
Fatty Acid Content of Four Salmonid Fish Consumed by Indigenous Peoples from the Yamal-Nenets Autonomous Okrug (Northwestern Siberia, Russia)

 ,
, 
 and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling
2.2. Biochemical Assay
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gritsenko, D.; Efimova, E. Planning for a sustainable Arctic: Regional development in the Yamalo-Nenets Autonomous Okrug (Russia). In Russia’s Far North: The Contested Energy Frontier; Tynkkynen, V.-P., Tabata, S., Gritsenko, D., Goto, M., Eds.; Routledge: New York, NY, USA, 2018; pp. 67–83. [Google Scholar]
- Andronov, S.; Lobanov, A.; Popov, A.; Luo, Y.; Shaduyko, O.; Fesyun, A.; Lobanova, L.; Bogdanova, E.; Kobel’kova, I. Changing diets and traditional lifestyle of Siberian Arctic Indigenous Peoples and effects on health and well-being. Ambio 2021, 50, 2060–2071. [Google Scholar] [CrossRef] [PubMed]
- Lobanova, L.P.; Lobanov, A.A.; Popov, A.I. Transformation of the diet of the indigenous Nenets population. Sci. Bull. Yamalo-Nenets Auton. Okrug 2013, 4, 24–25. (In Russian) [Google Scholar]
- Bogdanov, V.D.; Melnichenko, I.P. Characteristics of the ichthyofauna of the Yamal Peninsula (the Yamal-Nenets autonomous district). Fauna Ural. Sib. 2016, 1, 105–113. (In Russian) [Google Scholar]
- Isakov, P.V.; Selyukov, A.G. Whitefish in the Ecosystem of the Gulf of Ob; Tyumen State University Publishing House: Tyumen, Russia, 2010. (In Russian) [Google Scholar]
- Darinov, E.N. Materials Explaining the Total Allowable Catches of Aquatic Biological Resources in the Inland Waters of the Tyumen Region, Including the Yamalo-Nenets and Khanty-Mansi Autonomous Districts, with the Exception of Inland Sea Waters, for 2022 (with Environmental Impact Assessment); Tyumen Branch of VNIRO: Tyumen, Russia, 2021. (In Russian) [Google Scholar]
- Melnichenko, I.P.; Bogdanov, V.D. The status of fish stocks and prospects of fishing in the reservoirs of the Yamal district of the Yamalo-Nenets Autonomous Okrug. Sci. Bull. Yamalo-Nenets Auton. Okrug 2009, 1, 47–61. (In Russian) [Google Scholar]
- Gladyshev, M.I.; Sushchik, N.N.; Makhutova, O.N.; Glushchenko, L.A.; Rudchenko, A.E.; Makhrov, A.A.; Borovikova, E.A.; Dgebuadze, Y.Y. Fatty acid composition and contents of seven commercial fish species of genus Coregonus from Russian subarctic water bodies. Lipids 2017, 52, 1033–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sushchik, N.N.; Makhutova, O.N.; Rudchenko, A.E.; Glushchenko, L.A.; Shulepina, S.P.; Kolmakova, A.A.; Gladyshev, M.I. Comparison of fatty acid contents in major lipid classes of seven salmonid species from Siberian Arctic lakes. Biomolecules 2020, 10, 419. [Google Scholar] [CrossRef] [Green Version]
