
Oilseed Supplementation Improves Milk Composition and Fatty Acid Profile of Cow Milk: A Meta-Analysis and Meta-Regression

 ,
,  and
and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
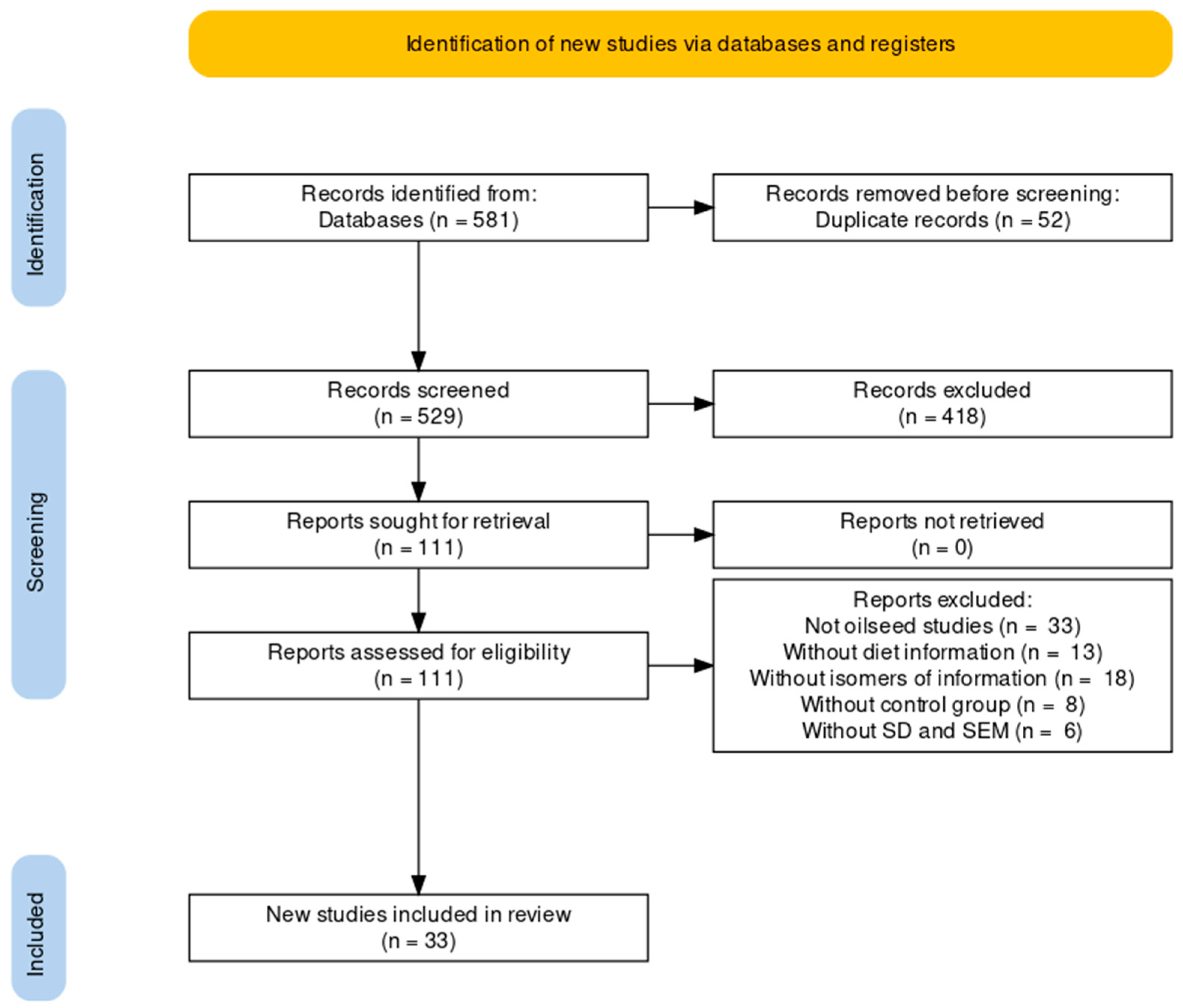
2.1. Search Strategy
2.2. Eligibility Criteria
2.3. Database
2.4. Meta-Analytical Procedure
3. Results
3.1. Milk Yield and Composition
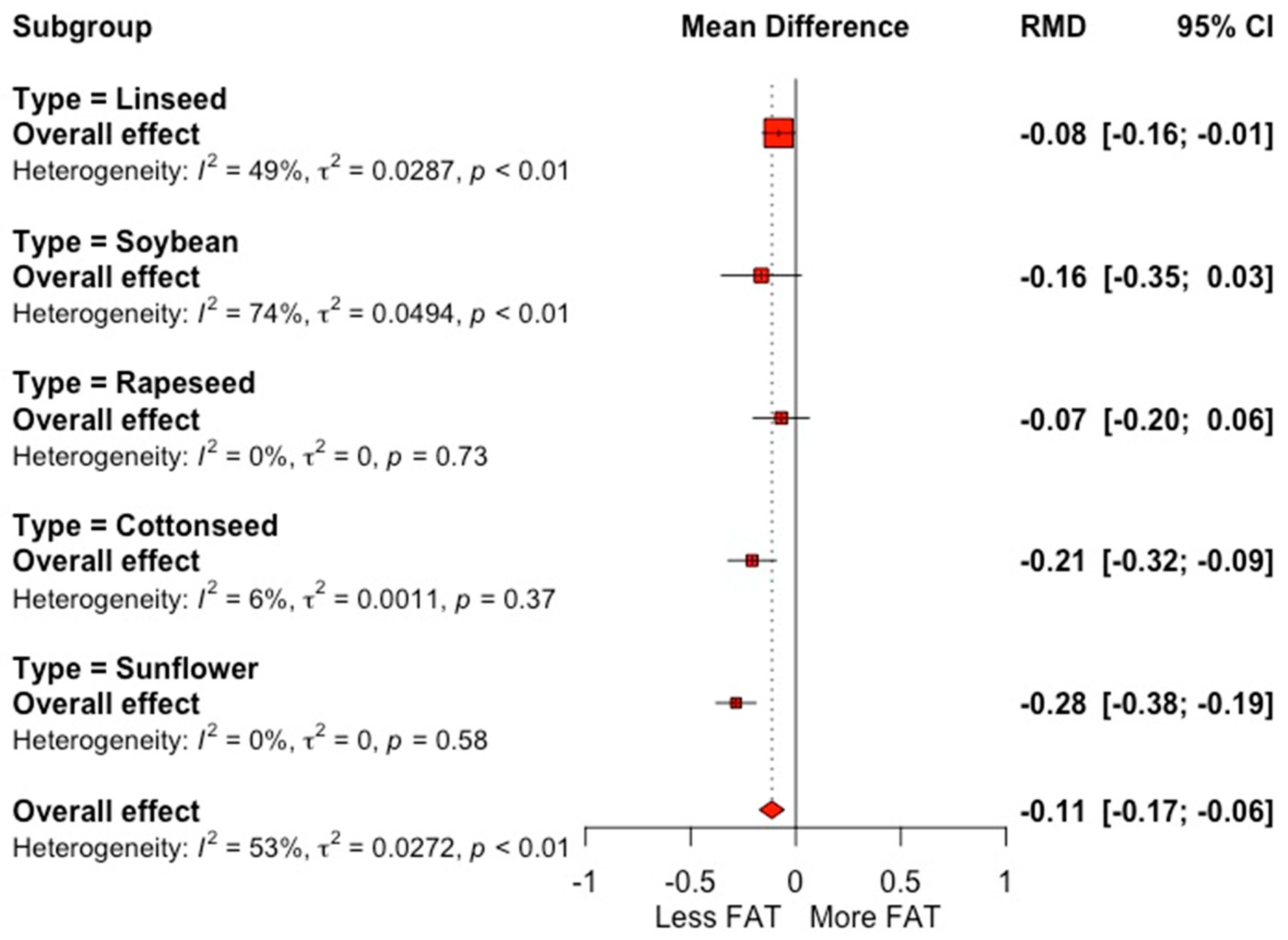
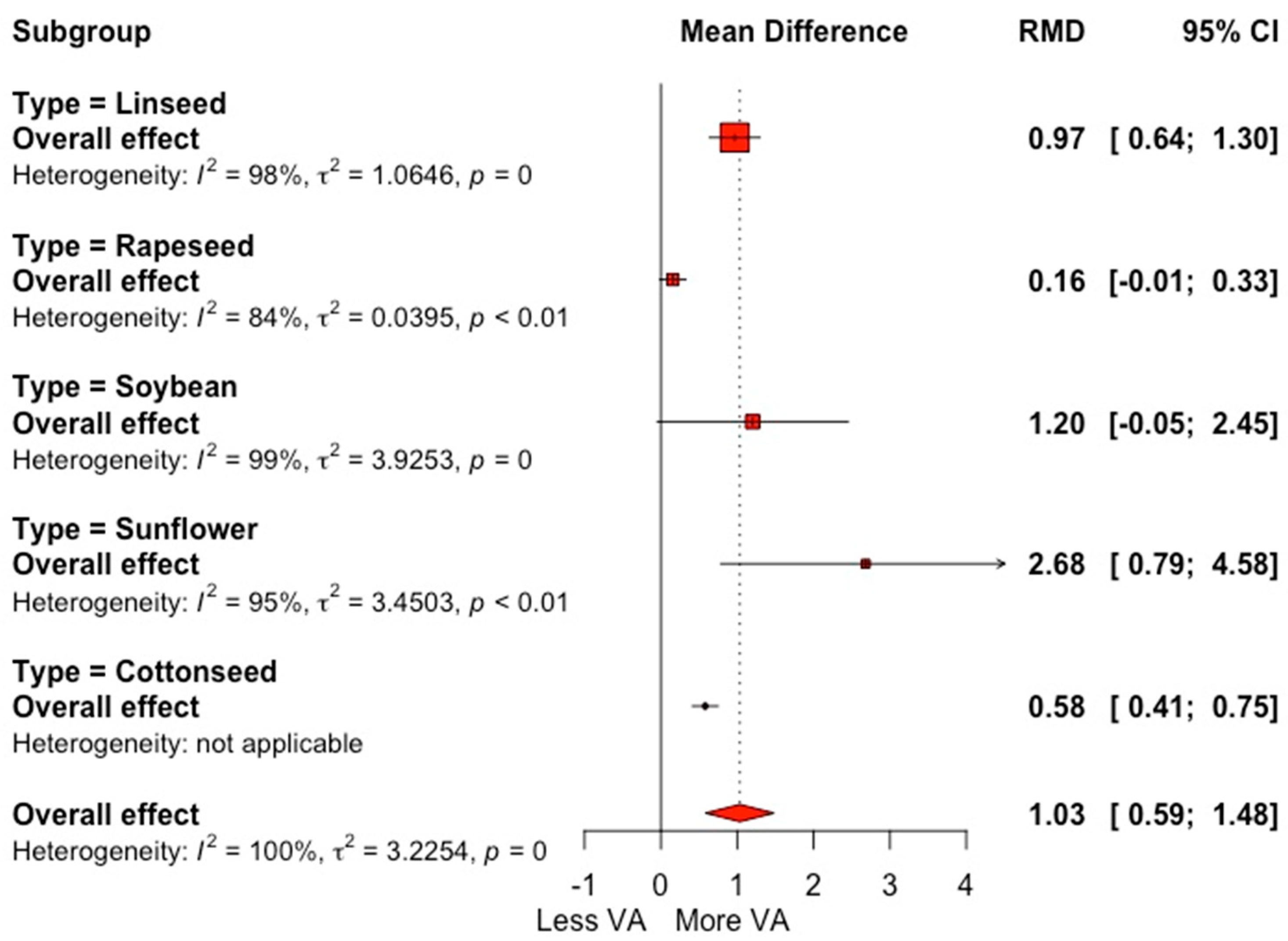
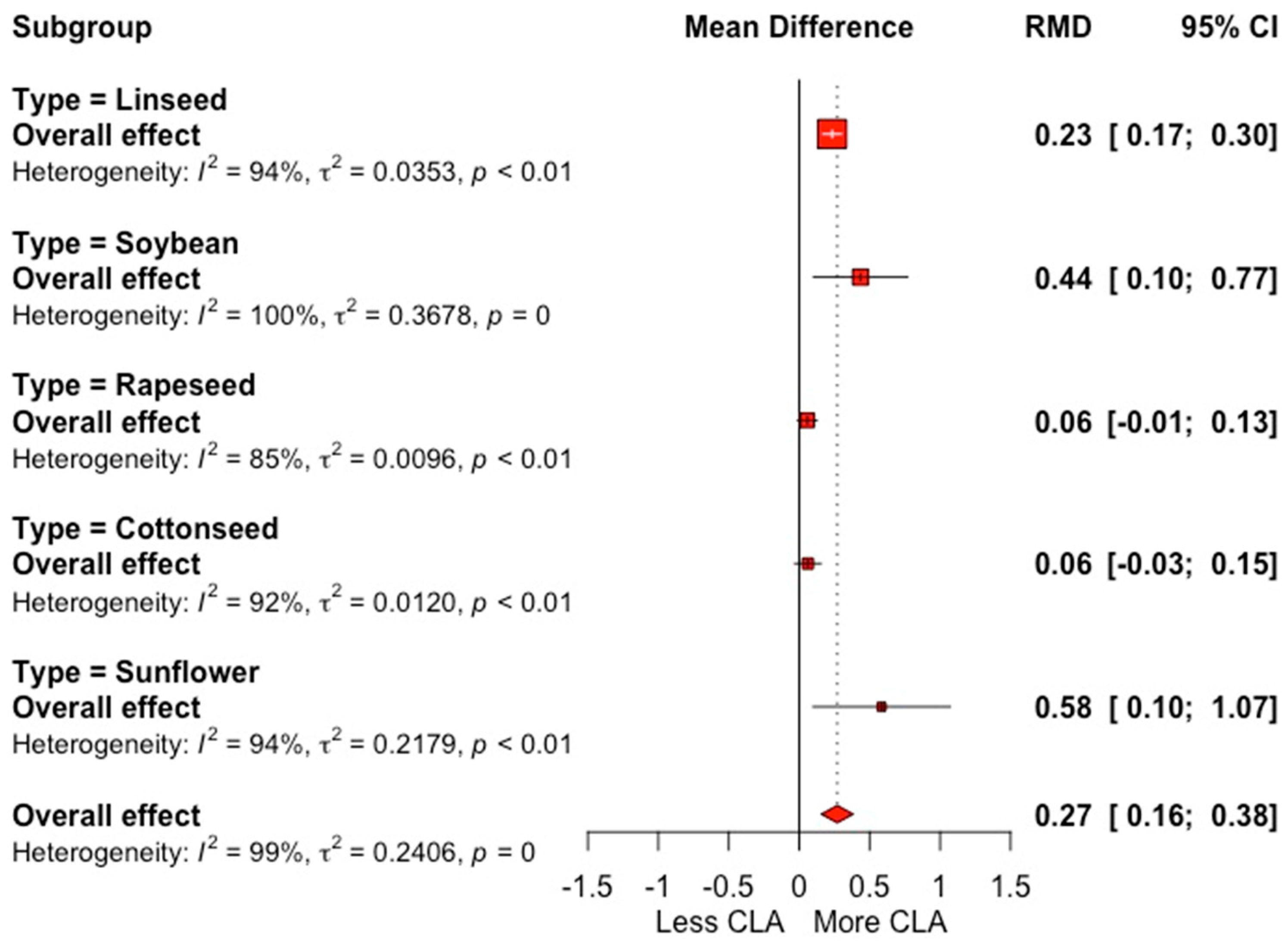
3.2. Fatty Acid Profile
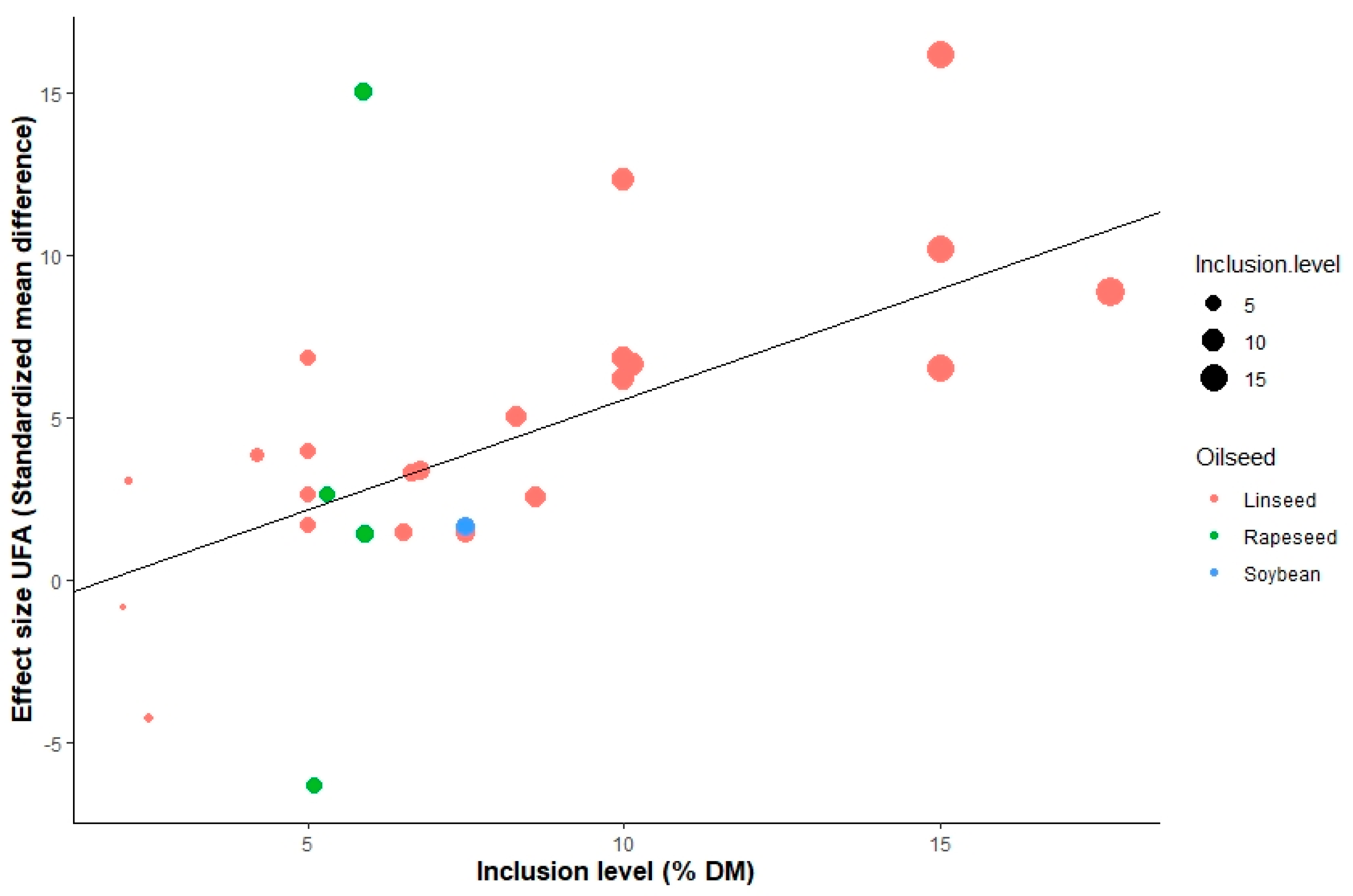
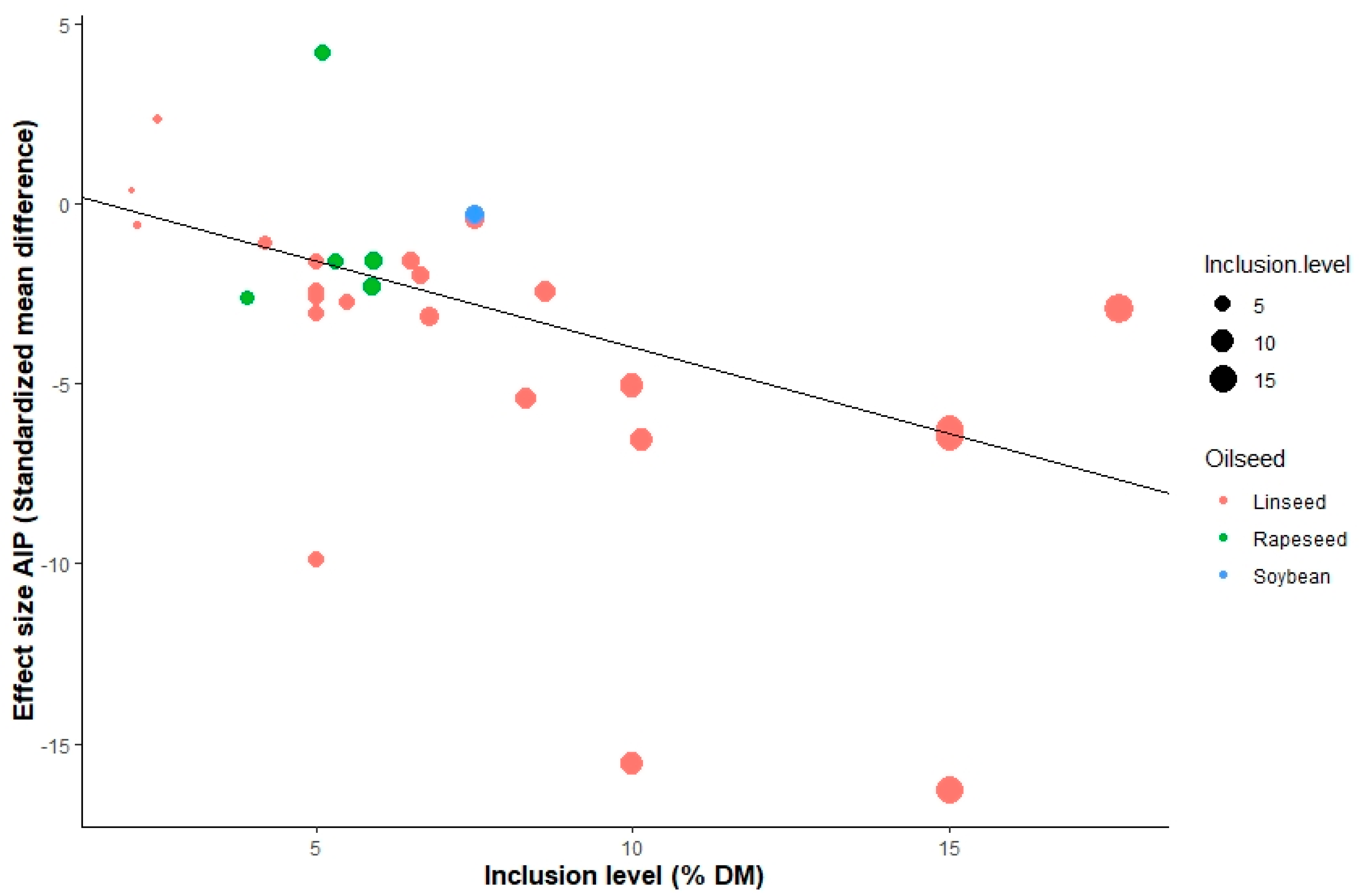
3.3. Meta-Regression
4. Discussion
4.1. Milk Yield and Composition
4.2. Milk Fatty Acid Profile
4.3. Meta-Regression
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steijns, J.M. Milk ingredients as nutraceuticals. Int. J. Dairy Technol. 2001, 54, 81–88. [Google Scholar] [CrossRef]
- O’Mahony, J.A.; Fox, P.F. Milk: An overview. Milk Proteins 2014, 19–73. [Google Scholar] [CrossRef]
- Collomb, M.; Schmid, A.; Sieber, R.; Wechsler, D.; Ryhänen, E.L. Conjugated linoleic acids in milk fat: Variation and physiological effects. Int. Dairy J. 2006, 16, 1347–1361. [Google Scholar] [CrossRef]
- Vargas-Bello-Pérez, E.; Garnsworthy, P.C. Trans fatty acids and their role in the milk of dairy cows. Int. J. Agric. Nat. Res. 2013, 40, 449–473. [Google Scholar] [CrossRef]
- Bionaz, M.; Vargas-Bello-Pérez, E.; Busato, S. Advances in fatty acids nutrition in dairy cows: From gut to cells and effects on performance. J. Anim. Sci. BioTechnol. 2020, 11, 110. [Google Scholar] [CrossRef] [PubMed]
- Hartigh, L.J. Conjugated linoleic acid effects on cancer, obesity, and atherosclerosis: A review of pre-clinical and human trials with current perspectives. Nutrients 2019, 11, 370. [Google Scholar] [CrossRef]
- Fritsche, J.; Rickert, R.; Steinhart, H.; Yurawecz, M.P. Formation, contents, and estimation of daily intake of conjugated linoleic acid isomers and trans-fatty acids in foods. In Advances in Conjugated Linoleic Acid Research; Yurawecz, M.P., Ed.; AOCS Press: Champaign, Il, USA, 1999; Volume 1, pp. 378–396. [Google Scholar]
- Lovegrove, J.A.; Givens, D.I. Dairy food products: Good or bad for cardiometabolic disease? Nutr. Res. Rev. 2016, 29, 249–267. [Google Scholar] [CrossRef]
- Koba, K.; Yanagita, T.K. Health benefits of conjugated linoleic acid (CLA). Obes. Res. Clin. Pract. 2014, 8, e525–e532. [Google Scholar] [CrossRef]
- Ip, C.; Banni, S.; Angioni, E.; Carta, G.; McGinley, J.; Thompson, H.J.; Barbano, D.; Bauman, D. Conjugated linoleic acid–enriched butter fat alters mammary gland morphogenesis and reduces cancer risk in rats. J. Nutr. 1999, 129, 2135–2142. [Google Scholar] [CrossRef]
- Parodi, P.W. Milk fat in human nutrition. Aust. J. Dairy Technol. 2004, 59, 3. [Google Scholar]
- McGuire, M.A.; McGuire, M.K. Conjugated linoleic acid (CLA): A ruminant fatty acid with beneficial effects on. J. Anim. Sci. 2000, 77, 1–8. [Google Scholar] [CrossRef]
- Bauman, D.E.; Harvatine, K.J.; Lock, A.L. Nutrigenomics, rumen-derived bioactive fatty acids, and the regulation of milk fat synthesis. Annu. Rev. Nutr. 2011, 31, 299–319. [Google Scholar] [CrossRef] [PubMed]
- Palmquist, D.L. Omega-3 fatty acids in metabolism, health, and nutrition and for modified animal product foods. Prof. Anim. Sci. 2009, 25, 207–249. [Google Scholar] [CrossRef]
- Angeles-Hernandez, J.C.; Miranda, M.; Muñoz-Benitez, A.L.; Vieyra-Alberto, R.; Morales-Aguilar, N.; Paz, E.A.; Gonzalez-Ronquillo, M. Zinc supplementation improves growth performance in small ruminants: A systematic review and meta-regression analysis. Anim. Prod. Sci. 2021, 61, 621–629. [Google Scholar] [CrossRef]
- Balduzzi, S.; Rücker, G.; Schwarzer, G. How to perform a meta-analysis with R: A practical tutorial. Evid. Based Ment. Health. 2019, 22, 153–160. [Google Scholar] [CrossRef] [PubMed]
- DerSimonian, R.; Laird, N. Meta-analysis in clinical trials revisited. Contemp. Clin. Trials 2015, 45, 139–145. [Google Scholar] [CrossRef]
- Hedges, L.V. Distribution theory for Glass’s estimator of effect size and related estimators. J. Educ. Behav. Stat. 1981, 6, 107–128. [Google Scholar] [CrossRef]
