
Pathological Findings in Eurasian Otters (Lutra lutra) Found Dead between 2015−2020 in Schleswig-Holstein, Germany

 , , ,
, , ,  , , ,
, , , 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
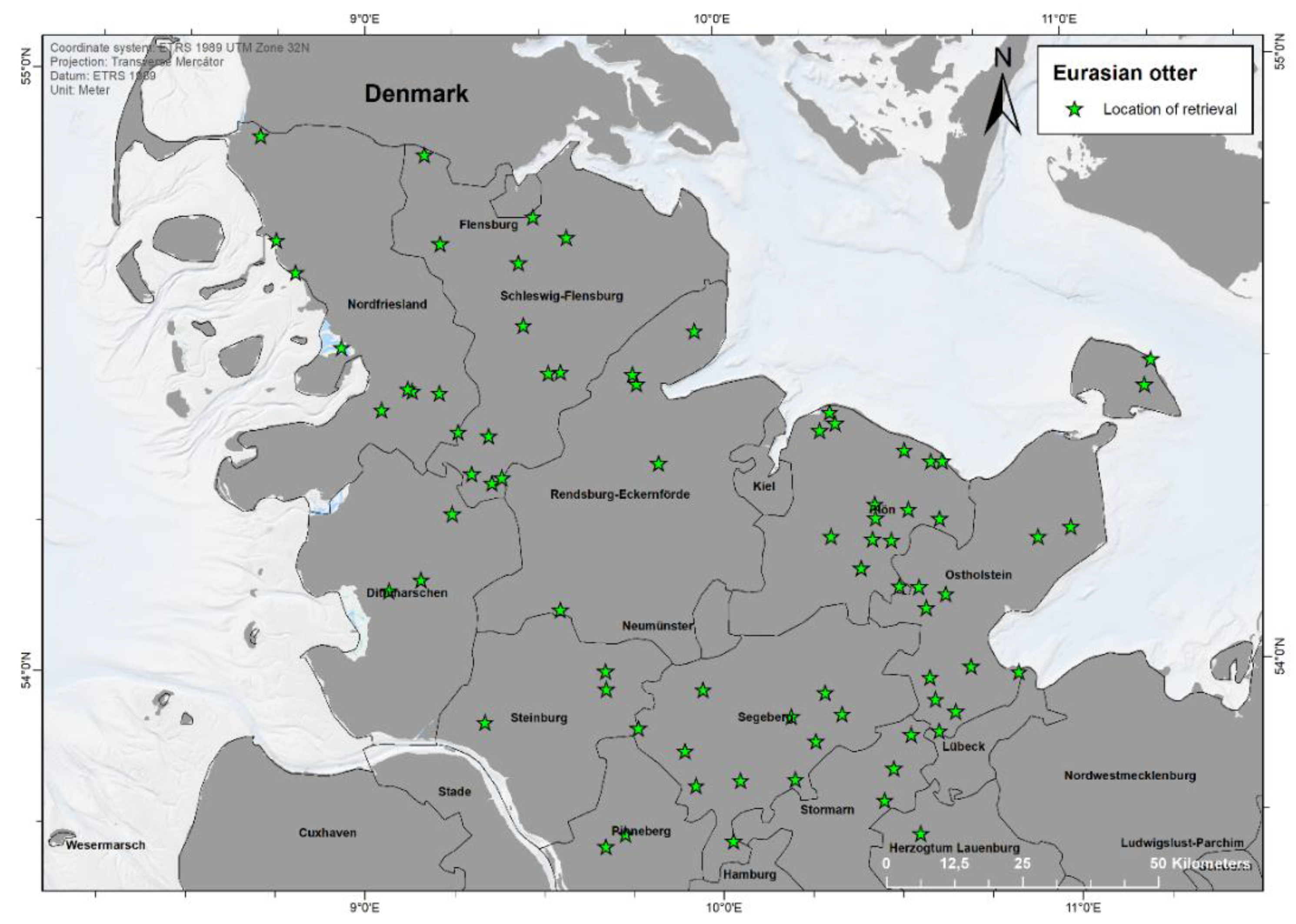
2.1. Network and Carcass Collection
2.2. Dissection
2.3. Histology
2.4. Mycology
2.5. Bacteriology
2.6. Virology
2.7. Parasitology
2.8. Age Determination
3. Results
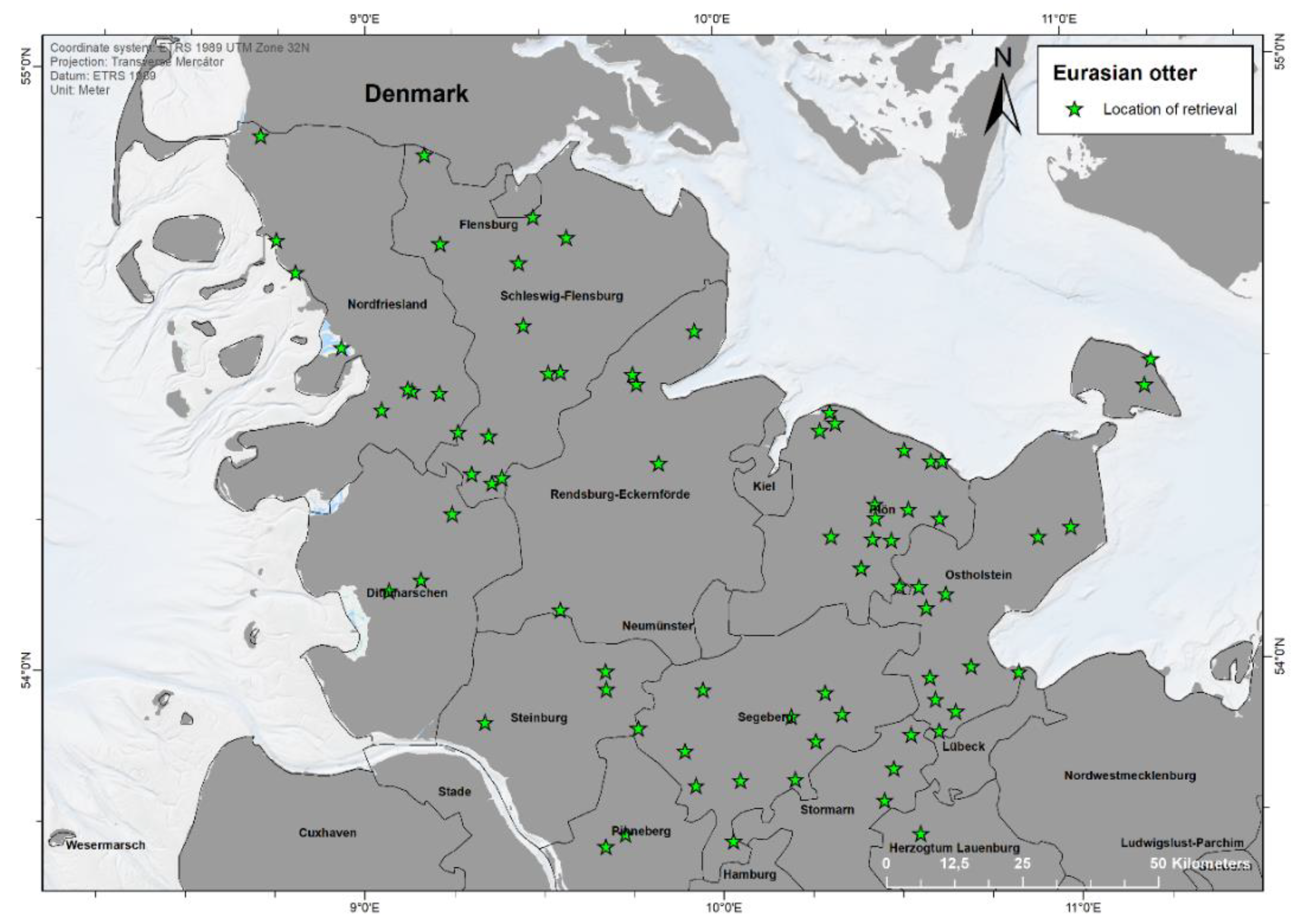
3.1. Location and Finding Data
3.2. Sex and Age
3.3. Pathological Findings
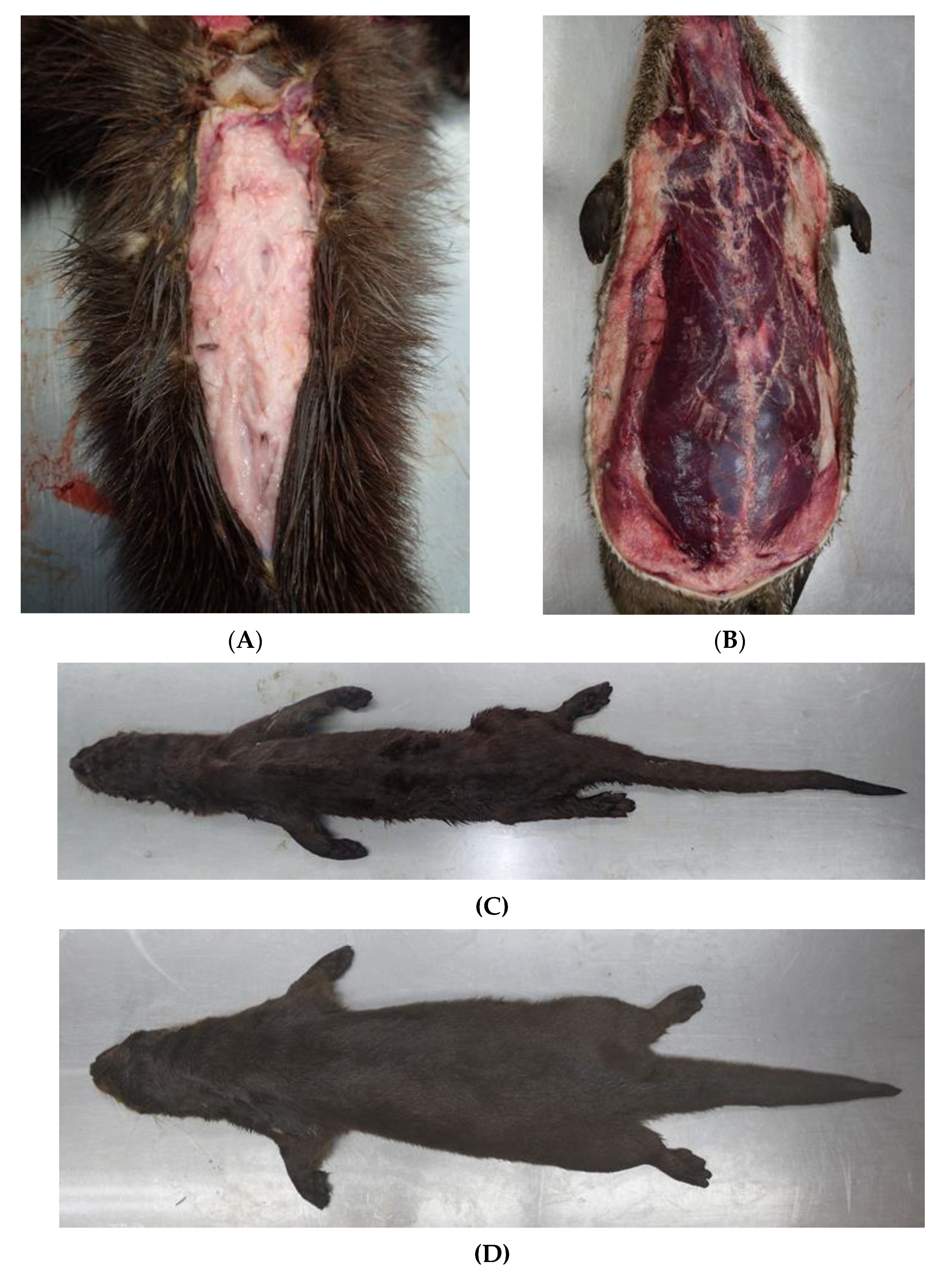
3.3.1. Nutritional Status
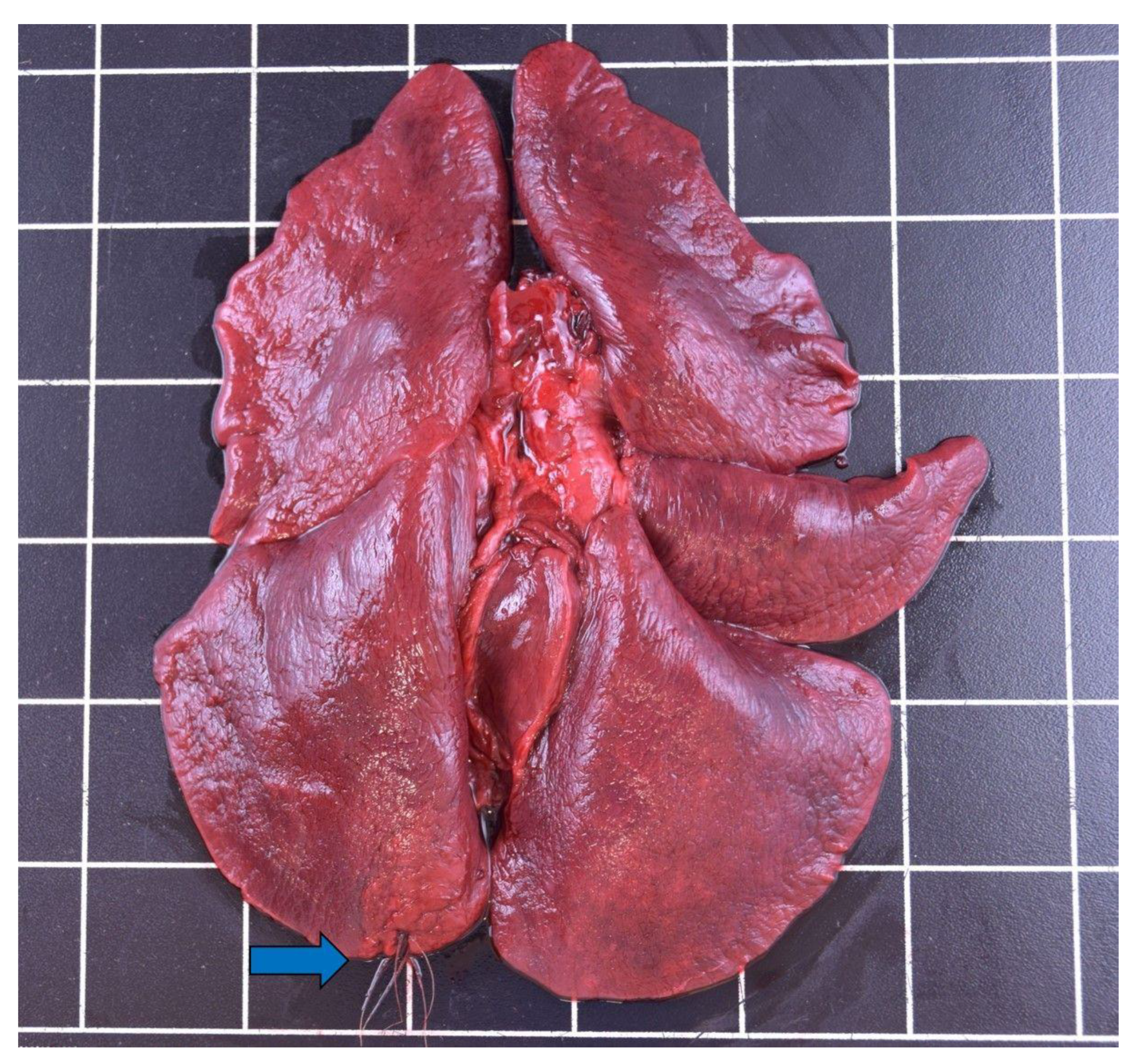
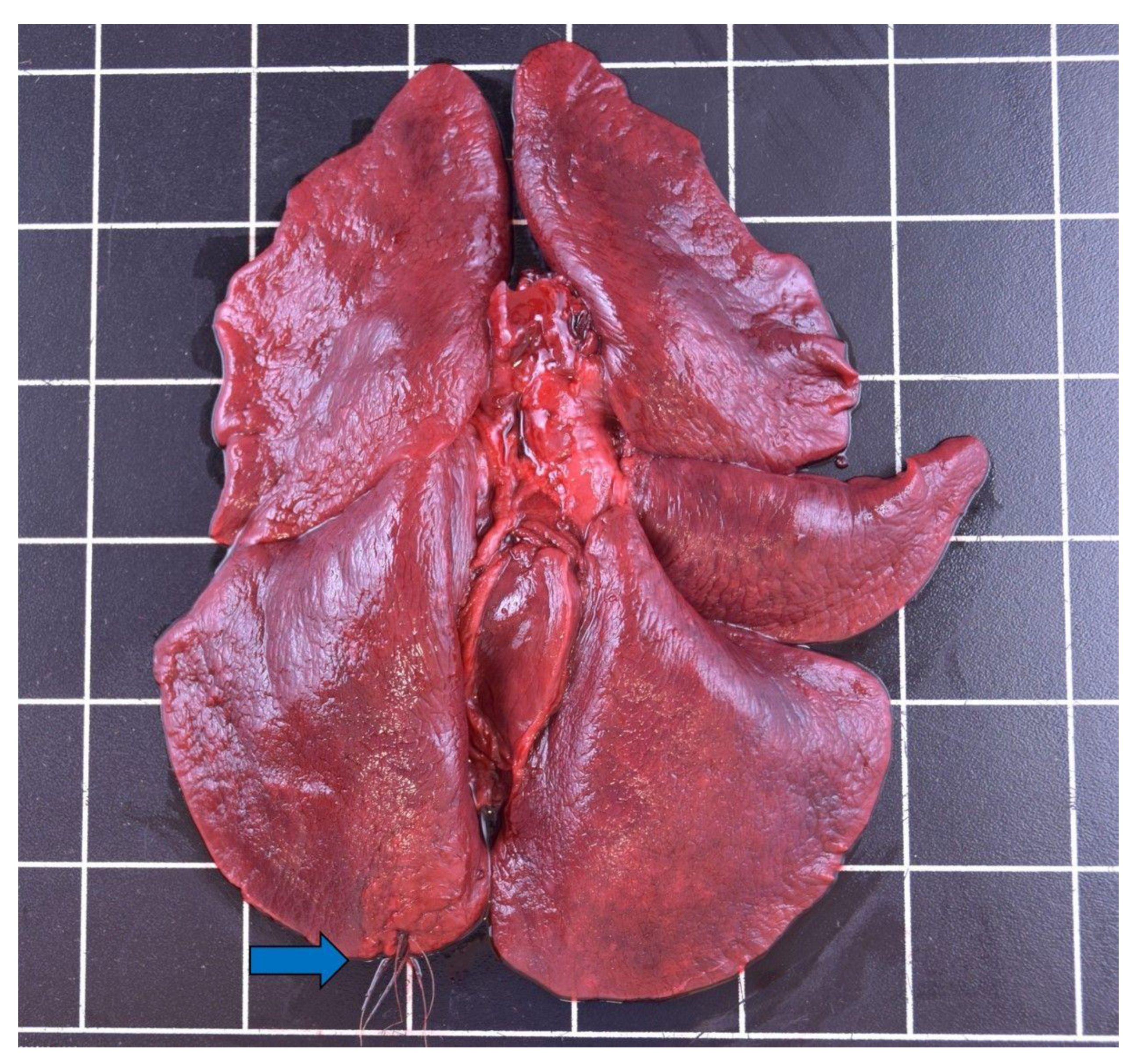
3.3.2. Respiratory Tract
3.3.3. Cardiovascular System
3.3.4. Thoracic Cavity
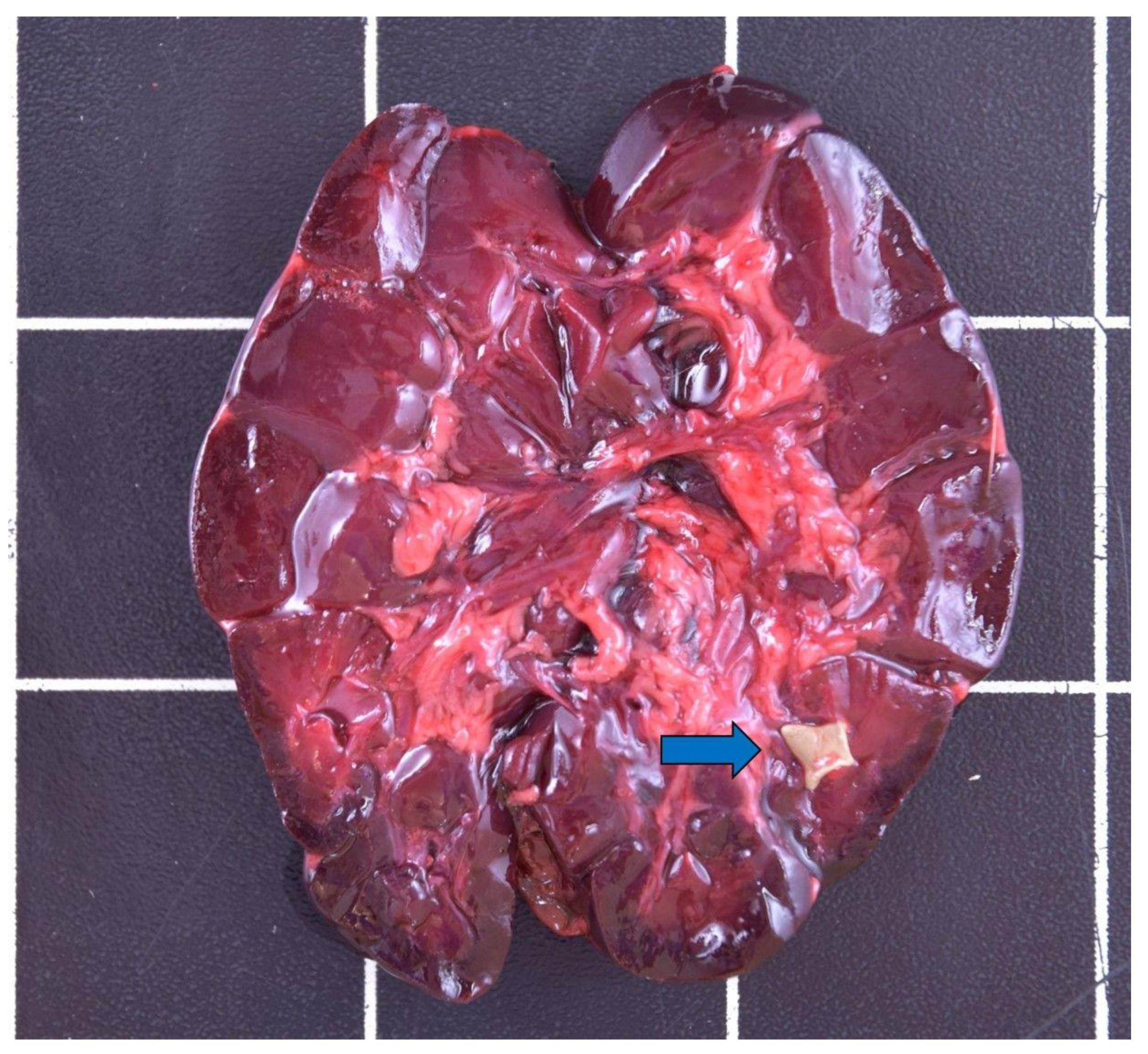
3.3.5. Alimentary Tract
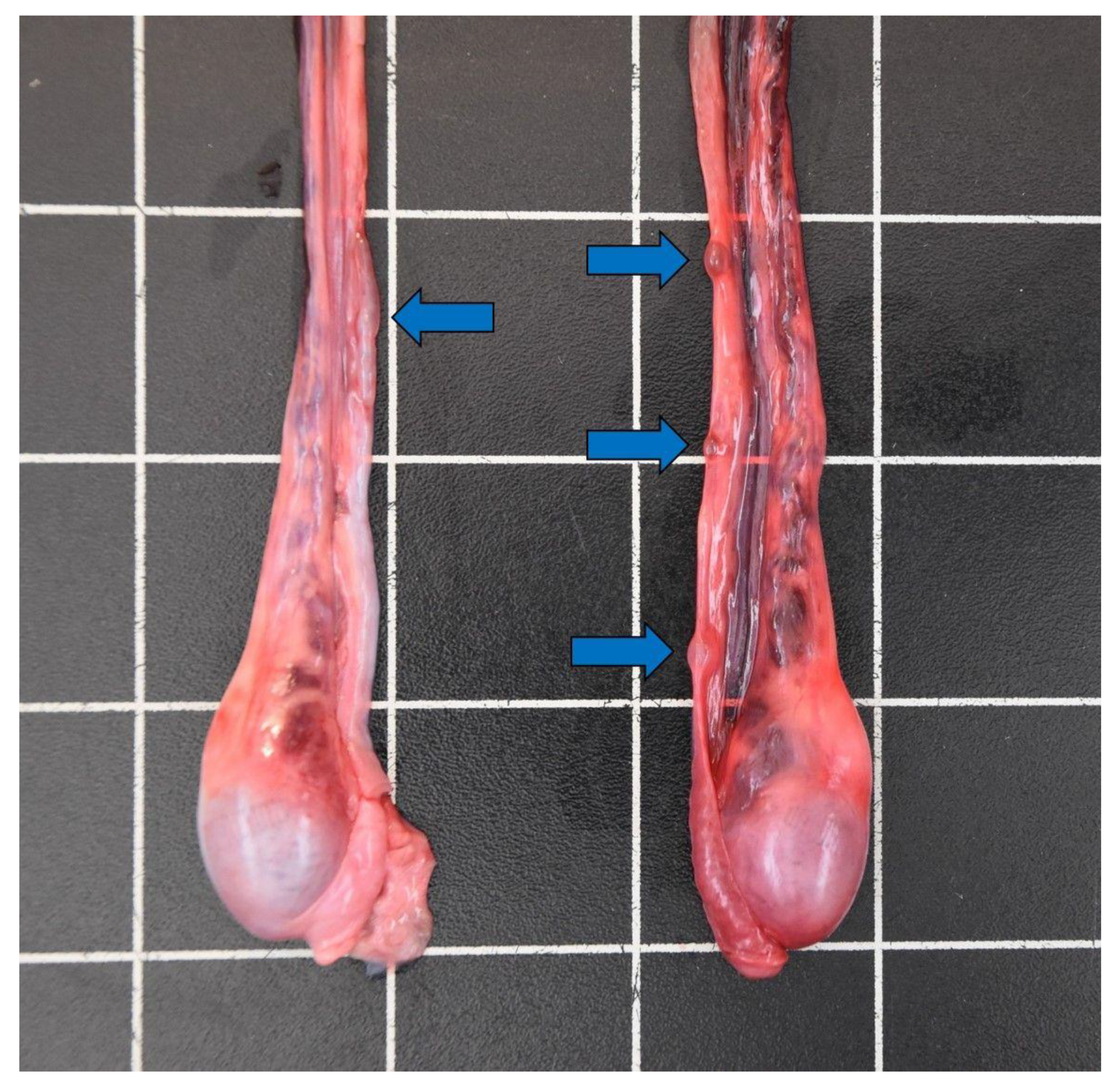
3.3.6. Urinary and Reproductive Tract
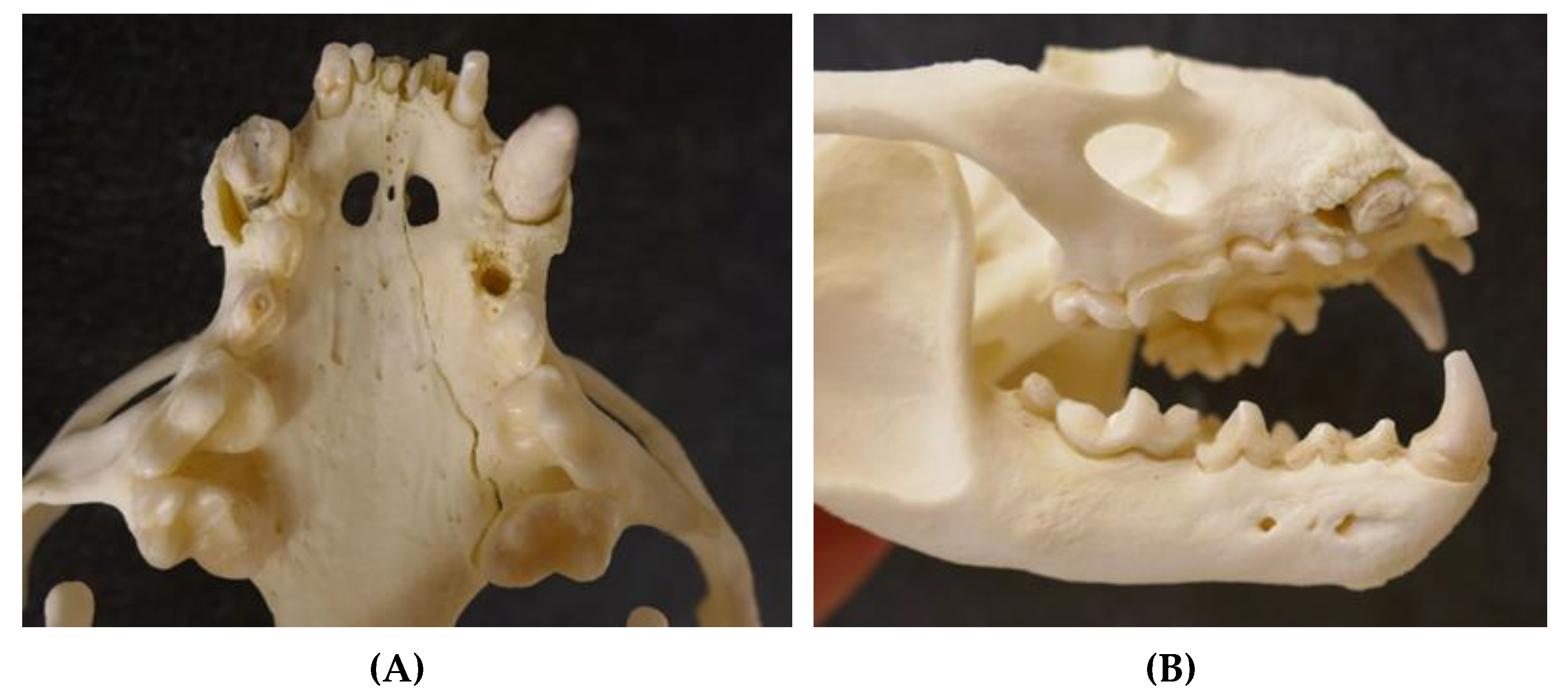
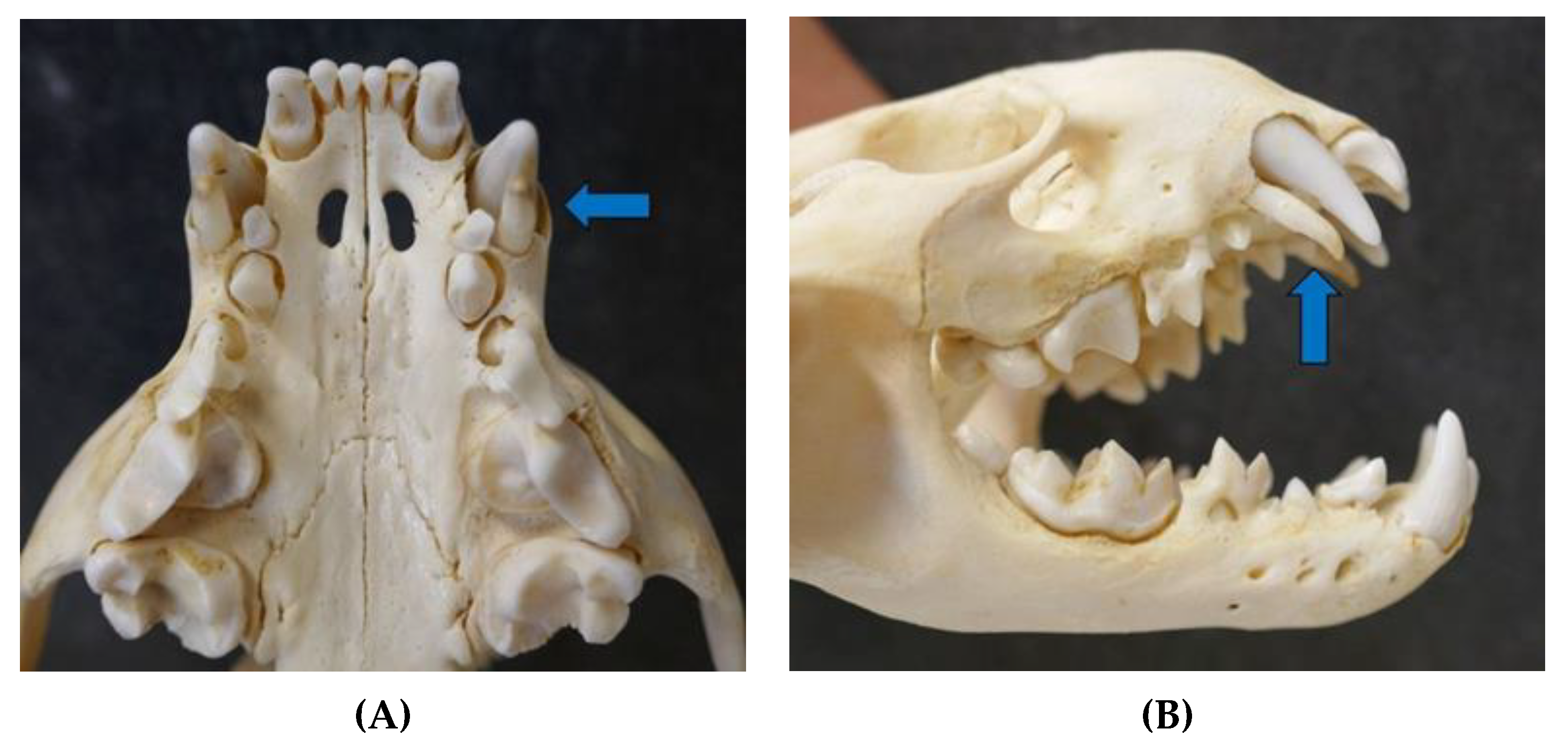
3.3.7. Skin and Subcutis
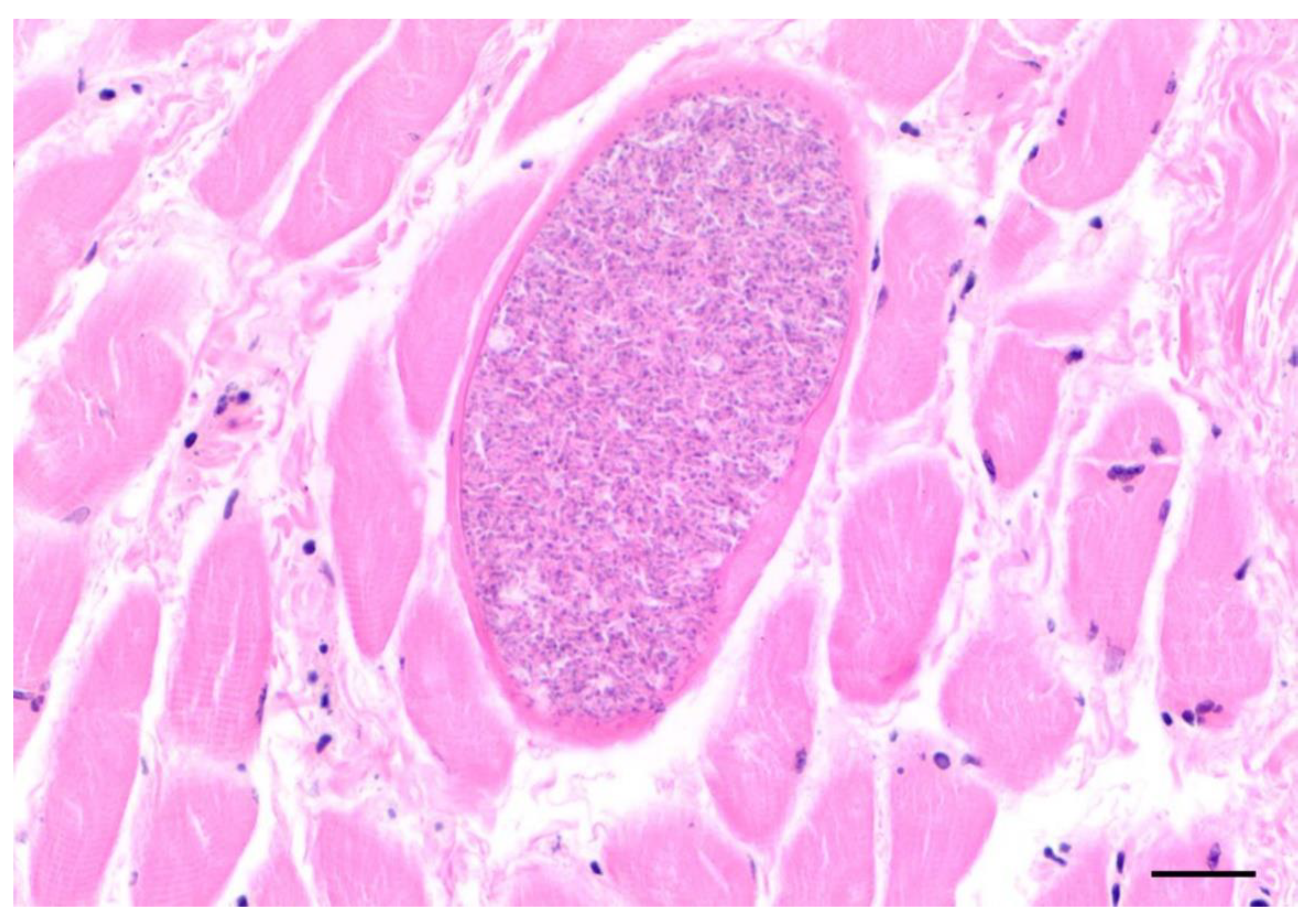
3.3.8. Musculoskeletal System
3.3.9. Central Nervous System, Eyes, and Ears
3.3.10. Hematopoietic and Endocrine System
3.4. Microbiology
3.5. Virology
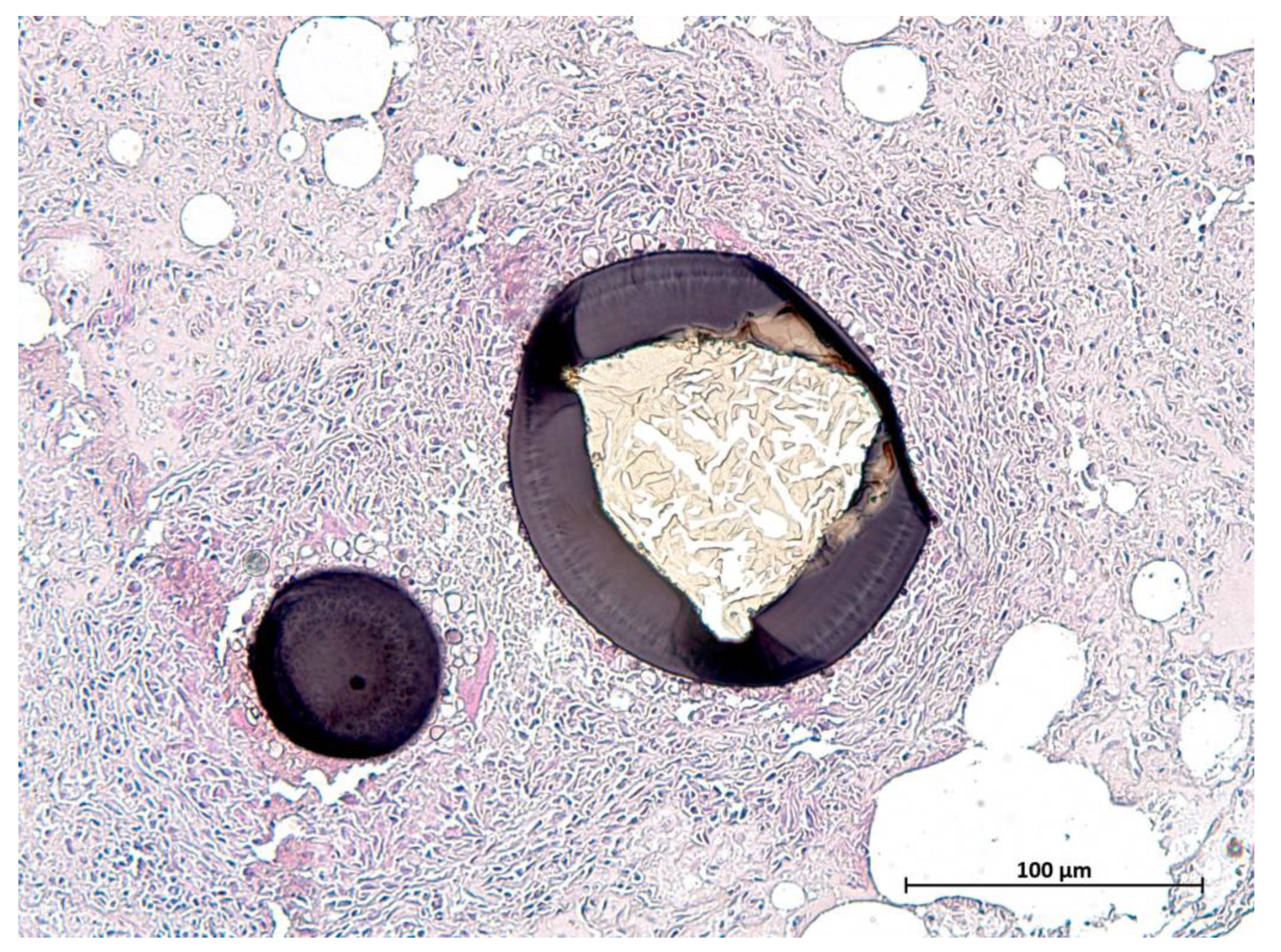
3.6. Parasitology
3.7. Causes of Death
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Conroy, J.W.H.; Chanin, P.R.F. The status of the Eurasian otter (Lutra lutra) in Europe- A review. J. Int. Otter Surviv. Fund 2000, 1, 7–28. [Google Scholar]
- Reuther, C. Fischotterschutz in Schleswig—Holstein; Ministerium für Umwelt, Natur und Forsten des Landes Schleswig-Holstein: Kiel, Germany, 2001. [Google Scholar]
- Duplaix, N.; Savage, M. The Global Otter Conservation Strategy; IUCN/SSC Otter Specialist Group: Salem, OR, USA, 2018. [Google Scholar]
- Grünwald-Schwark, V.; Zachos, F.E.; Honnen, A.-C.; Borkenhagen, P.; Krüger, F.; Wagner, J.; Drews, A.; Krekemeyer, A.; Schmüser, H.; Fichtner, A.; et al. Der Fischotter (Lutra lutra) in Schleswig-Holstein—Signatur Einer Rückwandernden, Bedrohten Wirbeltierart und Konsequenzen für den Naturschutz; Natur und Landschaft; Verlag, W., Ed.; Kohlhammer: Stuttgart, Germany, 2012; Volume 87. [Google Scholar]
- Jahresbericht 2020—Zur Biologischen Vielfalt, Jagd und Artenschutz; Ministry of Energy Transition, Agriculture, Environment, Nature and Digitalisation of Schleswig-Holstein (MELUND): Kiel, Germany, 2020.
- Honnen, A.C.; Petersen, B.; Kaßler, L.; Elmeros, M.; Roos, A.; Sommer, R.S.; Zachos, F.E. Genetic structure of Eurasian otter (Lutra lutra, Carnivora: Mustelidae) populations from the western Baltic sea region and its implications for the recolonization of north-western Germany. J. Zool. Syst. Evol. Res. 2010, 49, 169–175. [Google Scholar] [CrossRef]
- Bundestag. Gesetz über Naturschutz und Landschaftspflege (Bundesnaturschutzgesetz—BNatSchG); Bundesministerium der Justiz und für Verbraucherschutz: Berlin, Germany, 2021; pp. 1–67. [Google Scholar]
- EU Council Directive 92/43/EEC on the Conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Union 1992, 206, 7–50. [CrossRef]
- Roos, A.; Loy, A.; de Silva, P.; Hajkova, P.; Zemanova, B. Lutra lutra. IUCN Red List Threat Species. 2015. E.T12419A21935287. Available online: https://dx.doi.org/10.2305/IUCN.UK.2015-2.RLTS.T12419A21935287.en (accessed on 26 December 2021).
- Meinig, H.; Boye, P.; Dähne, M.; Hutterer, R.; Lang, J. Rote Liste und Gesamtartenliste der Säugetiere (Mammalia) Deutschlands. Nat. Und Biol. Vielfalt 2020, 170, 73. [Google Scholar] [CrossRef]