- Kalyakin, V.N.; Romanenko, F.A.; Molochaev, A.V.; Rogacheva, E.V.; Syroechkovsky, E.E. Gydansky Nature Reserve. Reserves of Russia. Nature Reserves of Siberia. II; Logata: Moscow, Russia, 2000. (In Russian) [Google Scholar]
- Folch, J.; Less, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Bichkaeva, F.A.; Baranova, N.G.; Vlasova, O.S.; Nesterova, E.V.; Bichkaev, A.A.; Shengof, B.A.; Tretyakova, T.V. Method for Measuring Mass Concentration of Methyl Esters of Fatty Acids in Biological Media by Gas-Liquid Chromatography Method. Patent 2 758 932(13) C1, 3 November 2021. [Google Scholar]
- Dvoretsky, A.G.; Bichkaeva, F.A.; Baranova, N.F.; Dvoretsky, V.G. Fatty acid composition of the Barents Sea red king crab (Paralithodes camtschaticus) leg meat. J. Food Compos. Anal. 2021, 98, 103826. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Bichkaeva, F.A.; Baranova, N.F.; Dvoretsky, V.G. Fatty acids in the circulatory system of an invasive king crab from the Barents Sea. J. Food Compos. Anal. 2022, 110, 104528. [Google Scholar] [CrossRef]
- Chen, J.; Liu, H. Nutritional indices for assessing fatty acids: A mini-review. Int. J. Mol. Sci. 2020, 21, 5695. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest of southern Wisconsin. Ecol. Monogr. 1957, 27, 225–349. [Google Scholar] [CrossRef]
- Clarke, K.R.; Somerfield, P.J.; Chapman, M.G. On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray–Curtis coefficient for denuded assemblages. J. Exp. Mar. Biol. Ecol. 2006, 330, 55–80. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; Primer-E.; Plymouth Marine Laboratory: Plymouth, UK, 2001. [Google Scholar]
- Yoshii, K.; Melnik, N.G.; Timoshkin, O.A.; Bondarenko, N.A.; Anoshko, N.; Yoshioka, T.; Wada, E. Stable isotope analyses of the pelagic food web in Lake Baikal. Limnol. Oceanogr. 1999, 44, 502–511. [Google Scholar] [CrossRef] [Green Version]
- Nikiforova, A.; Zamaratskaia, G.; Pickova, J. Fatty acid composition of salted and fermented products from Baikal omul (Coregonus autumnalis migratorius). J. Food Sci. Technol. 2020, 57, 595–605. [Google Scholar] [CrossRef]