- Appuhamy, J.R.N.; Strathe, A.B.; Jayasundara, S.; Wagner-Riddle, C.; Dijkstra, J.; France, J.; Kebreab, E. Anti-methanogenic effects of monensin in dairy and beef cattle: A meta-analysis. J. Dairy Sci. 2013, 96, 5161–5173. [Google Scholar] [CrossRef]
- Higgins, J.P.; Thompson, S.G.; Deeks, J.J.; Altman, D.G. Measuring inconsistency in meta-analyses. BMJ 2003, 327, 557–560. [Google Scholar] [CrossRef]
- Serghiou, S.; Goodman, S.N. Random-effects meta-analysis: Summarizing evidence with caveats. JAMA 2019, 321, 301–302. [Google Scholar] [CrossRef]
- Viechtbauer, W. Conducting meta-analyses in R with the metafor package. J. Stat. Softw. 2010, 36, 1–48. [Google Scholar] [CrossRef]
- Light, R.J.; Pillemer, D.B. Summing Up: The Science of Reviewing Research; Harvard University Press: Cambridge, MA, USA, 1984; pp. 16–17. [Google Scholar]
- Akraim, F.; Nicot, M.C.; Juaneda, P.; Enjalbert, F. Conjugated linolenic acid (CLnA), conjugated linoleic acid (CLA) and other biohydrogenation intermediates in plasma and milk fat of cows fed raw or extruded linseed. Animal 2007, 1, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Caroprese, M.; Marzano, A.; Marino, R.; Gliatta, G.; Muschio, A.; Sevi, A. Flaxseed supplementation improves fatty acid profile of cow milk. J. Dairy Sci. 2010, 93, 2580–2588. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Ji, P.; Cao, Z.J.; Li, S.L. Effect of processing whole cottonseed on nutrient digestibility and ruminal fermentation in dairy cows. J. Anim. Feed Sci. 2007, 16, 537–542. [Google Scholar] [CrossRef]
- Chichlowski, M.W.; Schroeder, J.W.; Park, C.S.; Keller, W.L.; Schimek, D.E. Altering the fatty acids in milk fat by including canola seed in dairy cattle diets. J. Dairy Sci. 2005, 88, 3084–3094. [Google Scholar] [CrossRef]
- Chilliard, Y.; Martin, C.; Rouel, J.; Doreau, M. Milk fatty acids in dairy cows fed whole crude linseed, extruded linseed, or linseed oil, and their relationship with methane output. J. Dairy Sci. 2009, 92, 5199–5211. [Google Scholar] [CrossRef]
- Côrtes, C.; da Silva-Kazama, D.C.; Kazama, R.; Gagnon, N.; Benchaar, C.; Santos, G.T.D.; Zeoula, L.M.; Petit, H.V. Milk composition, milk fatty acid profile, digestion, and ruminal fermentation in dairy cows fed whole flaxseed and calcium salts of flaxseed oil. J. Dairy Sci. 2010, 93, 3146–3157. [Google Scholar] [CrossRef]
- Dhiman, T.R.; Helmink, E.D.; McMahon, D.J.; Fife, R.L.; Pariza, M.W. Conjugated linoleic acid content of milk and cheese from cows fed extruded oilseeds. J. Dairy Sci. 1999, 82, 412–419. [Google Scholar] [CrossRef]
- Egger, P.; Holzer, G.; Segato, S.; Werth, E.; Schwienbacher, F.; Peratoner, G.; Andrighetto, I.; Kasal, A. Effects of oilseed supplements on milk production and quality in dairy cows fed a hay-based diet. Ital. J. Anim. Sci. 2007, 6, 395–405. [Google Scholar] [CrossRef][Green Version]
- Ferlay, A.; Martin, B.; Lerch, S.; Gobert, M.; Pradel, P.; Chilliard, Y. Effects of supplementation of maize silage diets with extruded linseed, vitamin E and plant extracts rich in polyphenols, and morning v. evening milking on milk fatty acid profiles in Holstein and Montbéliarde cows. Animal 2010, 4, 627–640. [Google Scholar] [CrossRef]
- Ferlay, A.; Doreau, M.; Martin, C.; Chilliard, Y. Effects of incremental amounts of extruded linseed on the milk fatty acid composition of dairy cows receiving hay or corn silage. J. Dairy Sci. 2013, 96, 6577–6595. [Google Scholar] [CrossRef] [PubMed]
- Gandra, J.R.; Mingoti, R.D.; Barletta, R.V.; Takiya, C.S.; Verdurico, L.C.; Freitas, J.E.; Paiva, P.G.; Jesús, E.F.; Calomeni, G.D.; Rennó, F.P. Effects of flaxseed, raw soybeans and calcium salts of fatty acids on apparent total tract digestibility, energy balance and milk fatty acid profile of transition cows. Animal 2016, 10, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Sun, T.; Li, J. Effect of oilseeds rich in linoleic and linolenic acids on milk production and milk fatty acid composition in dairy cows. Front. Agric. China 2009, 3, 311–318. [Google Scholar] [CrossRef]
- Gonthier, C.; Mustafa, A.F.; Ouellet, D.R.; Chouinard, P.Y.; Berthiaume, R.; Petit, H.V. Feeding micronized and extruded flaxseed to dairy cows: Effects on blood parameters and milk fatty acid composition. J. Dairy Sci. 2005, 88, 748–756. [Google Scholar] [CrossRef]