- Borkenhagen, P. Die Säugetiere Schleswig—Holsteins, Rote Liste; Ministerium für Energiewende, Landwirtschaft, Umwelt und ländliche Räume des Landes Schleswig-Holstein (MELUR): Kiel, Germany, 2014; p. 122. [Google Scholar]
- Kern, M. Kartierung zur Verbreitung des Fischotters (Lutra lutra) in Schleswig-Holstein nach der Stichprobenmethode des IUCN; Ministerium für Energiewende, Landwirtschaft, Umwelt und ländliche Räume des Landes Schleswig-Holstein (MELUR): Kiel, Germany, 2016. [Google Scholar]
- Newman, C.; Byrne, A.W. Musteloid diseases: Implications for conservation and species management. In Biology and Conservation of Musteloids; MacDonald, D.W., Newman, C., Harrington, L.A., Eds.; Oxford University Press: Oxford, UK, 2017; pp. 231–254. ISBN 978-0-19-875981-2. [Google Scholar]
- Chadwick, E. Post Mortem Study of Otters in England and Wales 1992–2003; Environment Agency: Bristol, UK, 2007. [Google Scholar]
- Simpson, V.R. Health Status of Otters in Southern and South West England 1996–2003; Environment Agency: Bristol, UK, 2007. [Google Scholar]
- Gutleb, A.C. Todesursachenforschung Fischotter, Grundlagen für ein Schutzkonzept von Lutra lutra L. 1758.pdf; WWF Austria: Vienna, Austria, 1994. [Google Scholar]
- Hauer, S.; Ansorge, H.; Zinke, O. Mortality patterns of otters (Lutra lutra) from eastern Germany. J. Zool. 2002, 256, 361–368. [Google Scholar] [CrossRef]
- Padalino, I.; Di Guardo, G.; Carbone, A.; Troiano, P.; Parisi, A.; Galante, D.; Cafiero, M.A.; Caruso, M.; Palazzo, L.; Guarino, L.; et al. Dolphin morbillivirus in Eurasian Otters, Italy. Emerg. Infect. Dis. 2019, 25, 372–374. [Google Scholar] [CrossRef]
- Madsen, A.B.; Dietz, H.H.; Henriksen, P.; Clausen, B. Survey of Danish free living otters Lutra lutra—A consecutive collection and necropsy of dead bodies. IUCN Otter Spec. Gr. Bull. 1999, 16, 65–75. [Google Scholar]
- Viscardi, M.; Santoro, M.; Clausi, M.T.; Cozzolino, L.; Decaro, N.; Colaianni, M.L.; Fusco, G. Molecular detection and characterization of carnivore parvoviruses in free-ranging Eurasian otters (Lutra lutra) in southern Italy. Transbound. Emerg. Dis. 2019, 66, 1864–1872. [Google Scholar] [CrossRef]
- Andersen, K.G.; Rambaut, A.; Lipkin, W.I.; Holmes, E.C.; Garry, R.F. The proximal origin of SARS-CoV-2. Nat. Med. 2020, 26, 450–452. [Google Scholar] [CrossRef] [Green Version]
- Molenaar, R.J.; Vreman, S.; Hakze-van der Honing, R.W.; Zwart, R.; de Rond, J.; Weesendorp, E.; Smit, L.A.M.; Koopmans, M.; Bouwstra, R.; Stegeman, A.; et al. Clinical and Pathological Findings in SARS-CoV-2 Disease Outbreaks in Farmed Mink (Neovison vison). Vet. Pathol. 2020, 57, 653–657. [Google Scholar] [CrossRef]
- Oliveira, M.; Sales-Luís, T.; Duarte, A.; Nunes, S.F.; Carneiro, C.; Tenreiro, T.; Tenreiro, R.; Santos-Reis, M.; Tavares, L.; Vilela, C.L. First assessment of microbial diversity in faecal microflora of Eurasian otter (Lutra lutra Linnaeus, 1758) in Portugal. Eur. J. Wildl. Res. 2008, 54, 245–252. [Google Scholar] [CrossRef]
- Lawson, P.A.; Foster, G.; Falsen, E.; Ohlen, M.; Collins, M.D. Vagococcus lutrae sp. nov., isolated from the common otter (Lutra lutra). Int. J. Syst. Bacteriol. 1999, 49, 1251–1254. [Google Scholar] [CrossRef]
- Simpson, V.R.; Gavier-Widen, D. Fatal adiaspiromycosis in a wild Eurasian otter (Lutra lutra). Vet. Rec. 2000, 147, 239–241. [Google Scholar] [CrossRef]
- Williams, B.H.; Burek-Huntington, K.A.; Miller, M. Mustelids. In Pathology of Wildlife and Zoo Animals; Terio, K.A., McAloose, D., St. Leger, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; p. 1084. [Google Scholar]
- Christian, A. Tick infestation (Ixodes) on the Eurasian Otter (Lutra lutra)—A long-term study. Soil Org. 2012, 84, 481–487. [Google Scholar]