- Gladyshev, M.I.; Makhrov, A.A.; Baydarov, I.V.; Safonova, S.S.; Golod, V.M.; Alekseyev, S.S.; Glushchenko, L.A.; Rudchenko, A.E.; Karpov, V.A.; Sushchik, N.N. Fatty acid composition and contents of fish of genus Salvelinus from natural ecosystems and aquaculture. Biomolecules 2022, 12, 144. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, R.; Klemetsen, A.; Alekseyev, S.; Adams, C.E.; Power, M. The role of Salvelinus in contemporary studies of evolution, trophic ecology and anthropogenic change. Hydrobiologia 2016, 783, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Bell, M.V.; Henderson, R.J.; Sargent, J.R. The role of polyunsaturated fatty acids in fish. Comp. Biochem. Physiol. B 1986, 83, 711–719. [Google Scholar] [CrossRef]
- Arts, M.T.; Kohler, C.C. Health and condition in fish: The influence of lipids on membrane competency and immune response? In Lipids in Aquatic Ecosystems; Arts, M.T., Kainz, M., Brett, M.T., Eds.; Springer: New York, NY, USA, 2009. [Google Scholar]
- Hazel, J.R. Effects of temperature on the structure and metabolism of cell membranes in fish. Am. J. Physiol. 1984, 246, 460–470. [Google Scholar] [CrossRef]
- Farkas, T.; Csengeria, I.; Majorosa, F.; Oláha, J. Metabolism of fatty acids in fish: III. Combined effect of environmental temperature and diet on formation and deposition of fatty acids in the carp, Cyprinus carpio Linnaeus 1758. Aquaculture 1980, 20, 29–40. [Google Scholar] [CrossRef]
- Arts, M.T.; Palmer, M.E.; Skiftesvik, A.B.; Jokinen, I.E.; Browman, H.I. UVB radiation variably affects n-3 fatty acids but elevated temperature reduces n-3 fatty acids in juvenile Atlantic salmon (Salmo salar). Lipids 2012, 47, 1181–1192. [Google Scholar] [CrossRef] [PubMed]
- Wijekoon, M.; Parrish, C.C.; Mansour, A. Effect of growth temperature on muscle lipid class and fatty acid composition in adult steelhead trout (Oncorhynchus mykiss) fed commercial diets with different ω6 to ω3 fatty acid ratios. J. Aquac. Res. Dev. 2021, 12, 643. [Google Scholar]
- Mzengereza, K.; Ishikawa, M.; Koshio, S.; Yokoyama, S.; Yukun, Z.; Shadrack, R.S.; Seo, S.; Duy Khoa, T.N.; Moss, A.; Dossou, S.; et al. Effect of substituting fish oil with camelina oil on growth performance, fatty acid profile, digestibility, liver histology, and antioxidative status of red seabream (Pagrus major). Animals 2021, 11, 1990. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, S.L.; Ghirmay, A.; Gong, Y.; Dahle, D.; Vasanth, G.; Sørensen, M.; Kiron, V. Growth, chemical composition, histology and antioxidant genes of Atlantic salmon (Salmo salar) fed whole or pre-processed Nannochloropsis oceanica and Tetraselmis sp. Fishes 2021, 6, 23. [Google Scholar] [CrossRef]
