9 | 2.5 | 1.8 | 1.4 | 1.4 |
| Limestone, g/kg | 10.4 | 10.8 | 10.6 | 10.6 | 9.4 | 10.7 | 10.5 | 10.5 |
| Mono-Dicalcium Phosphate, g/kg | 11.1 | 10.1 | 7 | 7 | 8.2 | 5 | 1.9 | 1.9 |
| Sodium Bicarbonate, g/kg | 2 | 2 | 3 | 3 | 2 | 2 | 2 | 2 |
| Premix 1, g/kg | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 |
| L-lysine HCl, g/kg | 5.8 | 6.2 | 6.4 | 6.4 | 3.8 | 5 | 5.1 | 5.1 |
| Methionine, g/kg | 5 | 4.9 | 4.4 | 4.4 | 4 | 3.7 | 3.1 | 3.1 |
| Threonine, g/kg | 3.3 | 3.3 | 3 | 3 | 2.1 | 2.1 | 1.8 | 1.8 |
| Quantum Blue 5G (1500 FTU/kg) | — | — | — | 0.3 | — | — | — | 0.3 |
| Quantum Blue 5G (500 FTU/kg) | 0.1 | 0.1 | — | — | 0.1 | 0.1 | — | — |
| Formulated composition | ||||||||
| Crude protein, g/kg | 215 | 215 | 209 | 209 (215) | 200 | 200 | 194 | 194 (200) |
| Metabolizable energy, MJ/kg | 12.6 | 12.6 | 12.2 | 12.2 (12.6) | 13.4 | 13.4 | 13.1 | 13.1 (13.4) |
| Dry matter, g/kg | 857 | 858 | 856 | 856 | 859 | 866 | 863 | 863 |
| Calcium, g/kg | 9.6 | 9.6 | 7.2 | 7.2 (9.6) | 8.5 | 8.5 | 6.1 | 6.1 (8.5) |
| Phosphorus (P), g/kg | 7.2 | 7.3 | 5.1 | 5.1 (7.3) | 6.4 | 6.7 | 4.5 | 4.5 (6.7) |
| Phytate- P, g/kg | 2.4 | 2.5 | 2.5 | 2.5 | 2.3 | 2.6 | 2.6 | 2.6 |
| Non Phytate-P (nPP), g/kg | 4.8 | 4.8 | 2.6 | 2.6 (4.8) | 4.1 | 4.1 | 1.9 | 1.9 (4.1) |
| Ca: nPP, g/kg | 2 | 2 | 2.8 | 2.8 (2.0) | 2.1 | 2.1 | 3.2 | 3.2 (2.1) |
| Lysine, g/kg | 14.3 | 14.3 | 14 | 14 | 12 | 12.1 | 11.8 | 11.8 |
| Methionine, g/kg | 7.6 | 7.6 | 7 | 7 | 6.5 | 6.4 | 5.8 | 5.8 |
| Threonine, g/kg | 10.1 | 10.2 | 9.6 | 9.6 | 8.5 | 8.7 | 8.1 | 8.1 |
| Tryptophan, g/kg | 2.4 | 2.4 | 2.1 | 2.1 | 2.1 | 2.1 | 1.8 | 1.8 |
| Starter/Grower | Finisher | |||||||
|---|---|---|---|---|---|---|---|---|
| PC1 | PC2 | NC | NC+ | PC1 | PC2 | NC | NC+ | |
| Gross energy, MJ/kg | 18.89 | 19.07 | 18.89 | 18.56 | 19.40 | 20.16 | 19.95 | 19.93 |
| Apparent metabolizable energy, MJ/kg | 12.67 | 12.79 | 12.5 | 12.61 | 13.33 | 13.84 | 13.52 | 13.86 |
| Dry matter, g/kg | 862 | 860 | 863 | 854 | 877 | 884 | 878 | 882 |
| Ether extract *, g/kg | 48.1 | 55.7 | 47.0 | 38.9 | 75.1 | 105.0 | 90.2 | 92.0 |