- Kliem, K.E.; Humphries, D.J.; Reynolds, C.K.; Morgan, R.; Givens, D.I. Effect of oilseed type on milk fatty acid composition of individual cows, and also bulk tank milk fatty acid composition from commercial farms. Animal 2017, 11, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Kliem, K.E.; Humphries, D.J.; Kirton, P.; Givens, D.I.; Reynolds, C.K. Differential effects of oilseed supplements on methane production and milk fatty acid concentrations in dairy cows. Animal 2019, 13, 309–317. [Google Scholar] [CrossRef]
- Lee, H.G.; Hong, Z.S.; Wang, J.H.; Xu, C.X.; Jin, Y.C.; Kim, T.K.; Kim, Y.J.; Song, M.K.; Choi, Y.J. Milk conjugated linoleic acid (CLA) profile and metabolic responses of dairy cows fed with high-temperature-micro-time (HTMT) treated diets containing high quantity extruded soybean (ESB). Asian-Australas. J. Anim. Sci. 2009, 22, 1504–1512. [Google Scholar] [CrossRef]
- Lerch, S.; Pires, J.A.A.; Delavaud, C.; Shingfield, K.J.; Pomiès, D.; Martin, B.; Chilliard, Y.; Ferlay, A. Rapeseed or linseed in dairy cow diets over 2 consecutive lactations: Effects on adipose fatty acid profile and carry-over effects on milk fat composition in subsequent early lactation. J. Dairy Sci. 2015, 98, 1005–1018. [Google Scholar] [CrossRef]
- Liu, S.J.; Wang, J.Q.; Bu, D.P.; Wei, H.Y.; Zhou, L.Y.; Luo, Q.J. The effect of dietary vegetable oilseeds supplement on fatty acid profiles in milk fat from lactating dairy cows. Agric. Sci. China 2007, 6, 1002–1008. [Google Scholar] [CrossRef]
- Liu, Z.L.; Yang, D.P.; Chen, P.; Lin, S.B.; Jiang, X.Y.; Zhao, W.S.; Li, J.M.; Dong, W.X. Effect of dietary sources of roasted oilseeds on blood parameters and milk fatty acid composition. Czech. J. Anim. Sci. 2008, 53, 219. [Google Scholar] [CrossRef]
- Livingstone, K.M.; Humphries, D.J.; Kirton, P.; Kliem, K.E.; Givens, D.I.; Reynolds, C.K. Effects of forage type and extruded linseed supplementation on methane production and milk fatty acid composition of lactating dairy cows. J. Dairy Sci. 2015, 98, 4000–4011. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.F.; Chouinard, P.Y.; Christensen, D.A. Effects of feeding micronised flaxseed on yield and composition of milk from Holstein cows. J. Sci. Food Agric. 2003, 83, 920–926. [Google Scholar] [CrossRef]
- Neveu, C.; Baurhoo, B.; Mustafa, A. Effect of feeding extruded flaxseed with different forage: Concentrate ratios on the performance of dairy cows. J. Dairy Sci. 2013, 96, 3886–3894. [Google Scholar] [CrossRef]
- Neveu, C.; Baurhoo, B.; Mustafa, A. Effect of feeding extruded flaxseed with different grains on the performance of dairy cows and milk fatty acid profile. J. Dairy Sci. 2014, 97, 1543–1551. [Google Scholar] [CrossRef] [PubMed]
- Oeffner, S.P.; Qu, Y.; Just, J.; Quezada, N.; Ramsing, E.; Keller, M.; Cherian, G.; Goddick, L.; Bobe, G. Effect of flaxseed supplementation rate and processing on the production, fatty acid profile, and texture of milk, butter, and cheese. J. Dairy Sci. 2013, 96, 1177–1188. [Google Scholar] [CrossRef]
- Puppel, K.; Kuczyńska, B.; Nałęcz-Tarwacka, T.; Grodzki, H. Influence of linseed variety on fatty acid profile in cow’s milk. J. Sci. Food Agric. 2013, 93, 2276–2280. [Google Scholar] [CrossRef]
- Razzaghi, A.; Valizadeh, R.; Naserian, A.A.; Mesgaran, M.D.; Carpenter, A.J.; Ghaffari, M.H. Effect of dietary sugar concentration and sunflower seed supplementation on lactation performance, ruminal fermentation, milk fatty acid profile, and blood metabolites of dairy cows. J. Dairy Sci. 2016, 99, 3539–3548. [Google Scholar] [CrossRef]
- Resende, T.L.; Kraft, J.; Soder, K.J.; Pereira, A.B.D.; Woitschach, D.E.; Reis, R.B.; Brito, A.F. Incremental amounts of ground flaxseed decrease milk yield but increase n-3 fatty acids and conjugated linoleic acids in dairy cows fed high-forage diets. J. Dairy Sci. 2015, 98, 4785–4799. [Google Scholar] [CrossRef]
- Roca-Fernandez, A.I.; Gonzalez-Rodriguez, A.; Vazquez-Yanez, O.P.; Fernandez-Casado, J.A. Effects of concentrate source (cottonseed vs. barley) on milk performance and fatty acids profile of Holstein-Friesian dairy cows. Iran. J. Appl. Anim. Sci. 2011, 1, 245–252. [Google Scholar]