- García-Márquez, J.; Galafat, A.; Alarcón, F.J.; Figueroa, F.L.; Martínez-Manzanares, E.; Arijo, S.; Abdala-Díaz, R.T. Cultivated and wild juvenile thick-lipped grey mullet, Chelon labrosus: A comparison from a nutritional point of view. Animals 2021, 11, 2112. [Google Scholar] [CrossRef]
- Batista, R.O.; Nobrega, R.O.; Schleder, D.D.; Pettigrew, J.E.; Fracalossi, D.M. Aurantiochytrium sp. meal improved body fatty acid profile and morphophysiology in Nile tilapia reared at low temperature. Fishes 2021, 6, 45. [Google Scholar] [CrossRef]
- Popov, P.A. Fish of Siberia: Distribution, Ecology, Catch; Novosibirsk State University: Novosibirsk, Russia, 2007. (In Russian) [Google Scholar]
- Stepanova, V.B.; Korshunov, S.A. On omul feeding in the Gulf of Gydan. Vestn. Ecol. For. Landsc. Stud. 2007, 8, 131–134. (In Russian) [Google Scholar]
- Vyshegorodcev, A.A. Fishes of the Yenisei; Nauka: Novosibirsk, Russia, 2000. (In Russian) [Google Scholar]
- Stepanova, V.B.; Stepanov, S.I. Importance of relict crustaceous in whitefish nutrition during under the ice period. Vestn. Ecol. For. Landsc. Stud. 2006, 6, 142–145. (In Russian) [Google Scholar]
- Henderson, R.J. Fatty acid metabolism in freshwater fish with particular reference to polyunsaturated fatty acids. Arch. Tierernaehrung 1996, 49, 5–22. [Google Scholar] [CrossRef]
- Bordignon, F.; Tomás-Vidal, A.; Trocino, A.; Milián Sorribes, M.C.; Jover-Cerdá, M.; Martínez-Llorens, S. Fatty acid signatures in different tissues of mediterranean yellowtail, Seriola dumerili (Risso, 1810), fed diets containing different levels of vegetable and fish oils. Animals 2020, 10, 198. [Google Scholar] [CrossRef] [Green Version]
- Happel, A.; Jonas, J.L.; McKenna, P.R.; Rinchard, J.; He, J.X.; Czesny, S.J. Spatial variability of lake trout diets in Lakes Huron and Michigan revealed by stomach content and fatty acid profiles. Can. J. Fish. Aquat. Sci. 2018, 75, 95–105. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Cucumaria in Russian waters of the Barents Sea: Biological aspects and aquaculture potential. Front. Mar. Sci. 2021, 8, 613453. [Google Scholar] [CrossRef]
