
Current State of In Vitro Embryo Production in African Lion (Panthera leo)

 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Animals
2.3. Transport of Ovaries until Processing
2.4. Oocyte Retrieval and In Vitro Maturation (IVM)
2.5. Fertilization of Oocytes
2.6. Embryo Culture
2.7. Vitrification
2.8. Statistical Analysis
3. Results
3.1. Impact of Age of Animals on Oocyte Numbers and IVM/IVF Potential
3.2. Method Changes over Time
3.2.1. Method Variations without Impact on IVM/IVF Outcome
3.2.2. Method Variations with Impact on IVM/IVF Outcome
3.3. Duration of Maturation
4. Discussion
5. Summary/Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Herrick, J.R.; Bond, J.B.; Magarey, G.M.; Bateman, H.L.; Krisher, R.L.; Dunford, S.A.; Swanson, W.F. Toward a Feline-Optimized Culture Medium: Impact of Ions, Carbohydrates, Essential Amino Acids, Vitamins, and Serum on Development and Metabolism of In Vitro Fertilization-Derived Feline Embryos Relative to Embryos Grown In Vivo 1. Biol. Reprod. 2007, 76, 858–870. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pope, C.E.; Gómez, M.C.; Dresser, B.L. In Vitro Production and Transfer of Cat Embryos in the 21st Century. Theriogenology 2006, 66, 59–71. [Google Scholar] [CrossRef]
- Wolfe, B.A.; Wild, D.E. Development to Blastocysts of Domestic Cat Oocytes Matured and Fertilized In Vitro after Prolonged Cold Storage. J. Reprod. Fertil. 1996, 106, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Zahmel, J.; Jänsch, S.; Jewgenow, K.; Luvoni, G.C. Inhibition of Apoptotic Pathways Improves DNA Integrity but Not Developmental Competence of Domestic Cat Immature Vitrified Oocytes. Front. Vet. Sci. 2020, 7, 588334. [Google Scholar] [CrossRef] [PubMed]
- Pope, C.E.; Gómez, M.C.; Kagawa, N.; Kuwayama, M.; Leibo, S.P.; Dresser, B.L. In Vivo Survival of Domestic Cat Oocytes after Vitrification, Intracytoplasmic Sperm Injection and Embryo Transfer. Theriogenology 2012, 77, 531–538. [Google Scholar] [CrossRef]
- Ochota, M.; Wojtasik, B.; Niżański, W. Survival Rate after Vitrification of Various Stages of Cat Embryos and Blastocyst with and without Artificially Collapsed Blastocoel Cavity. Reprod. Domest. Anim. 2017, 52, 281–287. [Google Scholar] [CrossRef]
- Sowińska, N.; Zahmel, J.; Niżański, W.; Hribal, R.; Fernandez-Gonzalez, L.; Jewgenow, K. Meiotic Status Does Not Affect the Vitrification Effectiveness of Domestic Cat Oocytes. Animals 2020, 10, 1371. [Google Scholar] [CrossRef]