- Sarrazin, P.; Mustafa, A.F.; Chouinard, P.Y.; Raghavan, G.S.V.; Sotocinal, S.A. Performance of dairy cows fed roasted sunflower seed. J. Sci. Food Agric. 2004, 84, 1179–1185. [Google Scholar] [CrossRef]
- Stergiadis, S.; Leifert, C.; Seal, C.J.; Eyre, M.D.; Steinshamn, H.; Butler, G. Improving the fatty acid profile of winter milk from housed cows with contrasting feeding regimes by oilseed supplementation. Food Chem. 2014, 164, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.T.; Wittenberg, K.M.; Przybylski, R. Bovine milk fatty acid profiles produced by feeding diets containing solin, flax, and canola. J. Dairy Sci. 2002, 85, 1191–1196. [Google Scholar] [CrossRef]
- Whitlock, L.A.; Schingoethe, D.J.; Hippen, A.R.; Kalscheur, K.F.; Baer, R.J.; Ramaswamy, N.; Kasperson, K.M. Fish oil and extruded soybeans fed in combination increase conjugated linoleic acids in milk of dairy cows more than when fed separately. J. Dairy Sci. 2002, 85, 234–243. [Google Scholar] [CrossRef]
- Ye, J.A.; Wang, C.; Wang, H.F.; Ye, H.W.; Wang, B.X.; Liu, H.Y.; Wang, Y.M.; Yang, Z.Q.; Liu, J.X. Milk production and fatty acid profile of dairy cows supplemented with flaxseed oil, soybean oil, or extruded soybeans. Acta Agric. Scand A Anim. Sci. 2009, 59, 121–129. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Dairy Cattle, 7th ed.; National Academic Press: Washington, DC, USA, 2001; p. 292. [Google Scholar]
- Vargas-Bello-Pérez, E.; Robles-Jimenez, L.E.; Ayala-Hernández, R.; Romero-Bernal, J.; Pescador-Salas, N.; Castelán-Ortega, O.A.; González-Ronquillo, M. Effects of calcium soaps from palm, canola and safflower oils on dry matter intake, nutrient digestibility, milk production, and milk composition in dairy goats. Animals 2020, 10, 1728. [Google Scholar] [CrossRef] [PubMed]
- Rabiee, A.R.; Breinhild, K.; Scott, W.; Golder, H.M.; Block, E.; Lean, I.J. Effect of fat additions to diets of dairy cattle on milk production and components: A meta-analysis and meta-regression. J. Dairy Sci. 2012, 95, 3225–3247. [Google Scholar] [CrossRef]
- Palmquist, D.L.; Jenkins, T.C. A 100-Year Review: Fat feeding of dairy cows. J. Dairy Sci. 2017, 100, 10061–10077. [Google Scholar] [CrossRef] [PubMed]
- Glasser, F.; Ferlay, A.; Chilliard, Y. Oilseed lipid supplements and fatty acid composition of cow milk: A meta-analysis. J. Dairy Sci. 2008, 91, 4687–4703. [Google Scholar] [CrossRef]
- Mahdavi, A.; Mahdavi, A.; Darabighane, B.; Mead, A.; Lee, M.R. Effects of soybean oil supplement to diets of lactating dairy cows, on productive performance, and milk fat acids profile: A meta-analysis. Ital. J. Anim. Sci. 2009, 18, 809–819. [Google Scholar] [CrossRef]
- Leduc, M.; Létourneau-Montminy, M.P.; Gervais, R.; Chouinard, P.Y. Effect of dietary flax seed and oil on milk yield, gross composition, and fatty acid profile in dairy cows: A meta-analysis and meta-regression. J. Dairy Sci. 2017, 100, 8906–8927. [Google Scholar] [CrossRef]
- Bauman, D.E.; Griinari, J.M. Nutritional regulation of milk fat synthesis. Annu. Rev. Nutr. 2003, 23, 203–227. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, T.C.; McGuire, M.A. Major advances in nutrition: Impact on milk composition. J. Dairy Sci. 2006, 89, 1302–1310. [Google Scholar] [CrossRef]
- Petit, H.V. Digestion, milk production, milk composition, and blood composition of dairy cows fed formaldehyde treated flaxseed or sunflower seed. J. Dairy Sci. 2003, 86, 2637–2646. [Google Scholar] [CrossRef]
- Wilde, P.F.; Dawson, R.M.C. The biohydrogenation of α-linoleic acid and oleic acid by rumen micro-organisms. Biochem. J. 1966, 98, 469. [Google Scholar] [CrossRef] [PubMed]
- Sterk, A.; Van Vuuren, A.M.; Hendriks, W.H.; Dijkstra, J. Effects of different fat sources, technological forms and characteristics of the basal diet on milk fatty acid profile in lactating dairy cows—A meta-analysis. J. Agric. Sci. 2012, 150, 495–517. [Google Scholar] [CrossRef]
- Khiaosa-ard, R.; Kreuzer, M.; Leiber, F. Apparent recovery of C18 polyunsaturated fatty acids from feed in cow milk: A meta-analysis of the importance of dietary fatty acids and feeding regimens in diets without fat supplementation. J. Dairy Sci. 2015, 98, 6399–6414. [Google Scholar] [CrossRef]