- Nikonova, A.A.; Shishlyannikov, S.M.; Volokitina, N.A.; Galachyants, Y.P.; Bukin, Y.S.; Blinov, V.V.; Gnatovsky, R.Y.; Vorobyeva, S.S. Fatty acid changes in nearshore phytoplankton under anthropogenic impact as a biodiversity risk factor for the world’s deepest lake Baikal. Diversity 2022, 14, 55. [Google Scholar] [CrossRef]
- Uauy, R.; Dangour, A.D. Nutrition in brain development and aging: Role of essential fatty acids. Nutr. Rev. 2006, 64, S24–S33. [Google Scholar] [CrossRef]
- Bork, C.; Mortensen, L.; Hjelmgaard, K.; Schmidt, E. Marine n-3 fatty acids and CVD: New insights from recent follow-up studies and clinical supplementation trials. Proc. Nutr. Soc. 2020, 79, 428–434. [Google Scholar] [CrossRef]
- Saccà, S.C.; Cutolo, C.A.; Ferrari, D.; Corazza, P.; Traverso, C.E. The eye, oxidative damage and polyunsaturated fatty acids. Nutrients 2018, 10, 668. [Google Scholar] [CrossRef] [Green Version]
- Hurst, S.; Zainal, Z.; Caterson, B.; Hughes, C.E.; Harwood, J.L. Dietary fatty acids and arthritis. Prostaglandins Leukot. Essent. Fat. Acids PLEFA 2010, 82, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes. Nutrients 2010, 2, 355–374. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, T.S.; Giromini, C.; Rebucci, R.; Baldi, A. Omega-3 polyunsaturated fatty acids counteract inflammatory and oxidative damage of non-transformed porcine enterocytes. Animals 2020, 10, 956. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Maekawa, M.; Katakura, M.; Hamazaki, K.; Matsuoka, Y. Possibility of polyunsaturated fatty acids for the prevention and treatment of neuropsychiatric illnesses. J. Pharmacol. Sci. 2014, 124, 294–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendel, M.; Heller, A.R. Anticancer actions of omega-3 fatty acids—Current state and future perspectives. Curr. Med. Chem. Anticancer Agents 2009, 9, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Andronov, S.V.; Lobanov, A.A.; Kochkin, R.A.; Bogdanova, E.N.; Popov, A.I.; Lobanova, L.P.; Lobanova, E.A.; Protasova, I.V. The role of traditional nutrition in preserving the health of indigenous population in the Arctic zone of Western Siberia. In Conservation of the Indigenous Population in the Arctic Zone of the Russian Federation under Conditions of Lifestyle Transformation and Climate Change; Bogdanova, E.N., Zadorin, M.Y., Eds.; KIRA: Arkhangelsk, Russia, 2018; pp. 142–144. (In Russian) [Google Scholar]
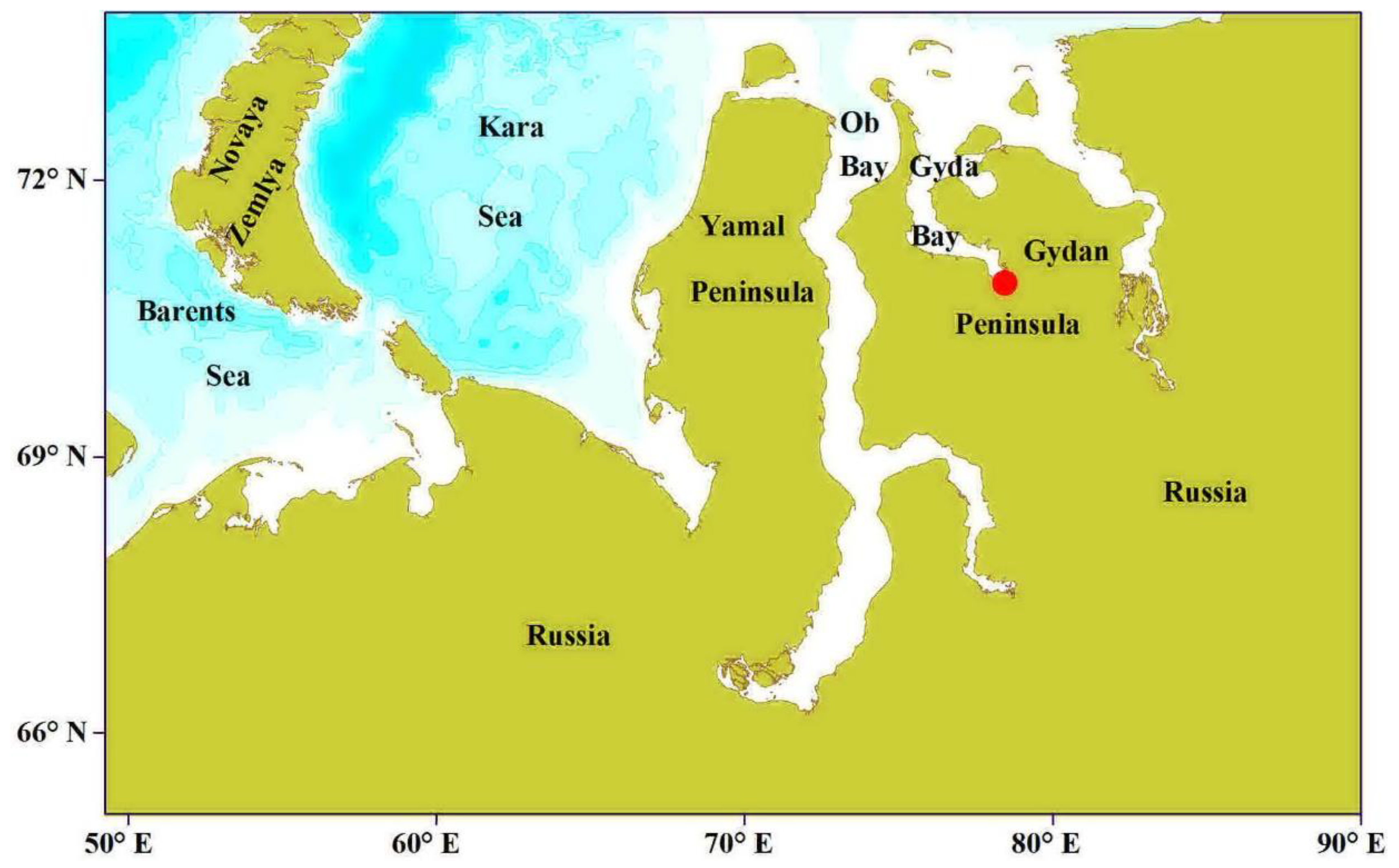