- Morselli, M.G.; Luvoni, G.C.; Comizzoli, P. The Nuclear and Developmental Competence of Cumulus-Oocyte Complexes Is Enhanced by Three-Dimensional Coculture with Conspecific Denuded Oocytes during in Vitro Maturation in the Domestic Cat Model. Reprod. Domest. Anim. 2017, 52 (Suppl. S2), 82–87. [Google Scholar] [CrossRef]
- Klaus, C.; Eder, S.; Franz, C.; Müller, K. Successful Cryopreservation of Domestic Cat (Felis Catus) Epididymal Sperm after Slow Equilibration to 15 or 10 °C. Reprod. Domest. Anim. 2016, 51, 195–203. [Google Scholar] [CrossRef]
- Buranaamnuay, K. Protocols for Sperm Cryopreservation in the Domestic Cat: A Review. Anim. Reprod. Sci. 2017, 183, 56–65. [Google Scholar] [CrossRef]
- Herrick, J.R. In Vitro Culture of Embryos from Domestic Cats. Methods Mol. Biol. 2019, 2006, 229–246. [Google Scholar] [CrossRef]
- Pope, C.E.; Gomez, M.C.; Dresser, B.L. In Vitro Embryo Production and Embryo Transfer in Domestic and Non-Domestic Cats. Theriogenology 2006, 66, 1518–1524. [Google Scholar] [CrossRef] [PubMed]
- Jewgenow, K.; Zahmel, J. Preservation of Female Genetic Resources in Feline Species. Theriogenology 2020, 156, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Swanson, W.F. Research in Nondomestic Species: Experiences in Reproductive Physiology Research for Conservation of Endangered Felids. ILAR J. 2003, 44, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Luther, I.; Jakop, U.; Lueders, I.; Tordiffe, A.; Franz, C.; Schiller, J.; Kotze, A.; Müller, K. Semen Cryopreservation and Radical Reduction Capacity of Seminal Fluid in Captive African Lion (Panthera leo). Theriogenology 2017, 89, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Shivaji, S.; Kholkute, S.D.; Verma, S.K.; Gaur, A.; Umapathy, G.; Singh, A.; Sontakke, S.; Shailaja, K.; Reddy, A.; Monika, S.; et al. Conservation of Wild Animals by Assisted Reproduction and Molecular Marker Technology. Indian J. Exp. Biol. 2003, 41, 710–723. [Google Scholar]
- Armstrong, D.L.; Crichton, E.G.; Dankof, S.M.; Schwalbach, L.M.J.; Gardner, D.K.; Loskutoff, N.M. 200Ovarian Stimulation, Laparoscopic Oocyte Retrieval, Ivf and Blastocyst Production Using Sequential Media in the African Lion (Panthera leo). Reprod. Fertil. Dev. 2004, 16, 221. [Google Scholar] [CrossRef]
- Fernandez-Gonzalez, L.; Hribal, R.; Stagegaard, J.; Zahmel, J.; Jewgenow, K. Production of Lion (Panthera leo) Blastocysts after Invitro Maturation of Oocytes and Intracytoplasmic Sperm Injection. Theriogenology 2015, 83, 995–999. [Google Scholar] [CrossRef]