- Sun, X.Q.; Gibbs, S.J. Diurnal variation in fatty acid profiles in rumen digesta from dairy cows grazing high-quality pasture. Anim. Feed Sci. Technol. 2012, 177, 152–160. [Google Scholar] [CrossRef]
- Turpeinen, A.M.; Mutanen, M.; Aro, A.; Salminen, I.; Basu, S.; Palmquist, D.L.; Griinari, J.M. Bioconversion of vaccenic acid to conjugated linoleic acid in humans. Am. J. Clin. Nutr. 2002, 76, 504–510. [Google Scholar] [CrossRef]
- Nguyen, Q.V.; Malau-Aduli, B.S.; Cavalieri, J.; Nichols, P.D.; Malau-Aduli, A.E. Enhancing omega-3 long-chain polyunsaturated fatty acid content of dairy-derived foods for human consumption. Nutrients 2019, 11, 743. [Google Scholar] [CrossRef]
- Wang, H.; Fox, C.S.; Troy, L.M.; Mckeown, N.M.; Jacques, P.F. Longitudinal association of dairy consumption with the changes in blood pressure and the risk of incident hypertension: The Framingham Heart Study. Br. J. Nutr. 2015, 114, 1887–1899. [Google Scholar] [CrossRef]
- Gholami, F.; Khoramdad, M.; Esmailnasab, N.; Moradi, G.; Nouri, B.; Safiri, S.; Alimohamadi, Y. The effect of dairy consumption on the prevention of cardiovascular diseases: A meta-analysis of prospective studies. J. Cardiovasc. Thorac. Res. 2017, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Eussen, S.J.; van Dongen, M.C.; Wijckmans, N.; den Biggelaar, L.; Elferink, S.J.O.; Singh-Povel, C.M.; Schram, M.T.; Sep, S.J.S.; van del Kallen, C.J.; Koster, A.; et al. Consumption of dairy foods in relation to impaired glucose metabolism and type 2 diabetes mellitus: The Maastricht Study. Br. J. Nutr. 2016, 115, 1453–1461. [Google Scholar] [CrossRef] [PubMed]
- Shahmirzadi, F.E.; Ghavamzadeh, S.; Zamani, T. The effect of conjugated linoleic acid supplementation on body composition, serum insulin and leptin in obese adults. Arch. Iran. Med. 2019, 22, 255. [Google Scholar]
- Kim, J.H.; Kim, Y.; Kim, Y.J.; Park, Y. Conjugated linoleic acid: Potential health benefits as a functional food ingredient. Annu. Rev. Food Sci. Technol. 2016, 7, 221–244. [Google Scholar] [CrossRef]
- Chilliard, Y.; Ferlay, A. Dietary lipids and forages interactions on cow and goat milk fatty acid composition and sensory properties. Reprod. Nutr. Dev. 2004, 44, 467–492. [Google Scholar] [CrossRef]
- Philippeau, C.; Lettat, A.; Martin, C.; Silberberg, M.; Morgavi, D.P.; Ferlay, A.; Berger, C.; Nozière, P. Effects of bacterial direct-fed microbials on ruminal characteristics, methane emission, and milk fatty acid composition in cows fed high-or low-starch diets. J. Dairy Sci. 2017, 100, 2637–2650. [Google Scholar] [CrossRef]
- Kholif, A.E.; Morsy, T.A.; Abdo, M.M. Crushed flaxseed versus flaxseed oil in the diets of Nubian goats: Effect on feed intake, digestion, ruminal fermentation, blood chemistry, milk production, milk composition and milk fatty acid profile. Anim. Feed Sci. Technol. 2018, 244, 66–75. [Google Scholar] [CrossRef]
- Pi, Y.; Ma, L.; Pierce, K.M.; Wang, H.R.; Xu, J.C.; Bu, D.P. Rubber seed oil and flaxseed oil supplementation alter digestion, ruminal fermentation and rumen fatty acid profile of dairy cows. Animal 2019, 13, 2811–2820. [Google Scholar] [CrossRef]
- Martínez, M.A.L.; Pérez, H.M.; Pérez, A.L.M.; Carrión, P.D.; Gómez, C.G.; Garzón, S.A.I. Efecto de los aceites y semillas en dietas para rumiantes sobre el perfil de ácidos grasos de la leche. Revisión. Rev. Mex. Cienc. Pecu. 2013, 4, 319–338. [Google Scholar]
- Maki, K.C.; Eren, F.; Cassens, M.E.; Dicklin, M.R.; Davidson, M.H. ω-6 polyunsaturated fatty acids and cardiometabolic health: Current evidence, controversies, and research gaps. Adv. Nutr. 2018, 9, 688–700. [Google Scholar] [CrossRef]
- Livingstone, K.M.; Lovegrove, J.A.; Cockcroft, J.R.; Elwood, P.C.; Pickering, J.E.; Givens, D.I. Does dairy food intake predict arterial stiffness and blood pressure in men? Evidence from the Caerphilly Prospective Study. Hypertension 2013, 61, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Plata-Reyes, D.A.; Hernández-Mendo, O.; Vieyra-Alberto, R.; Albarrán-Portillo, B.; Martínez-García, C.G.; Arriaga-Jordán, C.M. Kikuyu grass in winter–spring time in small-scale dairy systems in the highlands of central Mexico in terms of cow performance and fatty acid profile of milk. Trop. Anim. Health Prod. 2021, 53, 225. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, J.C.; Ramírez, C.H.; Lutz, G. Caracterización y determinación del potencial Aterogénico de quesos producidos en Costa Rica. Rev. Cien. Tec. 2006, 24. [Google Scholar]