| Ash *, g/kg | 69.0 | 68.1 | 63.6 | 63.5 | 60.9 | 62.4 | 60.9 | 57.2 |
| Neutral detergent fibre *, g/kg | 159 | 169.7 | 181.6 | 163.8 | 125.2 | 192.0 | 203.8 | 179.0 |
| Crude protein *, g/kg | 209 | 220 | 208 | 214 | 199 | 205 | 201 | 201 |
| Calcium, g/kg | 9.8 | 9.4 | 8.4 | 9.3 | 9.7 | 8.6 | 8.8 | 7.5 |
| Phosphorus, g/kg | 6.0 | 5.8 | 5.4 | 5.1 | 5.0 | 5.3 | 4.9 | 4.9 |
| Phytate-P, g/kg | 2.5 | 2.5 | 2.6 | 2.5 | 2.3 | 2.5 | 2.7 | 2.6 |
| Non phytate- phosphorus (nPP), g/kg | 3.5 | 3.3 | 2.8 | 2.6 | 2.7 | 2.8 | 2.2 | 2.3 |
| Lysine, g/kg | 15.1 | 15.1 | 13.7 | 14.8 | 12.8 | 12.5 | 11.5 | 11.5 |
| Methionine, g/kg | 7.0 | 7.0 | 6.2 | 6.8 | 6.4 | 6.6 | 5.9 | 6.1 |
| Threonine, g/kg | 10.6 | 10.6 | 9.5 | 10.1 | 9.1 | 9.2 | 8.9 | 9.1 |
| Tryptophan, g/kg | 2.8 | 2.8 | 2.4 | 2.6 | 2.4 | 2.3 | 2.3 | 2.3 |
| Quantum Blue phytase, FTU/kg | 541 | 563 | - | 1390 | 522 | 544 | - | 1170 |
| Endogenous phytase, FTU/kg | 206 | 407 | 550 | 290 | 344 | 332 | 365 | 640 |
| Total phytase activity, FTU/kg | 747 | 970 | 550 | 1680 | 866 | 876 | 365 | 1810 |
| PC1 | PC2 | NC | NC+ | Sem | p | |
|---|---|---|---|---|---|---|
| Weight day 28, g | 1910 c | 1838 b | 1656 a | 1848 b | 22.7 | <0.001 |
| Weight day 42, g | 3447 c | 3112 b | 2948 a | 3114 b | 53.8 | <0.001 |
| Days to reach target weight (2.2 kg) | 27.2 a | 30.1 b | 31.9 b | 30.2 b | 0.74 | <0.001 |
| Starter/Grower (days 0–28) | ||||||
| FI, g | 2400 c | 2258 b | 2089 a | 2248 b | 43.53 | <0.001 |
| BWG, g | 1870 c | 1798 b | 1616 a | 1808 bc | 22.74 | <0.001 |
| FCR, g/g | 1.28 | 1.26 | 1.29 | 1.24 | 0.020 | 0.297 |
| Finisher (days 28–42) | ||||||
| FI, g | 2232 b | 2045 ab | 1983 a | 2062 ab | 59.1 | 0.036 |
| BWG, g | 1537 b | 1274 a | 1292 a | 1266 a | 42.2 | <0.001 |
| FCR, g/g | 1.46 | 1.61 | 1.54 | 1.64 | 0.052 | 0.076 |
| Overall (days 0–42) | ||||||
| FI, g | 4633 c | 4327 b | 4072 a | 4309 ab | 79.7 | <0.001 |
| BWG, g | 3407 c | 3071 b | 2908 a | 3074 b | 53.8 | <0.001 |
| FCR, g/g | 1.36 | 1.41 | 1.40 | 1.40 | 0.019 | 0.277 |
| Whole-body DXA | ||||||