- Sherrard-Smith, E.; Chadwick, E.; Cable, J. Abiotic and Biotic Factors Associated with Tick Population Dynamics on a Mammalian Host: Ixodes hexagonus Infesting Otters, Lutra lutra. PLoS ONE 2012, 7, e47131. [Google Scholar] [CrossRef] [Green Version]
- Wahlström, K.; Nikkilä, T.; Uggla, A. Sarcocystis species in skeletal muscle of otter (Lutra lutra). Parasitology 1999, 118, 59–62. [Google Scholar] [CrossRef]
- Máca, O. Molecular identification of Sarcocystis lutrae in the European otter (Lutra lutra) and the European badger (Meles meles) from the Czech Republic. Parasitol. Res. 2018, 117, 943–945. [Google Scholar] [CrossRef]
- Shimalov, V.V.; Shimalov, V.T.; Shimalov, A.V. Helminth fauna of otter (Lutra lutra Linnaeus, 1758) in Belorussian Polesie. Parasitol. Res. 2000, 86, 528. [Google Scholar] [CrossRef]
- Sommer, R.; Griesau, A.; Ansorge, H.; Priemer, J. Daten zur Populationsökologie des Fischotters Lutra lutra (Linnaeus, 1758) in Mecklenburg-Vorpommern; Beiträge zur Jagd- und Wildtierforschung; Gesellschaft für Wildtier- und Jagdforschung e.V.: Leipzig, Germany, 2005; Volume 30. [Google Scholar]
- Torres, J.; Feliu, C.; Fernández-Morán, J.; Ruíz-Olmo, J.; Rosoux, R.; Santos-Reis, M.; Miquel, J.; Fons, R. Helminth parasites of the Eurasian otter Lutra lutra in southwest Europe. J. Helminthol. 2004, 78, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Mulville, W. A Non-Invasive Investigation of the Intestinal Helminths of the Eurasian Otter, Lutra lutra, in Freshwater and Estuarine Habitats. Master’s Thesis, University of Dublin, Dublin, Ireland, 2016. [Google Scholar]
- Chadwick, E.A.; Cable, J.; Chinchen, A.; Francis, J.; Guy, E.; Kean, E.F.; Paul, S.C.; Perkins, S.E.; Sherrard-Smith, E.; Wilkinson, C.; et al. Seroprevalence of Toxoplasma gondii in the Eurasian otter (Lutra lutra) in England and Wales. Parasites Vectors 2013, 6, 75. [Google Scholar] [CrossRef] [Green Version]
- Sobrino, R.; Cabezón, O.; Millán, J.; Pabón, M.; Arnal, M.C.; Luco, D.F.; Gortázar, C.; Dubey, J.P.; Almeria, S. Seroprevalence of Toxoplasma gondii antibodies in wild carnivores from Spain. Vet. Parasitol. 2007, 148, 187–192. [Google Scholar] [CrossRef]
- Ansorge, H.; Schipke, R.; Zinke, O. Population structure of the otter, Lutra lutra. Parameters and model for a Central European region. Z. Fur Saugetierkd.-Int. J. Mamm. Biol. 1997, 62, 143–151. [Google Scholar]
- Siebert, U.; Wünschmann, A.; Weiss, R.; Frank, H.; Benke, H.; Frese, K. Post-mortem Findings in Harbour Porpoises (Phocoena phocoena) from the German North and Baltic Seas. J. Comp. Pathol. 2001, 124, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Heggberget, T.M. Reproduction in the Female European Otter in Central and Northern Norway. J. Mammal. 1988, 69, 164–167. [Google Scholar] [CrossRef]
- Chadwick, E.A.; Sherrard-Smith, E. Pregnancy among otters (Lutra lutra) found dead in England and Wales: Foetal development and lack os seasonality. IUCN Otter Spec. Group Bull. 2010, 27, 33–41. [Google Scholar]
- Siebert, U.; Wohlsein, P.; Lehnert, K.; Baumgärtner, W. Pathological Findings in Harbour Seals (Phoca vitulina): 1996–2005. J. Comp. Pathol. 2007, 137, 47–58. [Google Scholar] [CrossRef]
- Rooms, I.; Mugisha, P.; Gambichler, T.; Hadaschik, E.; Esser, S.; Rath, P.M.; Haase, G.; Wilmes, D.; McCormick-Smith, I.; Rickerts, V. Disseminated emergomycosis in a person with HIV infection, Uganda. Emerg. Infect. Dis. 2019, 25, 1750–1751. [Google Scholar] [CrossRef] [PubMed]
- Prenger-Berninghoff, E.; Siebert, U.; Stede, M.; König, A.; Weiß, R.; Baljer, G. Incidence of Brucella species in marine mammals of the German North Sea. Dis. Aquat. Organ. 2008, 81, 65–71. [Google Scholar] [CrossRef]