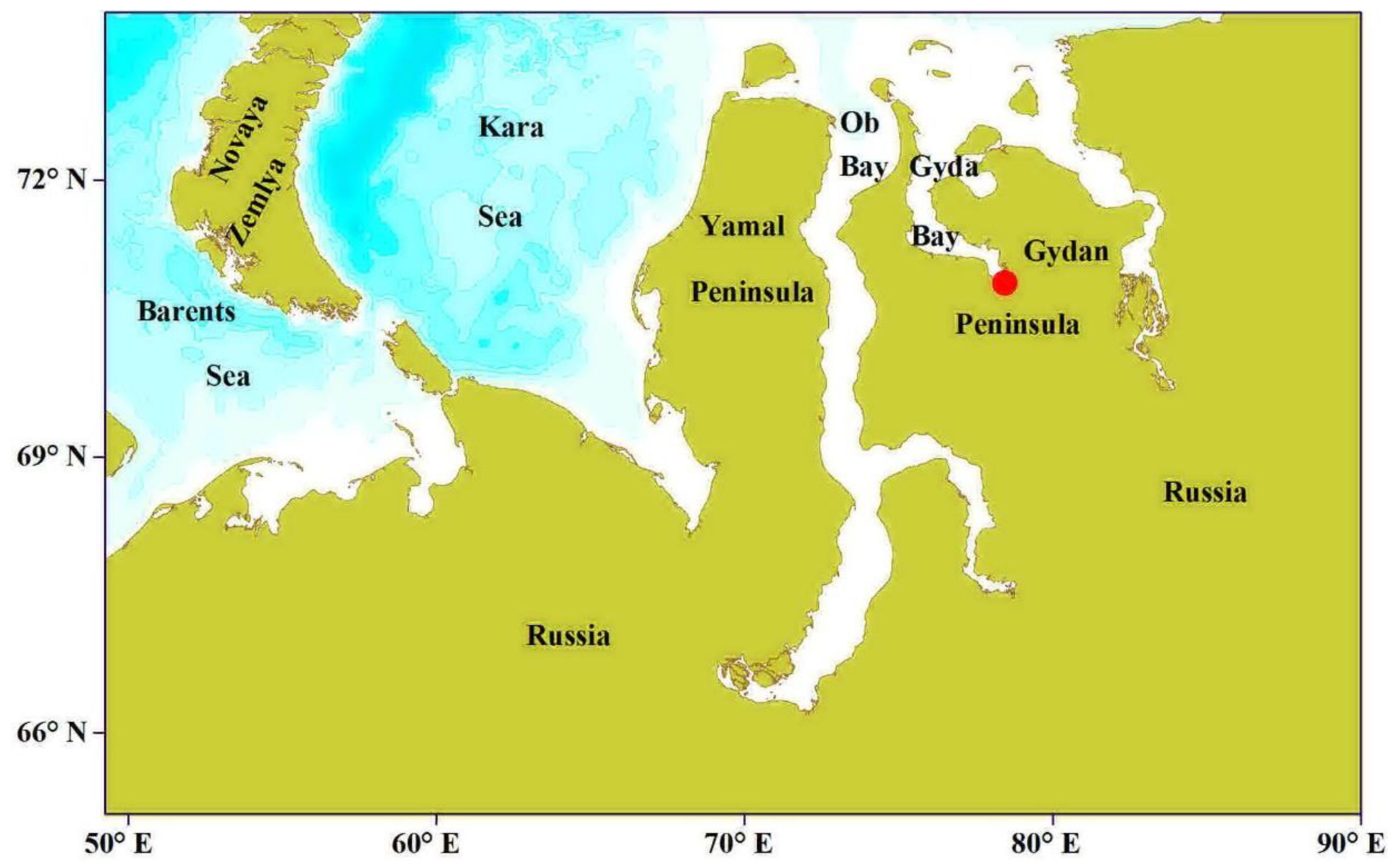{kind=link}
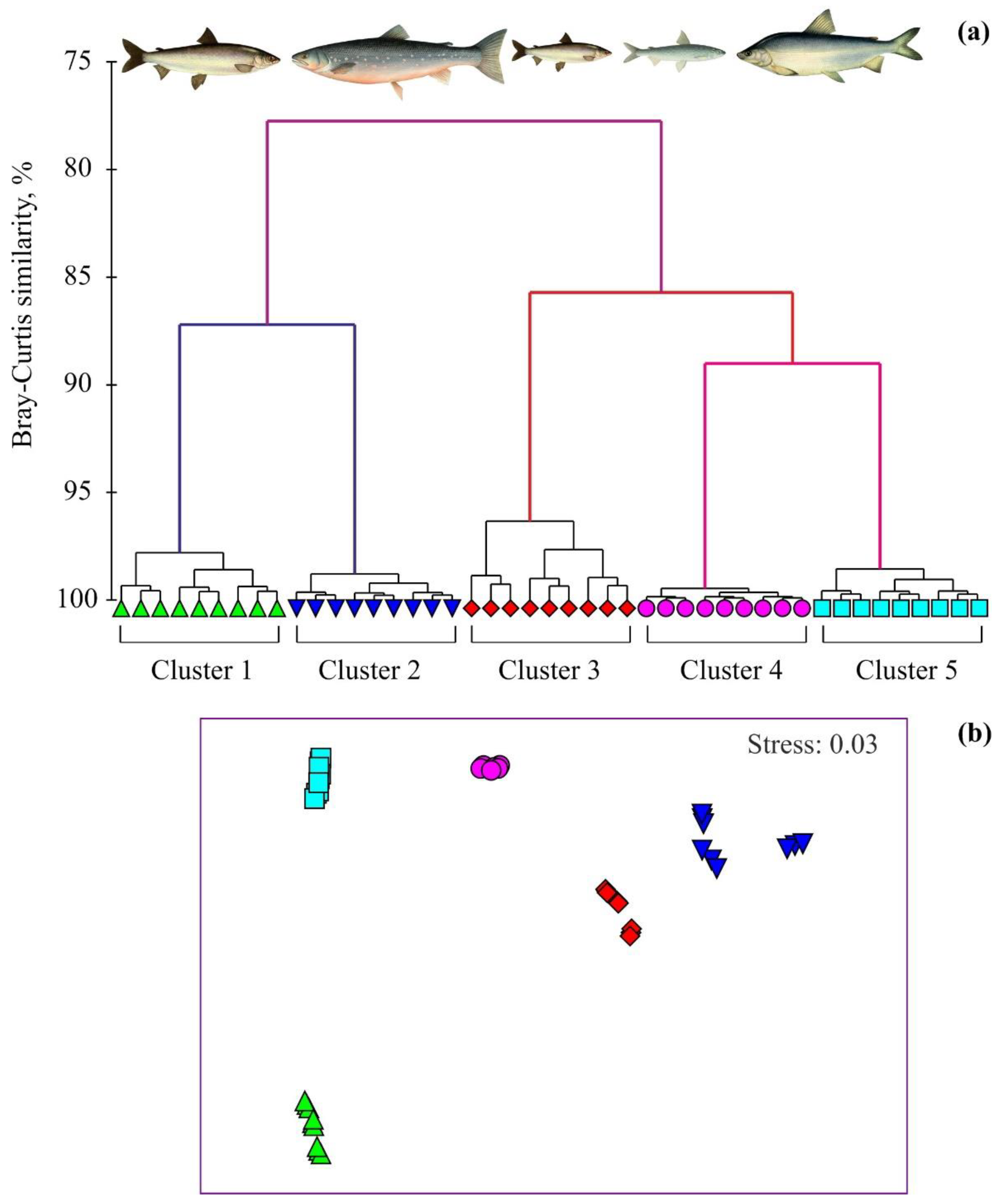
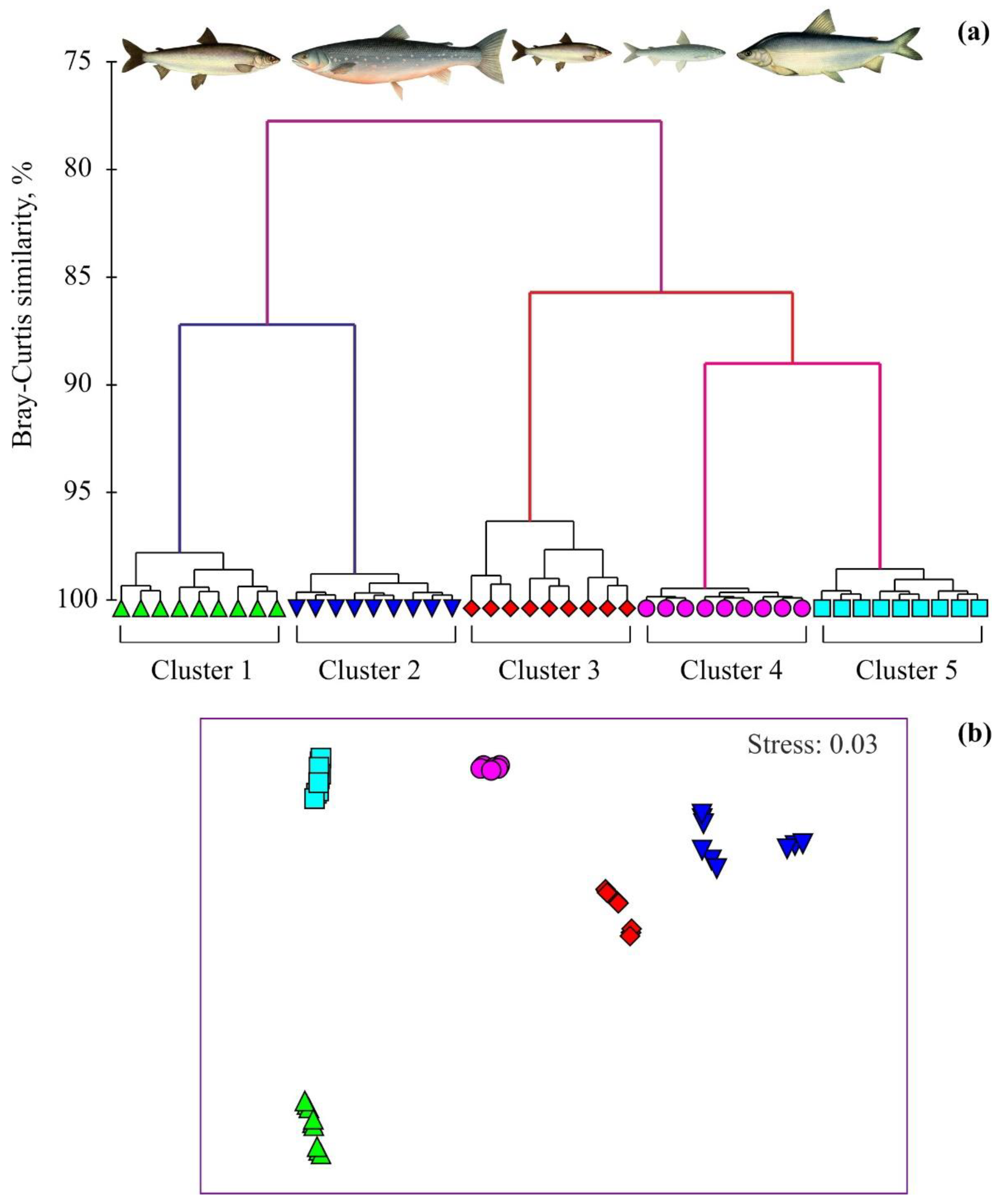{kind=link}
| Species Name | Common Name | Water Body | N | Month | L, cm | W, kg |
|---|---|---|---|---|---|---|
| Coregonus autumnalis (adult) | Arctic cisco | Gyda River | 9 | August | 25–29 | 1.35–1.6 |
| Coregonus autumnalis (juvenile) | Arctic cisco | Gyda River | 9 | August | 11–12 | 0.22–0.24 |
| Coregonus sardinella | Least cisco | Gyda River | 9 | August | 12 | 0.14–0.15 |
| Coregonus muksun | Muksun | Gyda River, Gyda Bay | 9 | September | 48–51 | 1.45–1.5 |
| Salvelinus alpinus | Arctic charr | Gyda River | 9 | September | 39 | 1.35–1.41 |
| Fatty Acid | Concentration, μg g−1 | % of Total Fatty Acids | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CAA | CAJ | CS | CM | SA | CAA | CAJ | CS | CM | SA | |
| C6:0 | 0.3 a | 0.9 a | 5.5 b | 4.7 b | 6.5 c | <0.01 a | 0.01 b | 0.05 c | 0.03 d | 0.02 e |
| C8:0 | 1.4 a | 5.4 b | 5.6 b | 6.2 b | 5.9 b | 0.01 a | 0.07 b | 0.05 c | 0.04 c | 0.02 d |
| C9:0 | 1.5 a | 3.6 a | 3.4 a | 3.9 ab | 6.8 b | 0.01 a | 0.05 b | 0.03 c | 0.03 c | 0.02 ac |
| C10:0 | 2.9 a | 7.5 b | 7.4 b | 8.2 b | 11.5 c | 0.01 a | 0.10 b | 0.07 c | 0.06 c | 0.04 d |
| C11:0 | 1.2 a | 1.2 a | 1.1 a | 1.0 a | 2.8 b | 0.01 a | 0.02 b | 0.01 a | 0.01 a | 0.01 a |
| C12:0 | 75.2 a | 52.6 b | 63.4 c | 69.8 a | 71.1 a | 0.37 a | 0.70 b | 0.56 c | 0.51 c | 0.26 d |
| C13:0 | 37.8 a | 54.3 b | 55.1 b | 68.3 c | 55.1 b | 0.19 a | 0.73 b | 0.49 ab | 0.49 ab | 0.20 a |
| C14:0 | 1190 a | 206 b | 418 c | 758 d | 1050 e | 5.89 a | 2.76 b | 3.69 c | 5.48 a | 3.78 c |
| C15:0 | 133 a | 61.3 b | 73.8 c | 108 a | 133 a | 0.66 a | 0.82 b | 0.65 a | 0.78 b | 0.48 c |
| C16:0 | 3550 a | 1720 b | 2280 c | 2970 d | 5400 e | 17.6 a | 23.1 b | 20.2 ab | 21.5 b | 19.5 a |
| C17:0 | 87.3 a | 37.7 b | 43.1 b | 89.7 a | 89.8 a | 0.43 ab | 0.50 a | 0.38 b | 0.65 c | 0.32 b |
| C18:0 | 547 a | 391 b | 556 a | 698 c | 1590 d | 2.72 a | 5.23 bc | 4.91 b | 5.05 b | 5.74 c |
| C20:0 | 34.0 a | 11.7 b | 11.7 b | 21.2 c | 42.0 d | 0.17 a | 0.16 a | 0.10 b | 0.15 a | 0.15 a |
| C21:0 | 3.1 a | 4.9 a | 10.8 b | 1.9 c | 2.8 a | 0.02 a | 0.07 b | 0.10 c | 0.01 a | 0.01 a |
| C22:0 | 12.9 a | 55.0 b | 30.3 c | 53.1 b | 47.6 b | 0.06 a | 0.73 b | 0.27 c | 0.38 d | 0.17 e |
| C23:0 | 3.0 a | 2.1 a | 1.9 a | 3.1 a | 11.9 b | 0.01 a | 0.03 b | 0.02 a | 0.02 ab | 0.04 c |