| BMD, g/cm2 | 0.43 b | 0.40 a | 0.39 a | 0.39 a | 0.004 | 0.003 |
| BMC, g | 93.16 c | 80.48 b | 75.77 a | 81.14 b | 1.05 | <0.001 |
| Bone mass, g | 219 d | 204 b | 197 a | 211 c | 2.0 | 0.007 |
| Lean mass, g | 2136 c | 1879 b | 1791 a | 1849 ab | 24.97 | <0.001 |
| Fat mass (g) | 1068 c | 943 a | 935 a | 994 b | 13.8 | 0.003 |
| Tibia Measurements | PC1 | PC2 | NC | NC+ | SEM | p |
|---|---|---|---|---|---|---|
| Weight, g | 25.3 | 23.7 | 24.0 | 24.6 | 0.74 | 0.544 |
| Diameter, mm | 8.83 | 8.60 | 8.88 | 8.80 | 0.204 | 0.710 |
| Length, mm | 110.0 a | 113.5 b | 113.7 b | 112.2 b | 0.879 | 0.046 |
| Max load, kg | 42.5 | 40.5 | 41.3 | 40.3 | 2.38 | 0.930 |
| Bone mineral density, g/cm2 * | 0.33 | 0.35 | 0.33 | 0.37 | 0.009 | 0.498 |
| Bone mineral content, g * | 0.60 | 0.67 | 0.62 | 0.70 | 0.02 | 0.369 |
| Apparent Total Tract Digestibility (ATTD), % | PC1 | PC2 | NC | NC+ | SEM | p |
|---|---|---|---|---|---|---|
| Dry matter | 74.9 a | 74.6 a | 74.2 a | 76.4 b | 0.40 | 0.002 |
| Gross energy | 78.0 ab | 77.8 a | 77.0 a | 79.0 b | 0.35 | 0.004 |
| Ash | 45.9 | 44.9 | 45.6 | 47.3 | 0.67 | 0.088 |
| Oil B | 82.7 | 83.5 | 80.3 | 81.6 | 0.86 | 0.056 |
| Neutral detergent fibre | 55.5 a | 57.4 ab | 58.5 bc | 59.7 c | 0.67 | <0.001 |
| Calcium | 50.4 b | 47.6 a | 48.2 ab | 54.2 c | 0.88 | <0.001 |
| Potassium | 41.0 a | 40.9 a | 41.6 a | 43.4 b | 0.58 | 0.014 |
| Magnesium | 33.2 a | 33.9 a | 33.6 a | 38.4 b | 0.53 | <0.001 |
| Nitrogen | 70.2 | 70.1 | 69.6 | 70.8 | 0.79 | 0.786 |
| Phosphorus | 76.2 | 76.5 | 75.7 | 78.2 | 0.01 | 0.400 |
| Apparent ileal digestibility (AID), % | ||||||
| Dry matter | 72.9 | 68.2 | 70.3 | 70.8 | 1.01 | 0.757 |
| Ash | 48.8 | 47.4 | 48.0 | 49.6 | 1.71 | 0.973 |
| Ileal concentrations, nmol/g | ||||||
| IP6 | 36,275 c | 25,674 b | 39,895 c | 8539 a | 2803.7 | <0.001 |
| IP5 | 11,383 c | 8033 b | 6969 b | 3718 a | 622.5 | <0.001 |
| IP4 | 5530 b | 5033 b | 2450 a | 5000 b | 366.6 | 0.009 |
| IP3 | 920 b | 1005 b | 220 a | 1226 b | 122.7 | 0.01 |
| IP2 | 460 c | 377 c | 138 a | 251 b | 35.8 | 0.009 |