- Siebert, U.; Rademaker, M.; Ulrich, S.A.; Wohlsein, P.; Ronnenberg, K.; Prenger-Berninghoff, E. Bacterial Microbiota in Harbor Seals (Phoca Vitulina) from the North Sea of Schleswig-Holstein, Germany, around the Time of Morbillivirus and Influenza Epidemics. J. Wildl. Dis. 2017, 53, 201–214. [Google Scholar] [CrossRef]
- Bisping, W.; Amtsberg, G. Farbatlas zur Diagnose Bakterieller Infektionserreger der Tiere; Paul Parey: Berlin, Germany, 1988. [Google Scholar]
- Burkhardt, F. Mikrobiologische Diagnostik; G. Thieme Verlag: Stuttgart, Germany, 1992. [Google Scholar]
- Carter, G.R. Essentials of Veterinary Microbiology, 5th ed.; Williams & Wilkins: Baltimore, MD, USA, 1995. [Google Scholar]
- Streck, A.F.; Rüster, D.; Truyen, U.; Homeier, T. An updated TaqMan real-time PCR for canine and feline parvoviruses. J. Virol. Methods 2013, 193, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Müller, G.; Wohlsein, P.; Beineke, A.; Haas, L.; Greiser-Wilke, I.; Siebert, U.; Fonfara, S.; Harder, T.; Stede, M.; Gruber, A.D.; et al. Phocine distemper in German seals, 2002. Emerg. Infect. Dis. 2004, 10, 723–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.W.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eurosurveillance 2020, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, M.S.C.; Barr, B.C.; Marsh, A.E.; Anderson, M.L.; Rowe, J.D.; Tarantal, A.F.; Hendrickx, A.G.; Sverlow, K.; Dubey, J.P.; Conrad, P.A. Identification of bovine Neospora parasites by PCR amplification and specific small-subunit rRNA sequence probe hybridization. J. Clin. Microbiol. 1996, 34, 1203–1208. [Google Scholar] [CrossRef] [Green Version]
- Heggberget, T.M. Age determination in the European Otter (Lutra lutra). Zeitschrift für Säugetierkunde; Verlag Paul Parey: Hamburg/Berlin, Germany, 1984; Volume 49, pp. 299–305. [Google Scholar]
- Sherrard-Smith, E.; Chadwick, E.A. Age structure of the otter (Lutra lutra) population in England and Wales, and problems with cementum ageing. IUCN Otter Spec. Bull. 2010, 27, 42–49. [Google Scholar]
- Lippitsch, P.; Zinke, O.; Ansorge, H. Altersstruktur tot aufgefundener Fischotter aus Ostsachsen; Naturforschende Gesellschaft der Oberlausitz e.V.: Görlitz, Germany, 2017. [Google Scholar]
- Takeshige, A.; Nakano, M.; Kondoh, D.; Tanaka, Y.; Sekiya, A.; Yaguchi, T.; Furuoka, H.; Toyotome, T. Adiaspore development and morphological characteristics in a mouse adiaspiromycosis model. Vet. Res. 2020, 51, 119. [Google Scholar] [CrossRef] [PubMed]
- Chanin, P. Otter road casualties. Hystrix Ital. J. Mammal. 2006, 17, 79–90. [Google Scholar] [CrossRef]
- Heggberget, T.M.; Christensen, H. Reproductive Timing in Eurasian Otters on the Coast of Norway. Ecography 1994, 17, 339–348. [Google Scholar] [CrossRef]
- Simpson, V.R. Health status of otters (Lutra lutra) in south-west England based on postmortem findings. Vet. Rec. 1997, 141, 191–197. [Google Scholar] [CrossRef]
- Philcox, C.K.; Grogan, A.L.; Macdonald, D.W. Patterns of otter Lutra lutra road mortality in Britain. J. Appl. Ecol. 1999, 36, 748–762. [Google Scholar] [CrossRef]
- Heggberget, T.M. Otters and Roads. In Proceedings of the Eurasian Otter Workshop, Padula, Italy, 23 October 2005; pp. 42–43. [Google Scholar]
- Weber, A.; Trost, M. Die Säugetierarten der Fauna- Flora-Habitat-Richtlinie im Land Sachsen-Anhalt: Fischotter (Lutra lutra L., 1758); Landesamt für Umweltschutz Sachsen-Anhalt: Halle, Germany, 2015; Volume 1. [Google Scholar]