- Zahmel, J.; Jänsch, S.; Jewgenow, K.; Sandgreen, D.M.; Skalborg Simonsen, K.; Colombo, M. Maturation and Fertilization of African Lion (Panthera leo) Oocytes after Vitrification. Cryobiology 2021, 98, 146–151. [Google Scholar] [CrossRef]
- Zahmel, J.; Fernandez-Gonzalez, L.; Jewgenow, K.; Müller, K. Felid-Gamete-Rescue within EAZA-Efforts and Results in Biobanking Felid Oocytes and Sperm. J. Zoo Aquar. Res. 2019, 7, 15–24. [Google Scholar] [CrossRef]
- Ringleb, J.; Waurich, R.; Wibbelt, G.; Streich, W.J.; Jewgenow, K. Prolonged Storage of Epididymal Spermatozoa Does Not Affect Their Capacity to Fertilise in Vitro-Matured Domestic Cat (Felis Catus) Oocytes When Using ICSI. Reprod. Fertil. Dev. 2011, 23, 818–825. [Google Scholar] [CrossRef] [PubMed]
- Hribal, R.; Jewgenow, K.; Braun, B.C.; Comizzoli, P. Influence of Culture Medium Composition on Relative MRNA Abundances in Domestic Cat Embryos. Reprod. Domest. Anim. 2013, 48, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Hribal, R.; Hachen, A.; Jewgenow, K.; Zahmel, J.; Fernandez-Gonzalez, L.; Braun, B.C. The Influence of Recombinant Feline Oviductin on Different Aspects of Domestic Cat (Felis Catus) IVF and Embryo Quality. Theriogenology 2014, 82, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Jewgenow, K.; Fernandez-Gonzalez, L.; Jänsch, S.; Viertel, D.; Zahmel, J. Brilliant Cresyl Blue Staining Allows the Selection for Developmentally Competent Immature Feline Oocytes. Theriogenology 2019, 126, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.-C.; Zahmel, J.; Jewgenow, K.; Comizzoli, P. Desiccated Cat Spermatozoa Retain DNA Integrity and Developmental Potential after Prolonged Storage and Shipping at Non-Cryogenic Temperatures. J. Assist. Reprod. Genet. 2022, 39, 141–151. [Google Scholar] [CrossRef]
- Fernandez-Gonzalez, L.; Kozhevnikova, V.; Brusentsev, E.; Jänsch, S.; Amstislavsky, S.; Jewgenow, K. IGF-I Medium Supplementation Improves Singly Cultured Cat Oocyte Maturation and Embryo Development In Vitro. Animals 2021, 11, 1909. [Google Scholar] [CrossRef]
- Moghadam, A.R.E.; Moghadam, M.T.; Hemadi, M.; Saki, G. Oocyte Quality and Aging. JBRA Assist. Reprod. 2022, 26, 105–122. [Google Scholar] [CrossRef]
- Nagano, M.; Uchikura, K.; Takahashi, Y.; Hishinuma, M. Effect of Duration of in Vitro Maturation on Nuclear Maturation and Fertilizability of Feline Oocytes. Theriogenology 2008, 69, 231–236. [Google Scholar] [CrossRef]