| IP sum * | 54,453 b | 40,067 b | 49,602 b | 18,639 a | 5280 | <0.001 |
| Inositol | 1083 a | 2594 a | 3434 a | 6504 b | 532.2 | <0.001 |
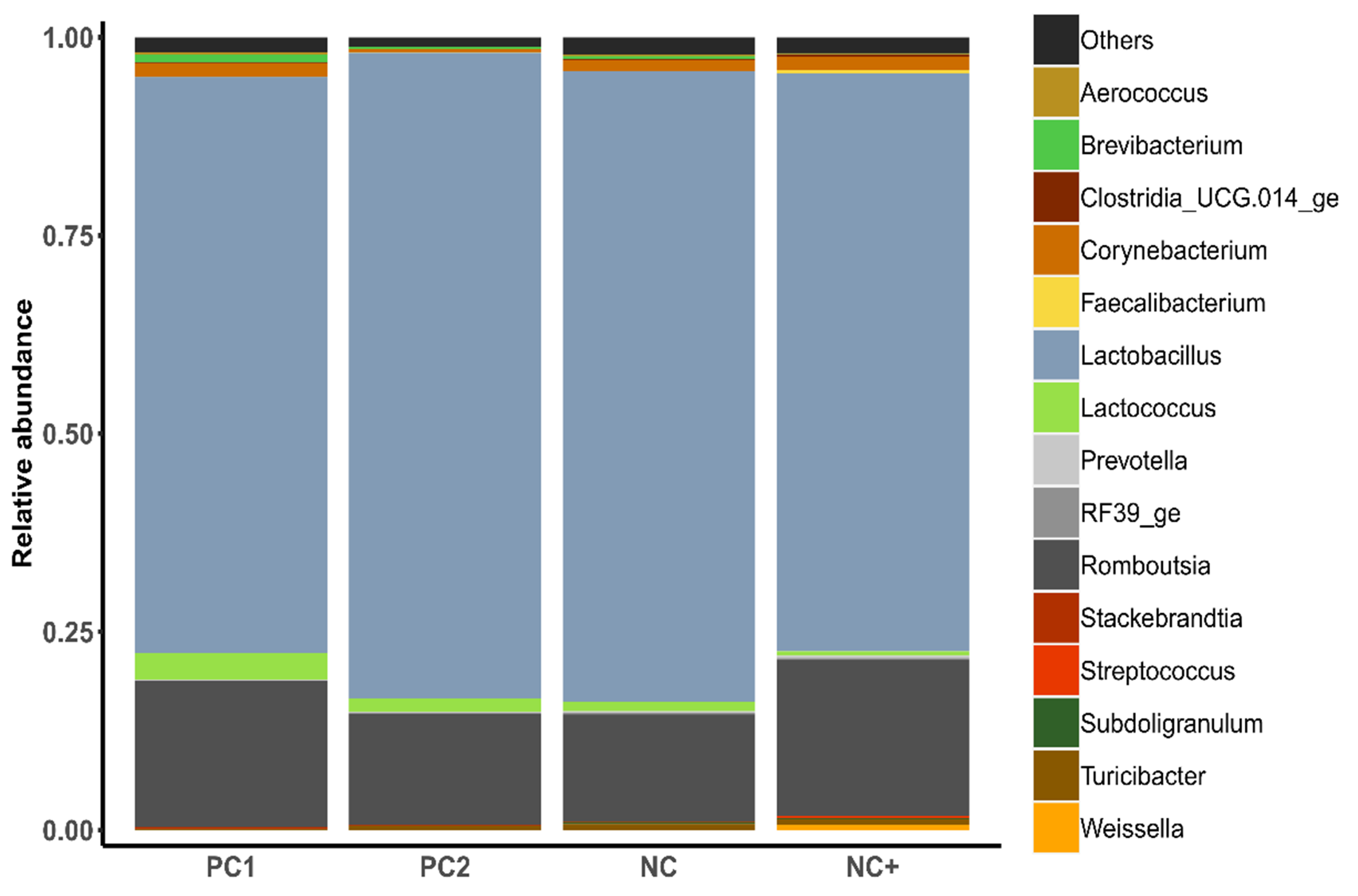
| Relative Abundance of Genera (Phylum), % | PC1 | PC2 | NC | NC+ | SEM | p |
|---|---|---|---|---|---|---|
| Weissella (Firmicutes) | 0.006 a | 0.005 a | 0.004 a | 0.398 b | 0.041 | <0.001 |
| Turicibacter (Firmicutes) | 0.064 a | 0.421 b | 0.379 b | 0.471 b | 0.060 | 0.002 |
| Clostridium sensu stricto 1 (Firmicutes) | 0.006 a | 0.038 b | 0.057 b | 0.053 b | 0.007 | 0.002 |
| Stackebrandtia (Actinobacteriota) | 0.167 ab | 0.055 b | 0.041 ab | 0.014 a | 0.021 | 0.030 |
| Alloprevotella (Bacteroidota) | 0.001 a | 0.017 b | 0.027 b | 0.046 ab | 0.006 | 0.038 |
| Brevibacterium (Actinobacteriota) | 0.667 ab | 0.226 b | 0.227 b | 0.061 a | 0.092 | 0.041 |
| PC1 | PC2 | NC | NC+ | SEM | p | |
|---|---|---|---|---|---|---|
| N balance | ||||||
| Starter/Grower (0–28 d) | ||||||
| N intake (g/day) | 2.86 b | 2.84 b | 2.48 a | 2.74 b | 0.074 | <0.001 |
| N retention (g/day) | 1.92 b | 1.85 b | 1.66 a | 1.86 b | 0.033 | <0.001 |
| N excretion (g/day) | 0.94 | 0.99 | 0.82 | 0.88 | 0.062 | 0.052 |
| N excretion (g/kg gain) | 14.07 | 15.5 | 14.17 | 13.67 | 0.956 | 0.266 |
| Finisher (29–42 d) | ||||||
| N intake (g/day) | 5.08 | 4.85 | 4.56 | 4.76 | 0.198 | 0.090 |
| N retention (g/day) | 3.16 b | 2.62 a | 2.66 a | 2.60 a | 0.123 | <0.001 |
| N excretion (g/day) | 1.92 | 2.23 | 1.90 | 2.16 | 0.176 | 0.169 |
| N excretion (g/kg gain) | 17.57 a | 24.61 b | 20.72 ab | 24.25 ab | 2.294 | 0.015 |
| Overall (0–42 d) | ||||||
| N intake (g/day) | 3.60 b | 3.49 b | 3.17 a | 3.41 ab | 0.088 | <0.001 |
| N retention (g/day) | 2.34 b | 2.11 a | 1.99 a | 2.11 a | 0.052 | <0.001 |
| N excretion (g/day) | 1.26 ab | 1.38 b | 1.18 a | 1.30 ab | 0.065 | 0.032 |
| N excretion (g/kg gain) | 15.58 a | 18.89 b | 17.02 ab | 17.81 ab | 0.918 | 0.010 |
| N excretion to reach target weight of 2.2 kg (g) * | 25.52 | 29.93 | 25.99 | 26.48 | 1.774 | 0.07 |