- Bradshaw, A.; Slater, F. A Postmortem Study of Otters (Lutra lutra) in England and Wales; Environment Agency: Bristol, UK, 2002. [Google Scholar]
- Lanszki, J.; Sugár, L.; Orosz, E.; Nagy, D. Biological data from post mortem analysis of otters in Hungary. Acta Zool. Acad. Sci. Hung. 2008, 54, 201–212. [Google Scholar]
- Buskirk, S.W.; Lindstedt, S.L. Sex Biases in Trapped Samples of Mustelidae. J. Mammal. 1989, 70, 88–97. [Google Scholar] [CrossRef]
- Ansorge, H. Notizen zur Altersbestimmung nach Wachstumslinien am Säugetierschädel. Methoden Feldökologischer Säugetierforsch. 1995, 1, 95–102. [Google Scholar]
- Hauer, S.; Ansorge, H.; Zinke, O. Reproductive performance of otters Lutra lutra (Linnaeus, 1758) in Eastern Germany: Low reproduction in a long-term strategy. Biol. J. Linn. Soc. 2002, 77, 329–340. [Google Scholar] [CrossRef]
- Kruuk, H.; Conroy, J.W.H. Mortality of Otters (Lutra lutra) in Shetland. J. Appl. Ecol. 1991, 28, 83–94. [Google Scholar] [CrossRef]
- Ruiz-Olmo, J.; Delibes, M.; Zapata, S. External morphometry, demography and mortality of the Otter Lutra lutra (Linneo, 1758) in the Iberian Peninsula. Galemys Boletín Inf. La Soc. Española Para La Conserv. Y Estud. Los Mamíferos 1998, 10, 239–251. [Google Scholar]
- Sidorovich, V.E. Structure, reproductive status and dynamics of the otter population in Byelorussia. Acta Theriol. 1991, 36, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Kruuk, H. Otters: Ecology, Behaviour and Conservation; Oxford University Press: Oxford, UK, 2006; ISBN 9780198565871. [Google Scholar]
- De Paoli, P. Biobanking in microbiology: From sample collection to epidemiology, diagnosis and research. FEMS Microbiol. Rev. 2005, 29, 897–910. [Google Scholar] [CrossRef] [Green Version]
- McGavin, D.M.; Zachary, J.F. Pathologie der Haustiere, 1st ed.; Elsevier GmbH: Munich, Germany, 2009; ISBN 978-3-437-58250-9. [Google Scholar]
- Blixenkrone-Møller, M.; Svansson, V.; Have, P.; Botner, A.; Nielsen, J. Infection studies in mink with seal-derived morbillivirus. Arch. Virol. 1989, 106, 165–170. [Google Scholar] [CrossRef]
- Williams, E.S.; Thorne, E.T.; Appel, M.J.G.; Belitsky, D.W. Canine Distemper in Black footed Ferrets (Mustela nigripes) from Wyoming. J. Wildl. Dis. 1988, 24, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Kimber, K.R.; Kollias, G.V.; Dubovi, E.J. Serologic survey of selected viral agents in recently captured wild north american river otters (lontra canadensis). J. Zoo Wildl. Med. 2000, 31, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Kimber, K.R.; Kollias, G.V. Infectious and parasitic diseases and contaminant-related problems of North American river otters (Lontra canadensis): A review. J. Zoo Wildl. Med. 2000, 31, 452–472. [Google Scholar] [CrossRef]
- Mos, L.; Ross, P.S.; McIntosh, D.; Raverty, S. Canine distemper virus in river otters in British Columbia as an emergent risk for coastal pinnipeds. Vet. Rec. 2003, 152, 237–239. [Google Scholar] [CrossRef]
- Wohlsein, P.; Puff, C.; Kreutzer, M.; Siebert, U.; Baumgärtner, W. Distemper—A permanent threat to marine mammals? Emerg. Infect. Dis. 2007, 13, 1959–1961. [Google Scholar] [CrossRef]
- Härkönen, T.; Dietz, R.; Reijnders, P.; Teilmann, J.; Harding, K.; Hall, A.; Brasseur, S.; Siebert, U.; Goodman, S.J.; Jepson, P.D.; et al. A review of the 1988 and 2002 phocine distemper virus epidemics in European harbour seals. Dis. Aquat. Organ. 2006, 68, 115–130. [Google Scholar] [CrossRef] [Green Version]
- Trent, R. Otters at Georgia Aquarium Test Positive for Coronavirus. Available online: https://edition.cnn.com/2021/04/19/us/georgia-aquarium-otters-covid-trnd/index.html (accessed on 18 August 2021).