- Kochan, J.; Nowak, A.; Młodawska, W.; Prochowska, S.; Partyka, A.; Skotnicki, J.; Niżański, W. Comparison of the Morphology and Developmental Potential of Oocytes Obtained from Prepubertal and Adult Domestic and Wild Cats. Animals 2021, 11, 20. [Google Scholar] [CrossRef]
- Cao, G.-Z.; Cui, X.-Y.; Lian, W.; Miao, H.-Y. Developmental Potential of Prepubertal Mouse Oocytes Is Compromised Due Mainly to Their Impaired Synthesis of Glutathione. PLoS ONE 2013, 8, 58018. [Google Scholar] [CrossRef]
- Gandolfi, F.; Milanesi, E.; Pocar, P.; Luciano, A.M.; Brevini, T.A.L.; Acocella, F.; Lauria, A.; Armstrong, D.T. Comparative Analysis of Calf and Cow Oocytes during in Vitro Maturation. Mol. Reprod. Dev. 1998, 49, 168–175. [Google Scholar] [CrossRef]
- Morton, K.M. Developmental Capabilities of Embryos Produced in Vitro from Prepubertal Lamb Oocytes. Reprod. Domest. Anim. 2008, 43 (Suppl. S2), 137–143. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Case, A. Advanced Reproductive Age and Fertility. J. Obstet. Gynaecol. Can. 2011, 33, 1165–1175. [Google Scholar] [CrossRef]
- Cimadomo, D.; Fabozzi, G.; Vaiarelli, A.; Ubaldi, N.; Ubaldi, F.M.; Rienzi, L. Impact of Maternal Age on Oocyte and Embryo Competence. Front. Endocrinol. 2018, 9, 327. [Google Scholar] [CrossRef]
- Johnston, L.A.; Donoghue, A.M.; O’Brien, S.J.; Wildt, D.E. Rescue and Maturation in Vitro of Follicular Oocytes Collected from Nondomestic Felid Species. Biol. Reprod. 1991, 45, 898–906. [Google Scholar] [CrossRef]
- Wildt, D.E. Genome Resource Banking for Wildlife Research, Management, and Conservation. ILAR J. 2000, 41, 228–234. [Google Scholar] [CrossRef]
- Comizzoli, P.; Holt, W.V. Recent Advances and Prospects in Germplasm Preservation of Rare and Endangered Species. Adv. Exp. Med. Biol. 2014, 753, 331–356. [Google Scholar] [CrossRef]
- Colombo, M.; Zahmel, J.; Binder, C.; Herbel, J.; Luvoni, G.C.; Jewgenow, K. Ovary Cold Storage and Shipment Affect Oocyte Yield and Cleavage Rate of Cat Immature Vitrified Oocytes. Cryobiology 2021, 98, 181–186. [Google Scholar] [CrossRef]
- Piras, A.R.; Burrai, G.; Pietro, A.F.; Falchi, L.; Zedda, M.T.; Pau, S.; Gadau, S.D.; Antuofermo, E.; Bebbere, D.; Ledda, S.; et al. Structure of Preantral Follicles, Oxidative Status and Developmental Competence of in Vitro Matured Oocytes after Ovary Storage at 4 °C in the Domestic Cat Model. Reprod. Biol. Endocrinol. 2018, 16, 76. [Google Scholar] [CrossRef]
- Wood, T.C.; Montali, R.J.; Wildt, D.E. Follicle-Oocyte Atresia and Temporal Taphonomy in Cold-Stored Domestic Cat Ovaries. Mol. Reprod. Dev. 1997, 46, 190–200. [Google Scholar] [CrossRef]