| C24:0 | 5.4 a | 5.3 a | 4.5 a | 8.8 b | 9.0 b | 0.03 a | 0.07 b | 0.04 a | 0.06 b | 0.03 a |
| C14:1n5t | 0.6 | - | - | - | - | <0.01 | - | - | - | - |
| C14:1n5c | 26.8 a | 6.7 b | 9.3 c | 13.3 d | 31.8 e | 0.13 a | 0.09 b | 0.08 b | 0.10 b | 0.11 c |
| C15:1 | 1.4 a | 0.5 b | 0.7 c | 0.5 b | 0.5 b | 0.01 a | 0.01 a | 0.01 a | <0.01 b | <0.01 b |
| C16:1n7t | 42.7 a | 0.8 b | 0.8 b | 0.8 b | - | 0.21 a | 0.01 b | 0.01 b | 0.01 b | - |
| C16:1n7c | 2670 a | 350 b | 942 c | 1370 d | 2810 a | 13.3 a | 4.69 b | 8.32 c | 9.94 d | 10.2 e |
| C17:1 | 25.1 a | 1.0 b | 2.0 b | 2.1 b | 2.9 b | 0.12 a | 0.01 bc | 0.02 c | 0.02 c | 0.01 b |
| C18:1n9t | 14.8 a | 13.0 a | 12.4 a | 31.7 b | 34.6 b | 0.07 a | 0.17 b | 0.11 c | 0.23 d | 0.12 c |
| C18:1n9c | 3750 a | 850 b | 1390 c | 2460 d | 6340 e | 18.6 a | 11.4 b | 12.3 b | 17.8 a | 22.9 c |
| C20:1 | 262 a | 48.9 b | 82.0 c | 157 d | 324 e | 1.30 a | 0.65 b | 0.72 b | 1.14 c | 1.17 c |
| C22:1 | 21.9 a | 3.1 b | 7.0 b | 26.2 a | 44.8 c | 0.11 a | 0.04 b | 0.06 b | 0.19 c | 0.16 d |
| C24:1 | 49.0 a | 10.0 b | 20.8 c | 80.4 d | 125.7 e | 0.24 a | 0.13 b | 0.18 c | 0.58 d | 0.45 e |
| C18:2n6t | 4.8 a | 1.0 b | - | - | - | 0.02 a | 0.01 b | - | - | - |
| C18:2n6 | 697 a | 168 b | 255 c | 224 b | 757 d | 3.46 a | 2.24 b | 2.26 b | 1.62 c | 2.73 d |
| C18:3n3 | 948 a | 179 b | 281 c | 170 b | 702 d | 4.71 a | 2.38 b | 2.48 b | 1.23 c | 2.53 b |
| C18:3n6 | 8.5 a | 19.7 a | 108.8 b | 100.9 b | 15.9 a | 0.04 a | 0.26 b | 0.96 c | 0.73 d | 0.06 a |
| C20:2n6 | 143 a | 38.0 b | 66.1 c | 39.4 b | 142 a | 0.71 a | 0.51 b | 0.58 c | 0.29 d | 0.51 b |
| C20:3n6 | 36.6 a | 9.2 b | 10.1 b | 30.0 c | 71.9 d | 0.18 a | 0.12 b | 0.09 c | 0.22 d | 0.26 e |
| C20:4n6 | 564 a | 260 b | 316 b | 371 bc | 669 d | 2.80 a | 3.46 b | 2.79 a | 2.69 a c | 2.41 c |
| C22:2n6 | 10.9 a | 3.4 b | 1.5 c | 6.1 d | 35.1 e | 0.05 a | 0.04 a | 0.01 b | 0.04 a | 0.13 c |
| C20:5n3 | 2110 a | 755 b | 1240 c | 1050 c | 1200 c | 10.5 a | 10.0 a | 10.9 a | 7.58 b | 4.34 c |
| C22:6n3 | 2480 a | 1900 a | 2680 b | 1960 a | 4310 c | 12.3 a | 25.2 b | 23.6 b | 14.2 a | 15.6 a |
| C20:3n3 | 95.1 a | 23.4 b | 35.2 c | 21.2 a | 155 d | 0.47 a | 0.31 b | 0.31 b | 0.15 c | 0.56 d |
| C22:4n6 | 36.5 a | 9.1 b | 11.7 b | 54.2 c | 150 d | 0.18 a | 0.12 b | 0.10 c | 0.39 d | 0.54 e |
| C22:3n3 | 2.1 a | 0.6 b | - | - | - | 0.01 a | 0.01 a | - | - | - |
| C22:5n6 | 189 a | 101 b | 132 b | 146 c | 448 d | 0.94 a | 1.34 b | 1.17 ab | 1.06 a | 1.62 c |