- Soyeurt, H.; Dardenne, P.; Gillon, A.; Croquet, C.; Vanderick, S.; Mayeres, P.; Bertozzi, C.; Gengler, N. Variation in fatty acid contents of milk and milk fat within and across breeds. J. Dairy Sci. 2006, 89, 4858–4865. [Google Scholar] [CrossRef]
- Carroll, S.M.; DePeters, E.J.; Taylor, S.J.; Rosenberg, H.; Pere-Monti, H.; Capps, V.A. Milk composition of Holstein, Jersey, and Brown Swiss cows in response to increasing levels of dietary fat. Anim. Feed Sci. Technol. 2006, 131, 451–473. [Google Scholar] [CrossRef]
- Wiking, L.; Björck, L.; Nielsen, J.H. Influence of feed composition on stability of fat globules during pumping of raw milk. Int. Dairy J. 2003, 13, 797–803. [Google Scholar] [CrossRef]
- Wiking, L.; Stagsted, J.; Björck, L.; Nielsen, J.H. Milk fat globule size is affected by fat production in dairy cows. Int. Dairy J. 2004, 14, 909–913. [Google Scholar] [CrossRef]
- Stoop, W.M.; Bovenhuis, H.; Heck, J.M.L.; Van Arendonk, J.A.M. Effect of lactation stage and energy status on milk fat composition of holstein-friesian cows. J. Dairy Sci. 2009, 92, 1469–1478. [Google Scholar] [CrossRef]
- Garnsworthy, P.C.; Masson, L.L.; Lock, A.L.; Mottram, T.T. Variation of milk citrate with stage of lactation and de novo fatty acid synthesis in dairy cows. J. Dairy Sci. 2006, 89, 1604–1612. [Google Scholar] [CrossRef]
- Gross, J.J.; Bruckmaier, R.M. Metabolic challenges in lactating dairy cows and their assessment via established and novel indicators in milk. Animal 2019, 13 (Suppl. 1), s75–s81. [Google Scholar] [CrossRef] [PubMed]
- Ueda, K.; Ferlay, A.; Chabrot, J.; Loor, J.J.; Chilliard, Y. Effect of linseed oil supplementation on ruminal digestion in dairy cows fed diets with different forage: Concentrate ratios. J. Dairy Sci. 2003, 86, 3999–4007. [Google Scholar] [CrossRef]
- Abuelfatah, K.; Zakaria, M.Z.A.B.; Meng, G.Y.; Sazili, A.Q. Changes in fatty acid composition and distribution of N-3 fatty acids in goat tissues fed different levels of whole linseed. Sci. World J. 2014, 2014, 934154. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Bu, D.P.; Wang, J.Q.; Liu, L.; Liang, S.; Wei, H.Y.; Zhou, L.Y.; Li, D.; Loor, J.J. Effect of incremental levels of fish oil supplementation on specific bacterial populations in bovine ruminal fluid. J. Anim. Physiol. Anim. Nutr. 2012, 96, 9–16. [Google Scholar] [CrossRef]
- Palmquist, D.L.; Lock, A.L.; Shingfield, K.J.; Bauman, D.E. Biosynthesis of conjugated linoleic acid in ruminants and humans. Adv. Food Nutr. Res. 2005, 50, 179–217. [Google Scholar]
- Collomb, M.; Sollberger, H.; Bütikofer, U.; Sieber, R.; Stoll, W.; Schaeren, W. Impact of a basal diet of hay and fodder beet supplemented with rapeseed, linseed and sunflowerseed on the fatty acid composition of milk fat. Int. Dairy J. 2004, 14, 549–559. [Google Scholar] [CrossRef]
- Vieyra-Alberto, R.; Arriaga-Jordán, C.M.; Domínguez-Vara, I.A.; Bórquez-Gastelum, J.L.; Morales-Almaráz, E. Efecto del aceite de soya sobre la concentración de los ácidos grasos vaccenico y ruménico en leche de vacas en pastoreo. Agrociencia 2017, 51, 299–313. [Google Scholar]
- Vieyra-Alberto, R.; Zetina-Martínez, R.E.; Olivares-Pérez, J.; Galicia-Aguilar, H.H.; Rojas-Hernández, S.; Ángeles-Hernández, J.C. Effect of soybean grain (Glycine max L.) supplementation on the production and fatty acid profile in milk of grazing cows in the dry tropics of Mexico. Trop. Anim. Health Prod. 2022, 54, 52. [Google Scholar] [CrossRef]
- Dhiman, T.R.; Zanten, K.V.; Satter, L.D. Effect of dietary fat source on fatty acid composition of cow’s milk. J. Sci. Food Agric. 1995, 69, 101–107. [Google Scholar] [CrossRef]