- Malo, A.F.; Martinez-Pastor, F.; Olivier, F.; Spies, T.; Roldan, E.R.S.; Bartels, P. 208Effect of Refrigeration and Cryopreservation on the Quality of Lion Epididymal Spermatozoa. Reprod. Fertil. Dev. 2004, 16, 225–226. [Google Scholar] [CrossRef]
- Nowak, A.; Kochan, J.; Świętek, E.; Kij, B.; Prochowska, S.; Witarski, W.; Bugno-Poniewierska, M.; Niżański, W. Survivability and Developmental Competences of Domestic Cat (Felis Catus) Oocytes after Cryotech Method Vitrification. Reprod. Domest. Anim. 2020, 55, 992–997. [Google Scholar] [CrossRef] [PubMed]
- Mokrousova, V.I.; Okotrub, K.A.; Brusentsev, E.Y.; Kizilova, E.A.; Surovtsev, N.V.; Amstislavsky, S.Y. Effects of Slow Freezing and Vitrification on Embryo Development in Domestic Cat. Reprod. Domest. Anim. 2020, 55, 1328–1336. [Google Scholar] [CrossRef] [PubMed]
- Luvoni, G.C. Gamete Cryopreservation in the Domestic Cat. Theriogenology 2006, 66, 101–111. [Google Scholar] [CrossRef]
- Katska-Ksiazkiewicz, L.; Ryńska, B.; Kania, G.; Smorag, Z.; Gajda, B.; Pieńkowski, M. Timing of Nuclear Maturation of Nonstored and Stored Domestic Cat Oocytes. Theriogenology 2003, 59, 1567–1574. [Google Scholar] [CrossRef]
- Zahmel, J.; Mundt, H.; Jewgenow, K.; Braun, B.C. Analysis of Gene Expression in Granulosa Cells Post-Maturation to Evaluate Oocyte Culture Systems in the Domestic Cat. Reprod. Domest. Anim. 2017, 52, 65–70. [Google Scholar] [CrossRef]
- Saeki, K.; Nagao, Y.; Hoshi, M.; Kainuma, H. Effects of Cumulus Cells on Sperm Penetration of Bovine Oocytes in Protein-Free Medium. Theriogenology 1994, 42, 1115–1123. [Google Scholar] [CrossRef]
- Wu, A.; Anupriwan, A.; Iamsaard, S.; Chakrabandhu, K.; Santos, D.C.; Rupar, T.; Tsang, B.K.; Carmona, E.; Tanphaichitr, N. Sperm Surface Arylsulfatase A Can Disperse the Cumulus Matrix of Cumulus Oocyte Complexes. J. Cell. Physiol. 2007, 213, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Gadella, B.M.; Boerke, A. An Update on Post-Ejaculatory Remodeling of the Sperm Surface before Mammalian Fertilization. Theriogenology 2016, 85, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Somfai, T.; Kikuchi, K.; Kaneda, M.; Akagi, S.; Watanabe, S.; Mizutani, E.; Haraguchi, S.; Dang-Nguyen, T.Q.; Inaba, Y.; Geshi, M.; et al. Cytoskeletal Abnormalities in Relation with Meiotic Competence and Ageing in Porcine and Bovine Oocytes during in Vitro Maturation. Anat. Histol. Embryol. 