| P excretion (g/day) | 0.44 | 0.43 | 0.42 | 0.38 | 0.029 | 0.208 |
| P excretion 0–28 d (g/kg gain) | 6.57 ab | 6.67 ab | 7.29 b | 5.86 a | 0.46 | 0.033 |
| P excretion 29–42 d (g/kg gain) | 4.00 | 4.70 | 4.56 | 4.19 | 0.31 | 0.097 |
| P excretion 0–42 d (g/kg gain) | 5.41 | 5.86 | 6.08 | 5.17 | 0.393 | 0.109 |
| Total P excretion to reach target weight of 2.2 kg (g) * | 11.9 | 12.9 | 13.4 | 11.4 | 0.865 | 0.109 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mulvenna, C.C.; McCormack, U.M.; Magowan, E.; McKillen, J.; Bedford, M.R.; Walk, C.L.; Oster, M.; Reyer, H.; Wimmers, K.; Fornara, D.A.; et al. The Growth Performance, Nutrient Digestibility, Gut Bacteria and Bone Strength of Broilers Offered Alternative, Sustainable Diets Varying in Nutrient Specification and Phytase Dose. Animals 2022, 12, 1669. https://doi.org/10.3390/ani12131669
Mulvenna CC, McCormack UM, Magowan E, McKillen J, Bedford MR, Walk CL, Oster M, Reyer H, Wimmers K, Fornara DA, et al. The Growth Performance, Nutrient Digestibility, Gut Bacteria and Bone Strength of Broilers Offered Alternative, Sustainable Diets Varying in Nutrient Specification and Phytase Dose. Animals. 2022; 12(13):1669. https://doi.org/10.3390/ani12131669
Chicago/Turabian StyleMulvenna, Christina C., Ursula M. McCormack, Elizabeth Magowan, John McKillen, Mike R. Bedford, Carrie L. Walk, Michael Oster, Henry Reyer, Klaus Wimmers, Dario A. Fornara, and et al. 2022. "The Growth Performance, Nutrient Digestibility, Gut Bacteria and Bone Strength of Broilers Offered Alternative, Sustainable Diets Varying in Nutrient Specification and Phytase Dose" Animals 12, no. 13: 1669. https://doi.org/10.3390/ani12131669
APA StyleMulvenna, C. C., McCormack, U. M., Magowan, E., McKillen, J., Bedford, M. R., Walk, C. L., Oster, M., Reyer, H., Wimmers, K., Fornara, D. A., & Ball, M. E. E. (2022). The Growth Performance, Nutrient Digestibility, Gut Bacteria and Bone Strength of Broilers Offered Alternative, Sustainable Diets Varying in Nutrient Specification and Phytase Dose. Animals, 12(13), 1669. https://doi.org/10.3390/ani12131669