- Rafiee-Yarandi, H.; Ghorbani, G.R.; Alikhani, M.; Sadeghi-Sefidmazgi, A.; Drackley, J.K. A comparison of the effect of soybeans roasted at different temperatures versus calcium salts of fatty acids on performance and milk fatty acid composition of mid-lactation Holstein cows. J. Dairy Sci. 2016, 99, 5422–5435. [Google Scholar] [CrossRef]
- Manso, T.; Gallardo, B.; Guerra-Rivas, C. Modifying milk and meat fat quality through feed changes. Small Rumin. Res. 2016, 142, 31–37. [Google Scholar] [CrossRef]
- Angeles-Hernandez, J.C.; Vieyra-Alberto, R.; Kebreab, E.; Appuhamy, J.A.R.N.; Dougherty, H.C.; Castelan-Ortega, O.; Gonzalez-Ronquillo, M. Effect of forage to concentrate ratio and fat supplementation on milk composition in dairy sheep: A meta-analysis. Livest. Sci. 2020, 238, 104069. [Google Scholar] [CrossRef]
- Loor, J.J.; Ferlay, A.; Ollier, A.; Doreau, M.; Chilliard, Y. Relationship among trans and conjugated fatty acids and bovine milk fat yield due to dietary concentrate and linseed oil. J. Dairy Sci. 2005, 88, 726–740. [Google Scholar] [CrossRef]
- Castro, T.; Martinez, D.; Isabel, B.; Cabezas, A.; Jimeno, V. Vegetable oils rich in polyunsaturated fatty acids supplementation of dairy cows’ diets: Effects on productive and reproductive performance. Animals 2019, 9, 205. [Google Scholar] [CrossRef] [PubMed]
- Federer, W.T. On Planning Repeated Measure Experiments; Cornell University: New York, NY, USA, 1986; pp. 1–7. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Breed 1 | Lactation Stage | Type 2 | Way 3 | Inclusion (%) | Washout (Days) | Design 4 |
|---|---|---|---|---|---|---|---|
| [24] | H | 2nd | Lin | Wh, Ex | 16.4, 16.7 | 19 | Cross |
| [25] | H | 1st | Lin | Wh | 6.5 | 14 | Long |
| [26] | H | 1st | Sb, Rap, Cott | Wh, Ex, Gr, | 5, 10.1, 10.5 | 14, 18 | Cross, Long |
| [27] | H | 1st | Rap | Gr | 14 | 14 | Long |
| [28] | H | 3rd | Lin | Wh | 12.4 | 23 | Cross |
| [29] | H | 1st | Lin | Wh | 4.2 | 21 | Cross |
| [30] | H | 2nd | Sb, Cott | Ex | 11.9 | 21 | Long |
| [31] | H | 2nd | Rap, Lin | Ex, Gr | 5, 6.1 | 21 | Cross |
| [32] | H | 1st | Lin | Ex | 17.7 | 21 | Long |
| [33] | H | 1st, 2nd | Lin | Ex | 5, 10, 15 | 21 | Cross |
| [34] | H | 1st | Lin, Sb | Wh | 8, 16 | 84 | Long |
| [35] | H | 1st | Lin, Sb | Rs | 7.5 | 10 | Cross |
| [36] | H | 3rd | Lin | Wh, Ex, Gr | 12.1, 12.7 | 21 | Cross |
| [37] | H | 3rd | Lin, Rap | Ex, Gr | 3.9, 5.5 | 21 | Cross |
| [38] | H | 2nd | Lin, Rap | Ex, Gr | 5.9, 8.6 | 21 | Cross |
| [39] | H | 2nd | Sb | Ex | 7.5, 5.0 | 7 | Long |
| [40] | H | 1st | Lin, Rap | Wh, Ex | 2.5, 5.1 | 120 | Long |
| [41] | H | 2nd | Sb | Wh, Ex | 8.5 | 21 | Long |
| [42] | H | 1st | Sb, Lin, Sun, Cott | Rs | 7.5 | 14 | Long |
| [43] | H | 1st | Lin | Ex | 5 | - | Cross |
| [44] | H | 1st | Lin | Wh, Gr | 7 | 14 | Cross |
| [45] | H | 1st | Lin | Ex | 6.5 | 14 | Cross |
| [46] | H | 1st | Lin | Ex | 10 | 14 | Cross |
| [47] | H | 2nd | Lin | Ex, Gr | 4.3, 8.1, 8.3, 11.8 | 14 | Cross |
| [48] | H | 2nd | Lin | Wh | 2.1, 2.2 | 7 | Long |
| [49] | H | 1st | Sun | Gr | 8 | 14 | Cross |
| [50] | JR | 2nd | Lin | Gr | 5, 10, 15 | 14 | Cross |
| [51] | H | 3rd | Cott | Wh | 12 | - | Long |
| [52] | H | 1st | Sun | Wh, Rs | 7.8 | 14 | Cross |
| [53] | H | - | Lin, Rap | Wh | 6.6, 6.8, 8.3, 10.5 | - | Long |
| [54] | H | - | Lin, Rap | Wh | 8.3 | 16 | Cross |
| [55] | H, BS | 1st | Sb | Ex | 10.6 | 14 | Cross |
| [56] | H | 2nd | Sb | Ex | 10.7 | 7 | Long |
| Item | n 1 | Control Means (SD) | Effect Size | Heterogeneity | ||||
|---|---|---|---|---|---|---|---|---|
| RMD 2 | p-Value | SMD 3 | p-Value | I2 RM 4 | I2 MM 5 | |||
| Milk yield (kg day−1) | 73 | 28.02 (6.59) | −0.25 | 0.200 | −0.06 | 0.330 | 50.2 | 0.0 |
| Fat content (g 100 g−1) | 70 | 3.86 (0.46) | −0.11 | 0.001 | −0.21 | 0.002 | 53.0 | 25.8 |
| Protein content (g 100 g−1) | 69 | 3.18 (0.2) | −0.04 | 0.007 | −0.20 | 0.003 | 65.4 | 33.1 |
| Lactose content (g 100 g−1) | 53 | 4.77 (0.19) | 0.01 | 0.280 | 0.07 | 0.270 | 43.5 | 19.0 |
| Oleic acid (g 100 g−1 FA) | 69 | 20.87 (5.04) | 3.44 | <0.001 | 1.50 | <0.001 | 92.4 | 82.3 |
| Linoleic acid (g 100 g−1 FA) | 71 | 2.12 (0.51) | 0.09 | 0.007 | 0.16 | 0.250 | 86.6 | 83.3 |
| Linolenic acid (g 100 g−1 FA) | 70 | 0.49 (0.25) | 0.28 | <0.001 | 1.68 | <0.001 | 97.8 | 76.7 |