2011, 40, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.B.; Ho, J.R.; Cortessis, V.; Chen, I.J.; Bendikson, K.A.; Paulson, R.J.; McGinnis, L.K.; Ahmady, A. What Is the Optimal Timing of Intracytoplasmic Sperm Injection (ICSI) after EGG Retrieval? A Randomized Controlled Trial. J. Assist. Reprod. Genet. 2021, 38, 2151–2156. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hamano, K.; Qian, X.Q.; Funauchi, K.; Furudate, M.; Minato, Y. Oocyte Activation and Parthenogenetic Development of Bovine Oocytes Following Intracytoplasmic Sperm Injection. Zygote 1999, 7, 233–237. [Google Scholar] [CrossRef] [PubMed]


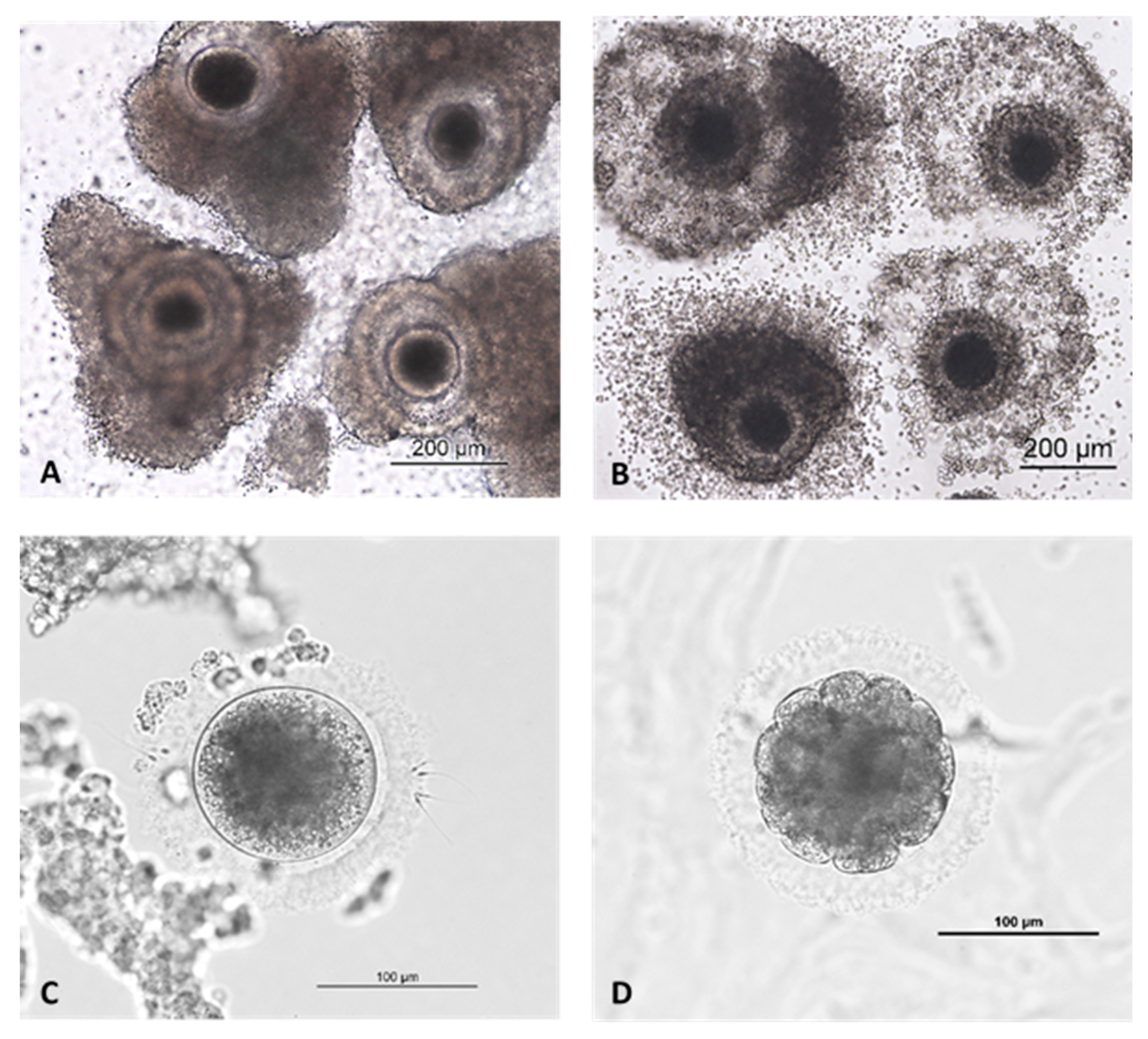{kind=link}
| African Lion | Year | Age (years) | Transport (h) | n Oocytes | Duration of IVM (h) | n Matured Oocytes | Maturation Rate (%) | n Embryos | Ferilization Rate (%) | ICSI/IVF | Sperm Source | Utilized Media |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2007 | 5 | 28 | 22 | 36 | 6 | 27.3 | 0 | 0 | IVF | lion cryo | A1, C, E |
| 2 | 2008 | 15 | 24 | 31 | 36 | 2 | 6.5 | − | − | − | − | A1 (IVM only) |
| 3 | 2008 | 1 | 24 | 31 | 36 | 15 | 48.4 | − | − | − | − | A1 (IVM only) |
| 4 | 2009 | 13 | 5 | 4 | 36 | 0 | 0 | − | − | − | − | A1, C, E |
| 5 | 2009 | 7 | 10 | 36 | A1, E | |||||||
| 6 | 7 | 10 | 71 | 19 | 26.8 | 0 | 0 | ICSI | lion cryo | |||
| 7 | 7 | 10 | ||||||||||
| 8 | 2010 | 1 | 6 | 40 | 34 | 8 | 20 | 1 | 12.5 | ICSI | lion cryo | A1, E |
| 9 | 1 | 6 | ||||||||||
| 10 | 2010 | 5 | 24 | 26 | 28 | 5 | 19.2 | 2 | 40 | ICSI | tiger fresh | A1, E |
| 11 | 2011 | 8 | 10 | 35 | 0 | A1, C, E | ||||||
| 12 | 7 | 10 | 37 | 18 | 48.6 | 0 | IVF | lion cryo | ||||
| 13 | 6 | 10 | ||||||||||
| 14 | 2012 | 1 | 24 | 4 | 24 | 1 | 25 | 0 | 0 | ICSI | lion cryo | A1, E |
| 15 | 2012 | 1 | 24 | 3 | 24 | 0 | 0 | − | − | − | − | A1 |
| 16 | 2012 | 14 | 10 | 36 | 9.1 | |||||||
| 17 | 6 | 10 | 55 | 11 | 20 | 1 | ICSI | lion cryo | A1, E | |||
| 18 | 6 | 10 | ||||||||||
| 19 | 2013 | 1 | 10 | 70 | 34 | 25 | 35.7 | 11 | 44 | ICSI | lion cryo | A1, A2, F |
| 20 | 1 | 10 | ||||||||||
| 21 | 1 | 10 | ||||||||||
| 22 | 1 | 10 | ||||||||||
| 23 | 2014 | 0.75 | 24 | 10 | 28 | 4 | 40 | 0 | 0 | ICSI | lion cryo | A1, F |
| 24 | 2015 | 1 | 10 | 66 | 35 | 49 | lion cryo/ DC fresh | A1, C, F | ||||
| 25 | 1 | 10 | 74.2 | 6 | 12.2 | ICSI/IVF | ||||||
| 26 | 1 | 10 | ||||||||||
| 27 | 2016 | 14 | 10 | 14 | 31 | 6 | 42.9 | 2 | 33.3 | IVF | lion cryo UC | A1, D, F |
| 28 | 2017 | 7 | 8 | 4 | 30 | 1 | 25 | 0 | 0 | ICSI | lion cryo | A1, F |
| 29 | 2018 | 11 | 28 | 23 | 25 | 5 | 21.7 | 0 | 0 | IVF | lion cryo | A1, D, F |