- Kim, Y.I.; Kim, S.G.; Kim, S.M.; Kim, E.H.; Park, S.J.; Yu, K.M.; Chang, J.H.; Kim, E.J.; Lee, S.; Casel, M.A.B.; et al. Infection and Rapid Transmission of SARS-CoV-2 in Ferrets. Cell Host Microbe 2020, 27, 704–709. [Google Scholar] [CrossRef]
- Simpson, V. Diseases of otters in Britain. Proc. First Otter Toxicol. Conf. 2000, 27, 41–45. [Google Scholar]
- Simpson, V.R. Patterns and significance of bite wounds in Eurasian otters (Lutra Lutra) in southern and south-west England. Vet. Rec. 2006, 158, 113–119. [Google Scholar] [CrossRef]
- Borman, A.M.; JIang, Y.; Dukik, K.; Sigler, L.; Schwartz, I.S.; de Hoog, S. Adiaspiromycosis and Diseases Caused by Related Fungi in Ajellomycetaceae. In Emerging and Epizootic Fungal Infections in Animals; Springer Nature Switzerland AG: Cham, Switzerland, 2018; pp. 147–158. ISBN 9783319720937. [Google Scholar]
- Simpson, V.R.; Tomlinson, A.J.; Stevenson, K.; McLuckie, J.A.; Benavides, J.; Dagleish, M.P. A post-mortem study of respiratory disease in small mustelids in south-west England. BMC Vet. Res. 2016, 12, 72. [Google Scholar] [CrossRef]
- Krivanec, K.; Otcenasek, M.; Slais, J. Adiaspiromycosis in large free living carnivores. Mycopathologia 1976, 58, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Malatesta, D.; Simpson, V.R.; Fusillo, R.; Marcelli, M.; Bongiovanni, L.; Romanucci, M.; Palmieri, C.; Salda, L. Della First description of adiaspiromycosis in an Eurasian otter (Lutra lutra) in Italy. Vet. Ital. 2014, 50, 199–202. [Google Scholar] [CrossRef]
- Salomonsen, C.M.; Boye, M.; Høiby, N.; Jensen, T.H.; Hammer, A.S. Comparison of histological lesions in mink with acute hemorrhagic Pneumonia associated with pseudomonas aeruginosa or Escherichia coli. Can. J. Vet. Res. 2013, 77, 199–204. [Google Scholar]
- Mushin, R.; Ziv, G. An epidemiological study of Pseudomonas aeruginosa in cattle and other animals by pyocine typing. J. Hyg. 1973, 71, 113–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smallbone, W.A.; Chadwick, E.A.; Francis, J.; Guy, E.; Perkins, S.E.; Sherrard-Smith, E.; Cable, J. East-West Divide: Temperature and land cover drive spatial variation of Toxoplasma gondii infection in Eurasian otters (Lutra lutra) from England and Wales. Parasitology 2017, 144, 1433–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denk, D.; de Neck, S.; Khaliq, S.; Stidworthy, M.F. Toxoplasmosis in zoo species: A retrospective review. Anim. Under Revi. 2021; submitted. [Google Scholar]
- Sanders, C.W.; Olfenbuttel, C.; Pacifici, K.; Hess, G.R.; Livingston, R.S.; DePerno, C.S. Leptospira, parvovirus, and toxoplasma in the North American river otter (Lontra canadensis) in North Carolina, USA. J. Wildl. Dis. 2020, 56, 791–802. [Google Scholar] [CrossRef]
- Schulman, F.Y.; Montali, R.J.; Hauer, P.J. Gastroenteritis Associated with Clostridium perfringens Type A in Black-footed Ferrets (Mustela nigripes). Vet. Pathol. 1993, 30, 308–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolbiecki, L.; Izdebska, J.N. New data on the parasites of the Eurasian otter (Lutra lutra). Oceanol. Hydrobiol. Stud. 2014, 43, 1–6. [Google Scholar] [CrossRef]
- Radev, V.; Kanev, I.; Khrusanov, D.; Fried, B. Reexamination of the life cycle of Isthmiophora melis (Trematoda: Echinostomatidae) on material from southeast Europe. Parazitologiia 2009, 43, 445–453. [Google Scholar]
- Nugaraitė, D.; Mažeika, V.; Paulauskas, A. Helminths of mustelids with overlapping ecological niches: Eurasian otter Lutra lutra (Linnaeus, 1758), American mink Neovison vison Schreber, 1777, and European polecat Mustela putorius Linnaeus, 1758. Helminthologia 2019, 56, 66–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nugaraitė, D.; Mažeika, V.; Paulauskas, A. Helminths of mustelids (Mustelidae) in Lithuania. Biologija 2014, 60, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Dimitrova, Z.M.; Tzvetkov, Y.; Todev, I. Occurrence of acanthocephalans in the Eurasian otter Lutra lutra (L.) (Carnivora, Mustelidae) in Bulgaria, with a survey of acanthocephalans recorded from this host species. Helminthologia 2008, 45, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Weber, H.B. Untersuchungen zur Urolithiasis beim Eurasischen Fischotter, Lutra lutra. Ph.D.Thesis, University of Veterinary Medicine Hannover, Foundation, Hannover, Germany, 2001. [Google Scholar]
- Simpson, V.R.; Tomlinson, A.J.; Molenaar, F.M.; Lawson, B.; Rogers, K.D. Renal calculi in wild Eurasian otters (Lutra lutra) in England. Vet. Rec. 2011, 169, 49. [Google Scholar] [CrossRef]
- Roos, A.M.; Ågren, E.O. High prevalence of proposed müllerian duct remnant cysts on the spermatic duct in wild Eurasian otters (Lutra lutra) from Sweden. PLoS ONE 2013, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Elmeros, M.; Madsen, A.B. On the reproduction biology of otters (Lutra lutra) from Denmark. Z. Fur Saugetierkd. 1999, 64, 193–200. [Google Scholar]
- Hertweck, K.; Schipke, R. Wissenschaftliche Mitteilungen des Niederösterreichischen Landesmuseum; Amt der Niederösterreichischen Landesregierung, Abteilung Kultur und Wissenschaft: St. Pölten, Austria, 2001; pp. 175–181. [Google Scholar]
- Altintas, I.; Andrews, V.; Larsen, M.V. First reported human bloodstream infection with Vagococcus lutrae. New Microbes New Infect. 2020, 34, 100649. [Google Scholar] [CrossRef]
- Garcia, V.; Abat, C.; Rolain, J.M. Report of the first Vagococcus lutrae human infection, Marseille, France. New Microbes New Infect. 2016, 9, 56–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gjerde, B.; Josefsen, T.D. Molecular characterisation of Sarcocystis lutrae n. sp. and Toxoplasma gondii from the musculature of two Eurasian otters (Lutra lutra) in Norway. Parasitol. Res. 2015, 114, 873–886. [Google Scholar] [CrossRef] [PubMed]