| Vaccenic acid (g 100 g−1 FA) | 70 | 1.61 (0.88) | 1.03 | <0.001 | 1.33 | <0.001 | 99.7 | 95.2 |
| CLA (g 100 g−1 FA) | 77 | 0.54 (0.24) | 0.27 | <0.001 | 1.28 | <0.001 | 99.4 | 78.5 |
| Σ Omega−3 PUFA (g 100 g−1 FA) | 33 | 0.74(0.27) | 0.46 | <0.001 | 2.23 | 0.001 | 97.1 | 53.0 |
| Σ Omega−6 PUFA (g 100 g−1 FA) | 33 | 2.12(0.54) | −0.01 | 0.780 | −0.09 | 0.530 | 81.0 | 48.0 |
| SFA (g 100 g−1 FA) | 30 | 63.54(17.03) | −6.51 | <0.001 | −1.73 | <0.001 | 83.1 | 7.6 |
| UFA (g 100 g−1 FA) | 28 | 28.72(5.95) | 8.32 | <0.001 | 3.79 | <0.001 | 93.5 | 61.7 |
| Atherogenic index (AI) | 30 | 3.01(0.88) | −1.01 | <0.001 | −2.47 | <0.001 | 91.5 | 73.9 |
| Item | Meta-Regression Parameters (β) | Best Model | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Animal | Diet and Oilseeds | Experimental | <I2 | R2 | |||||||||
| Breed | Stage | Type 1 | Way 2 | Intake | Linoleic | Linolenic | Forage | NDF | Washout | Design | |||
| Milk yield (kg day−1) | Rs −0.47 *; Wh −0.28 * | −0.05 *** | 100.0 | 100 | |||||||||
| Fat content (g 100 g−1) | JR 0.38 * | 2nd 0.39 * | Sun −0.46 * | Wh 0.21 * | 0.01 * | −0.07 * | 51.3 | 57.8 | |||||
| Protein content (g 100 g−1) | 1st 1.57 **; 3rd 1.57 * | 0.02 * | 49.3 | 53.1 | |||||||||
| Lactose content (g 100 g−1) | Lin −0.09 *; Rap −0.10 * | −0.005 *** | 56.3 | 74.7 | |||||||||
| Linoleic acid (g 100 g−1 FA) | JR −0.61 * | 1st 0.49 * | Rap −0.96 * | Gr 0.43 * | −0.02 * | 3.8 | 11.8 | ||||||
| Linolenic acid (g 100 g−1 FA) | 1st −2.4 *; 2nd −2.33 * 3rd −3.79 ** | Rs 4.16 ** | −0.12 * | −0.13 *** | −0.02 * | 21.5 | 33.5 | ||||||
| Oleic acid (g 100 g−1 FA) | 3rd −4.07 * | Rap 6.71 *: Sb 3.54*; Sun 4.92 ** | Gr 2.63 *; Rs 4.41 **; Wh 2.61 * | 0.33 * | −0.23 *** | −0.15 *** | −0.07 * | −0.05 ** | 10.9 | 9.8 | |||
| Vaccenic acid (g 100 g−1 FA) | Lin −3.79 ***; Rap −3.92 ***; Sb −3.11 *** | −0.04 * | −0.02 *** | Cross −1.31 * | 4.6 | 95.2 | |||||||
| CLA (g 100 g−1 FA) | Lin 1.29 *; Sb 1.65 *; Sun 1.96 * | Wh −0.18 ** | −0.09 * | −0.03 ** | Long 1.45 *** | 21.0 | 21.1 | ||||||
| Σ Omega-3 PUFA (g 100 g−1 FA) | JR 1.15 * | Rap 1.35 * | Gr −1.38 *; Rs −0.74 * | 0.04 * | 45.4 | 50.7 | |||||||
| Σ Omega-6 PUFA (g 100 g−1 FA) | Sb 0.67 * | Rs 0.49 ** | −0.01 * | −0.11 * | 40.7 | 75.1 | |||||||
| SFA (g 100 g−1 FA) | Wh 4.25 *** | −0.37 ** | 3.26 * | Long 4.85 ** | 90.7 | 96.7 | |||||||
| UFA (g 100 g−1 FA) | 1st −7.31 ** | Rap 12.25 ** | Rs −5.43 *; Wh −10.98 *; Gr −14.66 ** | 0.79 * | −0.08 * | 0.19 *** | −0.15 *** | 34.0 | 98.4 | ||||
| Atherogenic index (AI) | 1st −0.89 *; 2nd 1.15 * | Rs 1.81 ***; Wh 1.23 *** | −0.12 * | 0.01 ** | 19.2 | 93.9 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plata-Pérez, G.; Angeles-Hernandez, J.C.; Morales-Almaráz, E.; Del Razo-Rodríguez, O.E.; López-González, F.; Peláez-Acero, A.; Campos-Montiel, R.G.; Vargas-Bello-Pérez, E.; Vieyra-Alberto, R. Oilseed Supplementation Improves Milk Composition and Fatty Acid Profile of Cow Milk: A Meta-Analysis and Meta-Regression. Animals 2022, 12, 1642. https://doi.org/10.3390/ani12131642
Plata-Pérez G, Angeles-Hernandez JC, Morales-Almaráz E, Del Razo-Rodríguez OE, López-González F, Peláez-Acero A, Campos-Montiel RG, Vargas-Bello-Pérez E, Vieyra-Alberto R. Oilseed Supplementation Improves Milk Composition and Fatty Acid Profile of Cow Milk: A Meta-Analysis and Meta-Regression. Animals. 2022; 12(13):1642. https://doi.org/10.3390/ani12131642
Chicago/Turabian StylePlata-Pérez, Genaro, Juan C. Angeles-Hernandez, Ernesto Morales-Almaráz, Oscar E. Del Razo-Rodríguez, Felipe López-González, Armando Peláez-Acero, Rafael G. Campos-Montiel, Einar Vargas-Bello-Pérez, and Rodolfo Vieyra-Alberto. 2022. "Oilseed Supplementation Improves Milk Composition and Fatty Acid Profile of Cow Milk: A Meta-Analysis and Meta-Regression" Animals 12, no. 13: 1642. https://doi.org/10.3390/ani12131642
APA StylePlata-Pérez, G., Angeles-Hernandez, J. C., Morales-Almaráz, E., Del Razo-Rodríguez, O. E., López-González, F., Peláez-Acero, A., Campos-Montiel, R. G., Vargas-Bello-Pérez, E., & Vieyra-Alberto, R. (2022). Oilseed Supplementation Improves Milk Composition and Fatty Acid Profile of Cow Milk: A Meta-Analysis and Meta-Regression. Animals, 12(13), 1642. https://doi.org/10.3390/ani12131642