| 30 | 2019 | 7 | 24 | 8 | 24 | 3 | 37.5 | 0 | 0 | ICSI | lion cryo | A2, F |
| 31 | 2019 | 6 | 20 | 2 | 28 | 0 | 0 | − | − | − | − | A1 |
| 32 | 2019 | 7 | 0 | 9 * | 32−34 | 5 | 55.6 | 0 | 0 | ICSI | lion cryo | B, F |
| 33 | 2019 | 7 | 0 | 9 * | 32−34 | 1 | 11.1 | 0 | 0 | ICSI | lion cryo | B, F |
| 34 | 2019 | 5 | 0 | 28 * | 32−34 | 17 | 60.7 | 1 | 5.9 | ICSI | lion cryo | B, F |
| 35 | 2019 | 5 | 0 | 14 * | 32−34 | 10 | 71.4 | 2 | 20 | ICSI | lion cryo | B, F |
| 36 | 2020 | 1 | 0 | 54 | 30 | 8 | 44 | 0 | 0 | ICSI | lion fresh | B, F |
| 37 | 2021 | 13 | 0 | 10 | 28 | 5 | 50.0 | 3 | 60 | IVF | lion cryo | B, D, F |
| 38 | 2021 | 9 | 0 | 72 | 28 | 48 | 66.7 | 10 | 20.8 | IVF | lion cryo/fresh | B, D, F |
| 39 | 2021 | 9 | 0 | 17 | 28 | 15 | 88.2 | 3 | 20 | IVF | lion fresh | B, D, F |
| 40 | 2021 | 1 | 0 | 47 | 26/32 | 23 | 48.9 | 5 | 21.7 | IVF | lion cryo | B, D, G |
| 41 | 2021 | 1 | 0 | 23 | 29 | 14 | 60.9 | 6 | 42.9 | ICSI | lion cryo | B, G |
| 42 | 2021 | 1 | 0 | 70 | 26/32 | 31 | 44.3 | 11 | 35.5 | IVF | lion cryo | B, D, G |
| Total | Ø 5.3 | 874 (933) | 355 | 40.6 | 64 | 18.9 |
| Event | Basic Medium | Supplements | References |
|---|---|---|---|
| Transport: | |||
| T1 | Physiological saline solution | - | |
| T2 | Hepes-MEM (M7278) | 3 mg/mL BSA, 1:100 (v:v) Antibiotic Antimycotic Solution | [21] |
| Washing medium: | |||
| WM | M199 (M4530) | 3 mg/mL BSA, 1.4 mg/mL HEPES, 0.6 mg/mL sodium lactate, 0.25 mg/mL sodium pyruvate, 0.15 mg/mL L-glutamine, 0.1 mg/mL cysteine and 0.055 mg/mL gentamicin | [21] |
| IVM: | |||
| A1 | M199 (M4530) | WM + 0.05 IU/ml luteinizing hormone (LH) + 0.02 IU/ml pituitary follicle-stimulating hormone (FSH) | [21] |
| A2 | M199 (M4530) | WM + 10x LH and FSH | [22] |
| B | Quinn’s Advantage Protein Plus™ Blastocyst Medium (ART-1529) | + 10x LH and FSH | [19] |
| IVF: | |||
| C | Tyrode’s salts solution (T2397) | 6 mg/mL BSA, 1.2 mg/mL HEPES, 1.1 mg/mL sodium lactate, 0.1 mg/mL sodium pyruvate, 0.15 mg/mL L-glutamine, 2.2 IU/mL Heparin | [23] |
| D | M199 (M4530) | WM + 2.2 IU/mL Heparin | [24] |
| IVC: | |||
| E | M16 | 3 mg/mL BSA, 0.03 mg/ml gentamicin, 0.1 mM non-essential amino acids | [21] |
| F | HAM’s F10 (N6013) | 5% FBS, 0.11 mg/mL sodium pyruvate, 0.15 mg/mL L-glutamine, 0.06 mg/ml gentamicin | [22] |
| G | Quinn’s Advantage Protein Plus™ Blastocyst Medium (ART-1529) | 5% FBS | [25] |
| Sperm media: | |||
| H | Tyrode’s salts solution (T2397) | 6 mg/mL BSA, 1.2 mg/mL HEPES, 1.1 mg/mL sodium lactate, 0.1 mg/mL sodium pyruvate, 0.15 mg/mL L-glutamine | [21] |