- Prakas, P.; Strazdaitė-Žielienė, Ž.; Rudaitytė-Lukošienė, E.; Servienė, E.; Butkauskas, D. Molecular identification of Sarcocystis lutrae (Apicomplexa: Sarcocystidae) in muscles of five species of the family Mustelidae. Parasitol. Res. 2018, 117, 1989–1993. [Google Scholar] [CrossRef] [PubMed]
- Dolch, D.; Teubner, J.; Teubner, J. Haupttodesursachen des Fischotters—Lutra lutra (L., 1758) im Land Brandenburg in der Zeit von 1990 bis 1994. In Naturschutz und Landschaftspflege in Brandenburg; Landesamt für Umwelt Brandenburg (LfU): Potsdam, Germany, 1998; p. 101. [Google Scholar]
- Skarén, U. Analysis of One Hundred Otters Killed by Accidents in Central Finland. IUCN Otter Spec. Gr. Bull. 1992, 7, 9–12. [Google Scholar]
- Ansorge, H.; Fiedler, F.; Förster, F.; Geidezis, L.; Grohman, O.; Hempel, G.; Hertweck, K.; Jurisch, C.; Klenke, R.; Kubasch, H.; et al. Artenschutzprogramm Fischotter in Sachsen; Landesamt für Umwelt und Geologie: Radebeul, Germany, 1996. [Google Scholar]
- Ackermann, W.; Streitberger, M. Maßnahmenkonzepte für Ausgewählte Arten und Lebensraumtypen der FFH-Richtlinie Maßnahmenkonzepte für Ausgewählte Arten und Lebensraumtypen der FFH-Richtlinie; Bundesamt für Naturschutz (BfN): Bonn, Germany, 2016. [Google Scholar]
- Niedersächsischer Landesbetrieb für Wasserwirtschaft, Küsten- und Naturschutz (NLWKN). Niedersächsische Strategie zum Arten- und Biotopschutz Vollzugshinweise zum Schutz von Säugetierarten in Niedersachsen: Fischotter (Lutra lutra); NLWKN: Hannover, Germany, 2011. [Google Scholar]
- Striese, M.; Schreyer, R.M. Fischotter an Straßen—zur Passage von Brücken. In Tiere im Konflikt 1/1993 Fischotter Monitoring 1985–1991; Stubbe, M., Heidecke, D., Stubbe, A., Eds.; Martin-Luther-Universität Halle-Wittenberg: Halle, Germany, 1993; p. 84. [Google Scholar]
- Slater, F.M. An assessment of wildlife road casualties—The potential discrepancy between numbers counted and numbers killed. Web Ecol. 2002, 3, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Baker, P.J.; Harris, S.; Robertson, C.P.J.; Saunders, G.; White, P.C.L. Is it possible to monitor mammal population changes from counts of road traffic casualties? An analysis using Bristol’s red foxes Vulpes vulpes as an example. Mamm. Rev. 2004, 34, 115–130. [Google Scholar] [CrossRef]
- Weinberger, I.; Baumgartner, H. Der Fischotter—Ein heimlicher Jäger kehrt zurück, 1st ed.; Haupt Verlag: Bern, Austria, 2018; ISBN 978-3-258-08084-0. [Google Scholar]
- Brand, A.F.; Hynes, J.; Walker, L.A.; Glόria Pereira, M.; Lawlor, A.J.; Williams, R.J.; Shore, R.F.; Chadwick, E.A. Biological and anthropogenic predictors of metal concentration in the Eurasian otter, a sentinel of freshwater ecosystems. Environ. Pollut. 2020, 266, 9. [Google Scholar] [CrossRef] [PubMed]
- Westerberg, H.; Henle, K. Module 3: Diet and Damage Assessment. In Human—Wildlife Conflicts in Europe—Fisheries and Fisheating Vertebrates as a Model case; Klenke, R.A., Ring, I., Kranz, A., Jepsen, N., Rauschmayer, F., Henle, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 241–251. [Google Scholar]
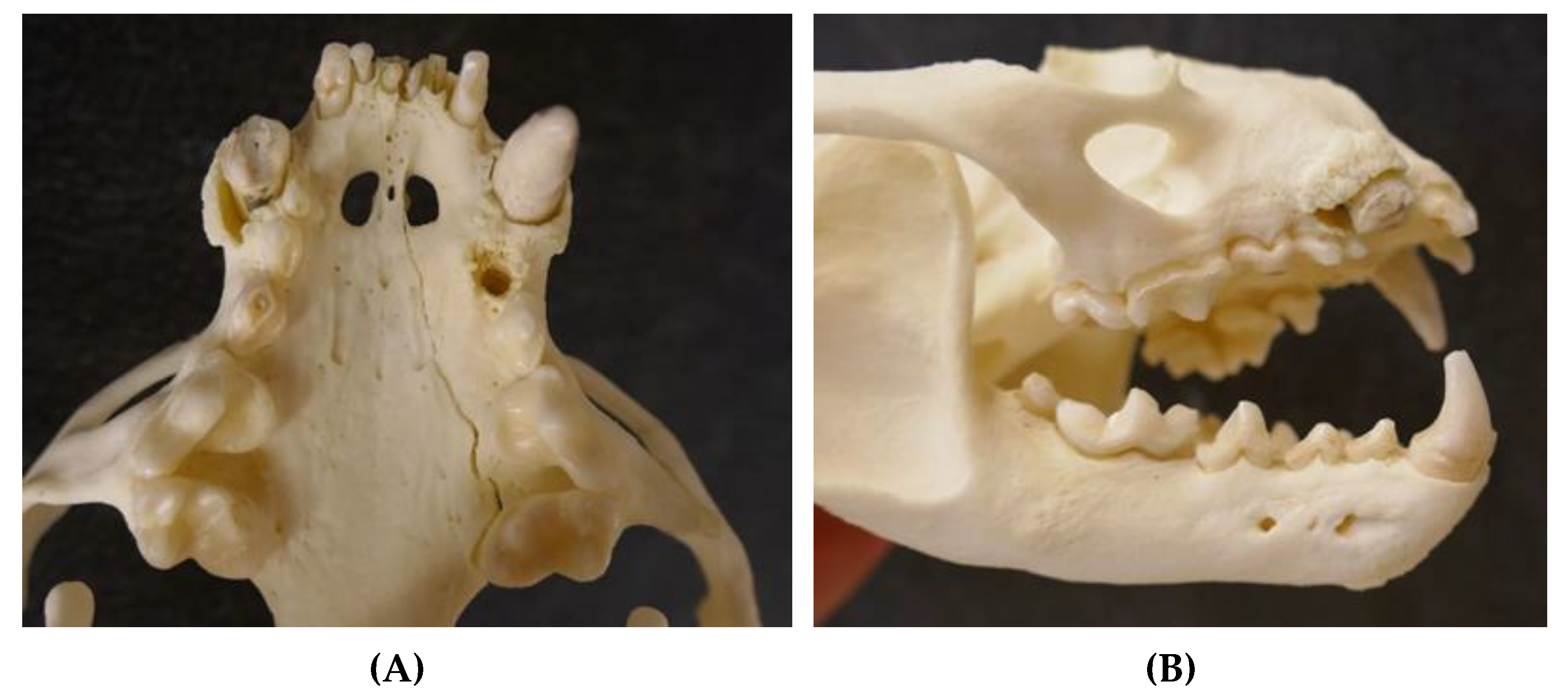

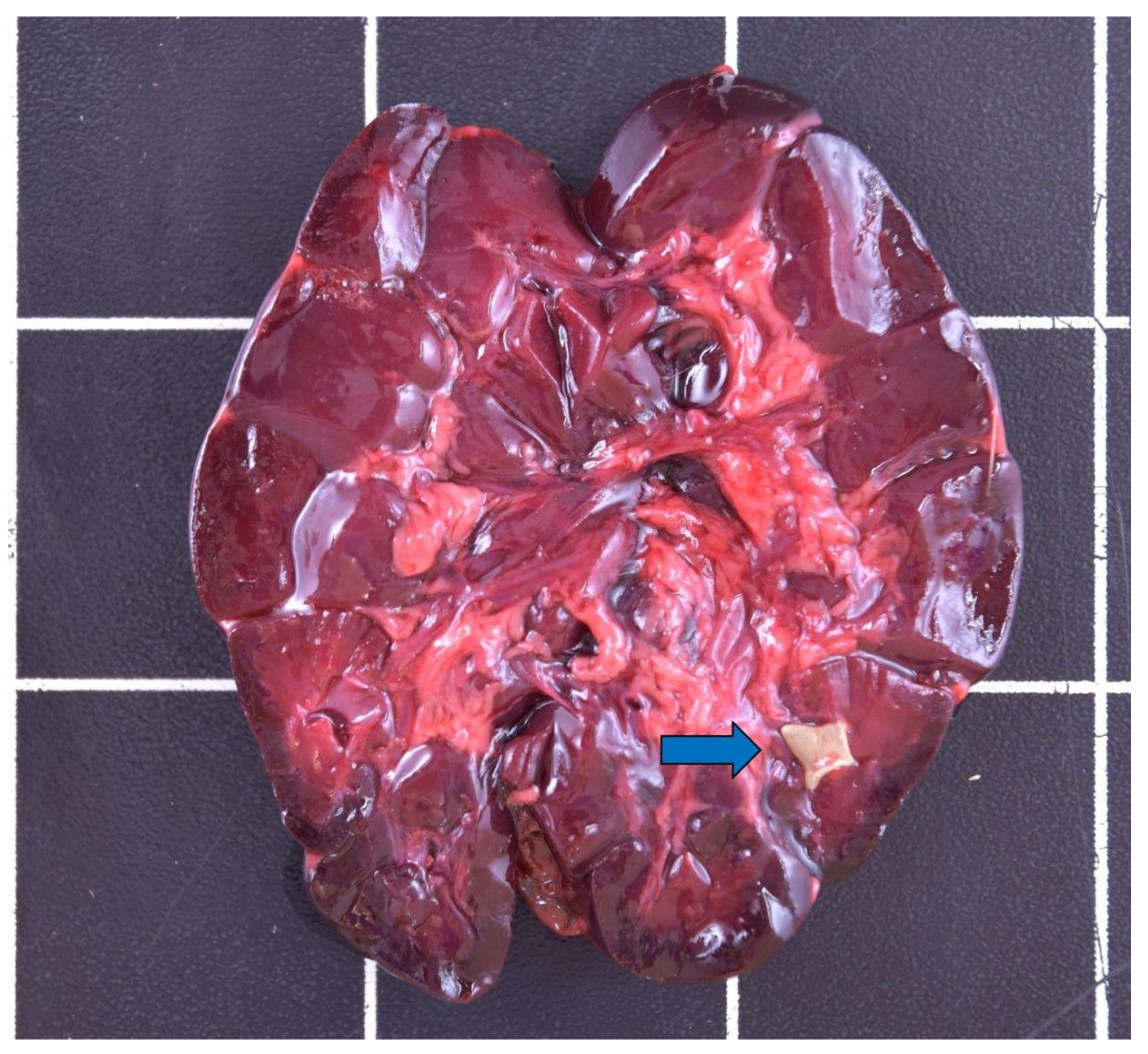
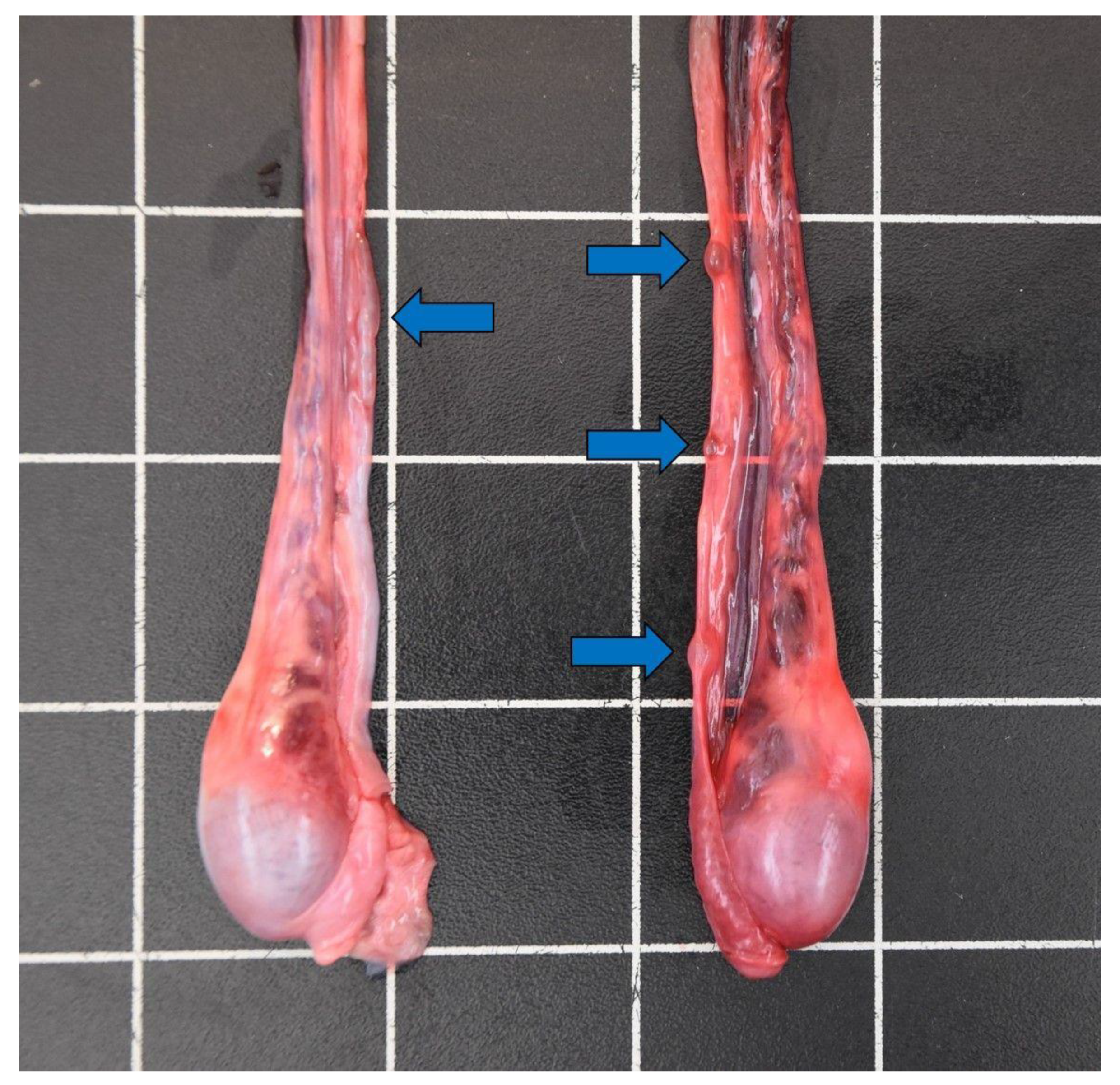
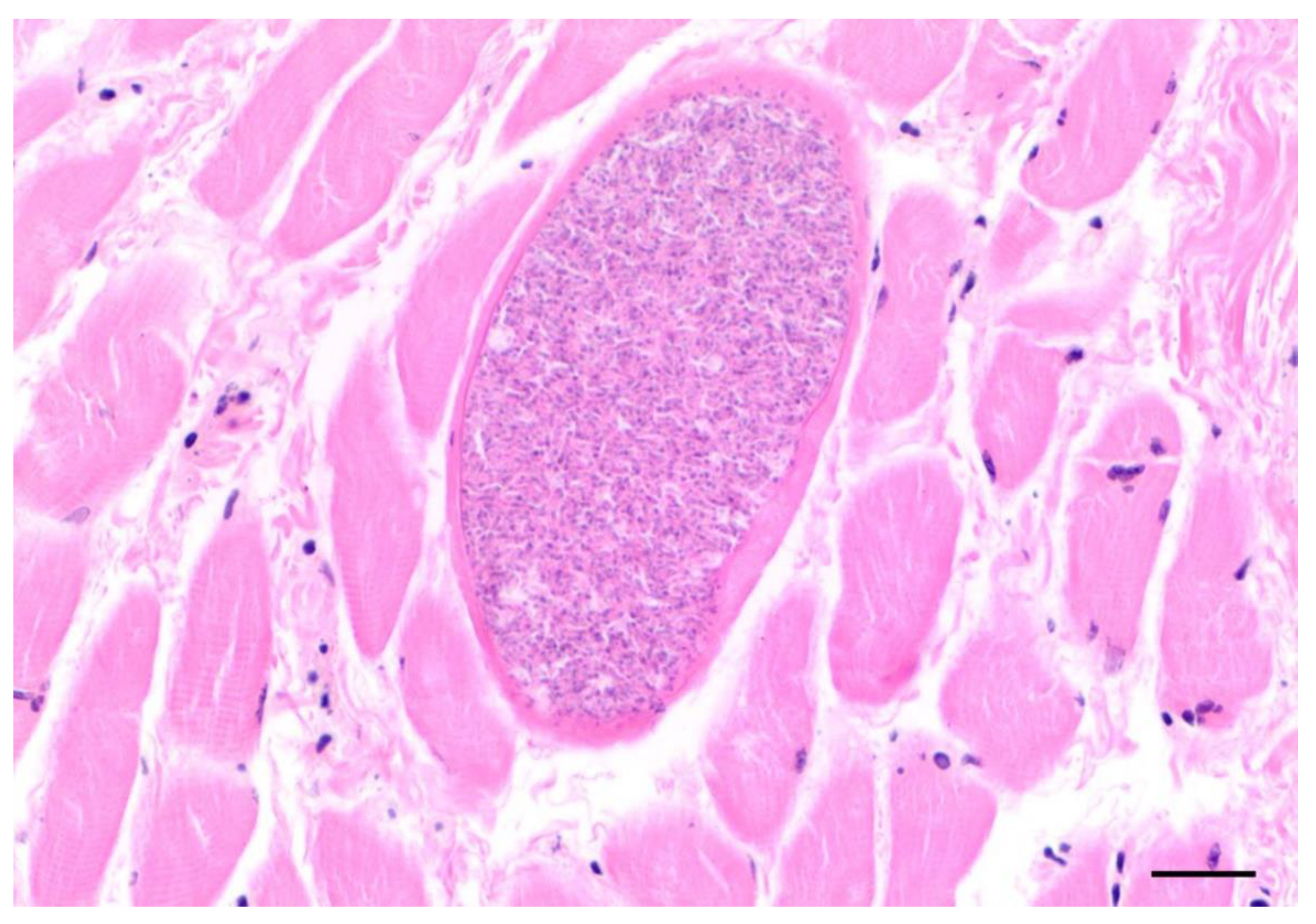

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age of Otters [Years] | Number of Otters | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Unknown Years | 2015–2018 | 2019 | 2020 | ||||||
| Male | Female | ND 1 | Male | Female | Male | Female | Male | Female | |
| <0.5 | / | / | / | / | / | 1 | / | 1 | / |
| 0.5–0.75 | / | / | / | 2 | 1 | 1 | 1 | 3 | 1 |
| 0.5–2.5 | / | / | 1 | 1 | 1 | 2 | 2 | 1 | 1 |
| 1.25–2.9 | / | / | / | 2 | 3 | 5 | 1 | 5 | 1 |
| 1.5–3.5 | / | / | / | / | / | / | 1 | 2 | 1 |
| 2.2–3.5 | / | / | / | 1 | / | 1 | 2 | 3 | 3 |
| 2.4–4.5 | / | 1 | / | / | 3 | / | 1 | / | / |
| 3.5–5.5 | 1 | / | / | / | 1 | / | 2 | 1 | 3 |
| 5.5–7.5 | / | / | / | / | / | / | / | 1 | / |
| Nutritional Status | Number of Otters | ||||
|---|---|---|---|---|---|
| Unknown Years | 2015–2018 | 2019 | 2020 | Total | |
| good | 5 | 8 | 17 | 22 | 52 |
| moderate | 1 | 7 | 3 | 3 | 14 |
| poor | / | 3 | 2 | 2 | 7 |
| ND 1 | 3 | 6 | 6 | 4 | 19 |
| Quantity | |||||
|---|---|---|---|---|---|
| Morphological Findings | Unknown Years | 2015–2018 | 2019 | 2020 | Total |
| Respiratory tract | |||||
| Pneumonia (purulent) | / | / | / | 1 | 1 |
| with intralesional foreign material | / | / | 1 | 1 | 2 |
| Pneumonia (nonpurulent) | / | 5 | 5 | 2 | 12 |
| Pneumonia (granulomatous) | 1 | 2 | 3 | 1 | 7 |
| with intralesional foreign material | 1 | / | 1 | / | 2 |
| Bronchopneumonia (nonpurulent) | 1 | / | 2 | / | 3 |
| with intralesinonal fungal spores | 1 | / | 1 | / | 2 |
| Bronchiolitis (nonpurulent) | / | / | 1 | / | 1 |
| Pleuritis (nonpurulent) | / | / | 1 | / | 1 |
| with bacteria pleura pulmonalis | / | / | 1 | / | 1 |
| Parenchymatous hemorrhages | / | / | 3 | 3 | 6 |
| Cardiovascular system | |||||
| Myocardial fibrosis with cell loss | 1 | / | / | / | 1 |
| Protozoal cysts myocard | / | 1 | 1 | / | 2 |
| Thoracic cavity | |||||
| Diaphragmatic serositis | / | / | 1 | / | 1 |
| Diaphragmatic edema | / | / | / | 1 | 1 |
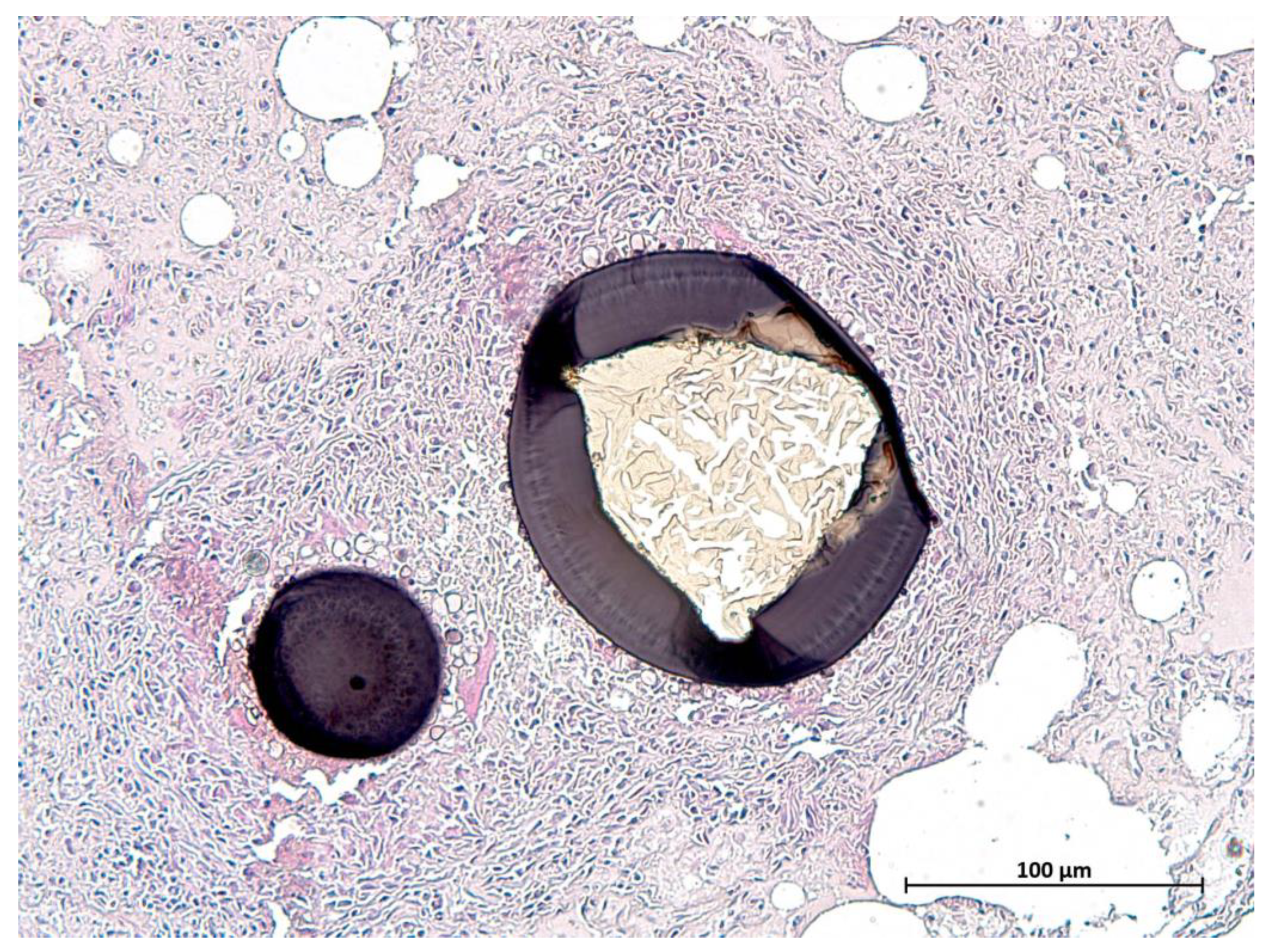
| Protozoal cysts diaphragm | / | 3 | 3 | 2 | 8 |
| Alimentary system | |||||
| Gastritis | / | / | 1 | / | 1 |
| Gastritis (ulcerative) | / | 1 | / | / | 1 |
| Erosions of gastric mucosa | / | 1 | 1 | / | 2 |
| Gastric edema (submucosal) | / | / | / | 1 | 1 |
| Enteritis (purulent) | / | / | / | 1 | 1 |
| Portal hepatitis | / | / | 1 | / | 1 |
| Portal fibrosis | 2 | / | / | / | 2 |
| Cholelithiasis | / | 1 | / | / | 1 |
| Protozoal cysts lingual musculature | / | 1 | 8 | 5 | 14 |
| Urinary and reproductive tract | |||||
| Nephrolithiasis | / | 2 | 1 | 1 | 4 |
| Urolithiasis | / | / | / | 1 | 1 |
| Cysts vas deferens | / | 2 | 4 | 12 | 18 |
| Placental scars | / | 1 | 2 | 1 | 4 |