| C22:5n3 | 273 a | 129 b | 154 b | 635 c | 809 d | 1.36 a | 1.71 b | 1.36 a | 4.59 c | 2.92 d |
| ∑SFA | 5680 a | 2620 b | 3580 c | 4880 d | 8530 e | 28.2 a | 35.1 b | 31.6 ab | 35.3 b | 30.8 ab |
| ∑MUFA | 6870 a | 1280 b | 2470 c | 4150 d | 9720 e | 34.1 a | 17.2 b | 21.8 c | 30.0 d | 35.1 a |
| ∑PUFA | 7600 a | 3600 b | 5290 c | 4800 c | 9470 d | 37.7 a | 47.7 b | 46.7 b | 34.7 a | 34.2 a |
| Total | 20,200 a | 7510 b | 11,300 c | 13,800 d | 27,800 e | 100 | 100 | 100 | 100 | 100 |
| ∑n-3 | 5910 a | 2990 b | 4380 c | 3830 b | 7180 d | 29.3 a | 39.6 b | 38.7 b | 27.7 a | 25.9 a |
| ∑n-6 | 1690 a | 609 b | 901 c | 972 c | 2290 d | 8.39 a | 8.11 a | 7.96 a | 7.03 b | 8.26 a |
| n-6/n-3 | 0.29 a | 0.21 b | 0.21 b | 0.25 c | 0.32 a | - | - | - | - | - |
| n-3/n-6 | 3.50 a | 4.87 b | 4.87 b | 3.94 c | 3.14 a | - | - | - | - | - |
| AI | 0.58 ab | 0.54 a | 0.52 a | 0.68 b | 0.50 a | - | - | - | - | - |
| TI | 0.24 ac | 0.24 ac | 0.22 a | 0.32 bc | 0.29 c | - | - | - | - | - |
| h/H | 2.30 a | 2.23 a | 2.38 a | 1.89 b | 2.33 a | - | - | - | - | - |
| ∑PUFA/∑SFA | 1.34 ab | 1.38 ab | 1.48 b | 0.98 c | 1.11 a | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dvoretsky, A.G.; Bichkaeva, F.A.; Vlasova, O.S.; Andronov, S.V.; Dvoretsky, V.G. Fatty Acid Content of Four Salmonid Fish Consumed by Indigenous Peoples from the Yamal-Nenets Autonomous Okrug (Northwestern Siberia, Russia). Animals 2022, 12, 1643. https://doi.org/10.3390/ani12131643
Dvoretsky AG, Bichkaeva FA, Vlasova OS, Andronov SV, Dvoretsky VG. Fatty Acid Content of Four Salmonid Fish Consumed by Indigenous Peoples from the Yamal-Nenets Autonomous Okrug (Northwestern Siberia, Russia). Animals. 2022; 12(13):1643. https://doi.org/10.3390/ani12131643
Chicago/Turabian StyleDvoretsky, Alexander G., Fatima A. Bichkaeva, Olga S. Vlasova, Sergei V. Andronov, and Vladimir G. Dvoretsky. 2022. "Fatty Acid Content of Four Salmonid Fish Consumed by Indigenous Peoples from the Yamal-Nenets Autonomous Okrug (Northwestern Siberia, Russia)" Animals 12, no. 13: 1643. https://doi.org/10.3390/ani12131643
APA StyleDvoretsky, A. G., Bichkaeva, F. A., Vlasova, O. S., Andronov, S. V., & Dvoretsky, V. G. (2022). Fatty Acid Content of Four Salmonid Fish Consumed by Indigenous Peoples from the Yamal-Nenets Autonomous Okrug (Northwestern Siberia, Russia). Animals, 12(13), 1643. https://doi.org/10.3390/ani12131643