| I | M199 (M7528) | [9] | |
| Mineral oil: | |||
| Sigma-Aldrich (M8410) | [21] | ||
| Reprodline Medical GmbH (REF451200) | [24] | ||
| Cooper Surgical (ART-4008-5) | not published |
| Lionesses | n | No. of Oocytes | No. of Oocytes/Animal | Maturation Rate | Cleavage Rate |
|---|---|---|---|---|---|
| Prepubertals | 17 | 418 | 24.6 | 42.6% a (178 of 418) | 24.5% a (40 of 163) |
| Adults | 25 | 515 | 20.6 | 38.8% a (177 of 456) | 13.7% b (24 of 175) |
| Total | 42 | 933 | 22.2 | 40.6% (355 of 874) | 18.9% (64 of 355) |
| Fertilization | n Lionesses | No. of Matured Oocytes | Cleavage Rate | Stage of Embryo | ||
|---|---|---|---|---|---|---|
| 2–16 Cells | Morula | Blastocyst | ||||
| Co-Culture | 14 | 183 | 20.2% a (37 of 183) | 33 [12] | 3 [3] | 1 [0] |
| ICSI | 26 | 155 | 17.4% a (27 of 155) | 22 [5] | 1 [0] | 4 [1] |
| Total | 40 | 338 | 18.9% (64 of 338) | 55 | 4 | 5 |
| Transport Duration (Location of Oocyte Collection) | n Lionesses | Mean Age of Lionesses | No. of Oocytes | Maturation Rate | Cleavage Rate |
|---|---|---|---|---|---|
| 0 h (on-site collection) | 11 | 5.3 | 412 | 50.1% (177 of 353) | 23.2% (41 of 177) |
| 10 h (IVF lab) | 21 | 5.3 | 361 | 38.0% (137 of 36) | 15.3% (21 of 137) |
| 24–28 h (IVF lab) | 10 | 5.3 | 160 | 25.6% (41 of 160) | 8.3% (2 of 24) |
| Maturation Time | n Lionesses | No. of Oocytes | Matured Oocytes n (%) | Cleaved Embryos n (%) |
|---|---|---|---|---|
| 24–28 h | 13 | 235 | 115 a (48.9) | 29 a (25.2) |
| 32–36 h | 28 | 544 | 211 b (38.8) | 27 b (12.8) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zahmel, J.; Skalborg Simonsen, K.; Stagegaard, J.; Palma-Vera, S.E.; Jewgenow, K. Current State of In Vitro Embryo Production in African Lion (Panthera leo). Animals 2022, 12, 1424. https://doi.org/10.3390/ani12111424
Zahmel J, Skalborg Simonsen K, Stagegaard J, Palma-Vera SE, Jewgenow K. Current State of In Vitro Embryo Production in African Lion (Panthera leo). Animals. 2022; 12(11):1424. https://doi.org/10.3390/ani12111424
Chicago/Turabian StyleZahmel, Jennifer, Kim Skalborg Simonsen, Julia Stagegaard, Sergio Eliseo Palma-Vera, and Katarina Jewgenow. 2022. "Current State of In Vitro Embryo Production in African Lion (Panthera leo)" Animals 12, no. 11: 1424. https://doi.org/10.3390/ani12111424
APA StyleZahmel, J., Skalborg Simonsen, K., Stagegaard, J., Palma-Vera, S. E., & Jewgenow, K. (2022). Current State of In Vitro Embryo Production in African Lion (Panthera leo). Animals, 12(11), 1424. https://doi.org/10.3390/ani12111424