| Skin and subcutis | |||||
| Epidermal hyperplasia | / | / | / | 1 | 1 |
| Dermal fibrosis | / | / | 1 | / | 1 |
| Subcutaneous bleeding | / | / | 1 | / | 1 |
| Bite wounds | / | 2 | 2 | 2 | 6 |
| Musculoskeletal system | |||||
| Hemorrhages periostal | / | / | / | 1 | 1 |
| Protozoal cysts skeletal muscle | / | 2 | 7 | 5 | 14 |
| Polydactylia | / | / | / | 1 | 1 |
| Central nervous system, eyes, and ears | |||||
| Leptomeningeal hemorrhages | / | / | 1 | 2 | 3 |
| Endophthalmitis | / | / | / | 1 | 1 |
| with retinal atrophy | / | / | / | 1 | 1 |
| Ectoparasites (ticks) | / | / | 2 | 5 | 7 |
| Hematopoetic and endocrine system | |||||
| Thyroid follicular cysts | / | 1 | 3 | 1 | 5 |
| Follicular hyperplasia (tonsil) | / | / | / | 3 | 3 |
| Follicular Hyperplasia (lymph node) | / | / | 3 | 3 | 6 |
| Follicular hyalinosis and depletion (lymph node) | 2 | 1 | 8 | 10 | 21 |
| Hemosiderosis lymph node | 1 | 1 | / | 3 | 5 |
| Anthracosis lymph node | 1 | / | / | 2 | 3 |
| Lymphoid depletion | / | / | / | 1 | 1 |
| Adrenalitis | 1 | 2 | 3 | 4 | 10 |
| Nodular hyperplasia adrenal gland | / | / | / | 2 | 2 |
| Fibrosis and mineralization adrenal gland | 1 | / | / | / | 1 |
| Periadrenal bleeding | / | / | 1 | / | 1 |
| Follicular hyperplasia (spleen) | / | / | 1 | / | 1 |
| Follicular hyalinosis and depletion (spleen) | / | 2 | 11 | 7 | 20 |
| Follicular hyalinosis (Peyer’s patches) | / | / | 1 | / | 1 |
| Bacteria | Liver | Spleen | Kidney | Lung | Pulm. Lymph Node | Intestine | Mes. Lymph Node | Brain | Repro | Sceletal Muscle | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Acinetobacter spp. | / | / | / | 2 | / | / | / | / | / | / | 2 |
| α-haem. Streptococci | 3 | 2 | 4 | 7 | 2 | 3 | 1 | 1 | / | 1 | 24 |
| Buttiauxella spp. | / | / | / | 1 | / | / | / | 2 | / | / | 3 |
| Candida famata | / | 1 | / | / | / | / | / | / | / | / | 1 |
| Carnobacterium divergens | 4 | 5 | 6 | 6 | 2 | 2 | 1 | / | / | 1 | 27 |
| Carnobacterium maltaromaticum | 2 | 1 | 1 | 1 | / | 1 | 2 | 1 | 1 | / | 10 |
| Clostridium colicanis | / | / | / | / | / | 1 | / | / | / | / | 1 |
| Clostridium perfringens | / | / | / | / | / | 3 | / | / | / | / | 3 |
| Enterococcus faecalis | / | / | / | 1 | / | 1 | / | / | / | 1 | 3 |
| Erwinia spp. | / | / | / | / | / | / | / | / | / | 1 | 1 |
| Escherichia coli | / | / | / | 2 | / | 1 | / | 1 | / | / | 4 |
| Ewingella americana | / | / | / | 1 | 1 | / | / | / | / | / | 2 |
| Yeasts (not specified further) | 1 | 1 | 1 | / | / | / | / | / | / | / | 3 |
| Kurtia spp. | 1 | / | 1 | / | / | / | / | / | / | / | 2 |
| Lactobacillus sakei | / | / | / | / | / | / | 1 | / | / | / | 1 |
| Macrococcus caseolyticus | / | / | / | 1 | / | / | / | / | / | / | 1 |
| Macrococcus spp. | / | / | 1 | 1 | / | / | 1 | / | / | / | 3 |
| Micrococcus luteus | / | / | / | 1 | / | / | / | / | / | / | 1 |
| Pantoea spp. | 1 | 2 | 1 | 3 | / | / | / | / | / | 1 | 8 |
| Paraclostridium bifermentans | / | / | / | / | / | 1 | / | / | / | / | 1 |
| Pseudomonas spp. | 3 | 2 | 5 | 7 | 3 | 3 | 2 | 1 | 1 | / | 27 |
| Psychrobacter spp. | 2 | 2 | 1 | 2 | / | / | / | / | / | / | 7 |
| Rhanella aquatilis | / | / | / | 2 | / | / | / | / | / | / | 2 |
| Rhanella spp. | / | / | 1 | 1 | / | / | / | 1 | / | / | 3 |
| Serratia fonticola | / | / | / | 3 | 1 | / | / | 1 | / | / | 5 |
| Serratia spp. | / | / | / | / | / | / | / | 1 | / | / | 1 |
| Staphylococcus equorum | / | / | / | 1 | / | / | / | / | / | / | 1 |
| Staphylococcus lutrae | / | / | / | / | / | 1 | / | / | / | / | 1 |
| Staphylococcus saprophyticus | / | / | / | 1 | / | / | / | / | / | / | 1 |
| Staphylococcus sciuri | / | / | / | 1 | / | / | / | / | / | / | 1 |
| Stenotrophomonas maltophilia | / | / | / | 1 | / | / | / | / | / | / | 1 |
| Stenotrophomonas spp. | / | / | / | / | / | / | / | 1 | / | / | 1 |
| Streptococcus canis | / | / | / | 1 | / | / | / | / | / | / | 1 |
| Streptococcus dysgalactiae | / | / | / | / | 1 | / | / | / | / | / | 1 |
| Vagococcus lutrae | / | / | / | / | / | 1 | 1 | / | / | / | 2 |
| Yarrowia lipolytica | / | / | / | / | / | / | / | / | 1 | / | 1 |
| γ-haem. Streptococci | / | / | 1 | 6 | 1 | 1 | / | 2 | / | / | 11 |
| Total | 17 | 16 | 23 | 53 | 11 | 19 | 9 | 12 | 3 | 5 |
| Years of Finding | Cause of Death | |||
|---|---|---|---|---|
| Trauma | Starvation | Disease | Unclear | |
| unknown years | 3 | / | / | 3 |
| 2015–2018 | 17 | / | / | 10 |
| 2019 | 26 | 1 | 1 | / |
| 2020 | 28 | 2 | / | 1 |
| Total | 74 | 3 | 1 | 14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rohner, S.; Wohlsein, P.; Prenger-Berninghoff, E.; Ewers, C.; Waindok, P.; Strube, C.; Baechlein, C.; Becher, P.; Wilmes, D.; Rickerts, V.; et al. Pathological Findings in Eurasian Otters (Lutra lutra) Found Dead between 2015−2020 in Schleswig-Holstein, Germany. Animals 2022, 12, 59. https://doi.org/10.3390/ani12010059
Rohner S, Wohlsein P, Prenger-Berninghoff E, Ewers C, Waindok P, Strube C, Baechlein C, Becher P, Wilmes D, Rickerts V, et al. Pathological Findings in Eurasian Otters (Lutra lutra) Found Dead between 2015−2020 in Schleswig-Holstein, Germany. Animals. 2022; 12(1):59. https://doi.org/10.3390/ani12010059
Chicago/Turabian StyleRohner, Simon, Peter Wohlsein, Ellen Prenger-Berninghoff, Christa Ewers, Patrick Waindok, Christina Strube, Christine Baechlein, Paul Becher, Dunja Wilmes, Volker Rickerts, and et al. 2022. "Pathological Findings in Eurasian Otters (Lutra lutra) Found Dead between 2015−2020 in Schleswig-Holstein, Germany" Animals 12, no. 1: 59. https://doi.org/10.3390/ani12010059
APA StyleRohner, S., Wohlsein, P., Prenger-Berninghoff, E., Ewers, C., Waindok, P., Strube, C., Baechlein, C., Becher, P., Wilmes, D., Rickerts, V., & Siebert, U. (2022). Pathological Findings in Eurasian Otters (Lutra lutra) Found Dead between 2015−2020 in Schleswig-Holstein, Germany. Animals, 12(1), 59. https://doi.org/10.3390/ani12010059