
Feather Corticosterone Measurements and Behavioral Observations in the Great White Pelican (Pelecanus onocrotalus) Living under Different Flight Restraint Conditions in German Zoos

 , , , , ,
, , , , ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Study Design and List of the Participating Zoos
2.3. Behavioral Observation
2.4. Feather Collection
2.5. Corticosterone Extraction and Corticosterone Concentration Measurements
2.6. Statistics
3. Results
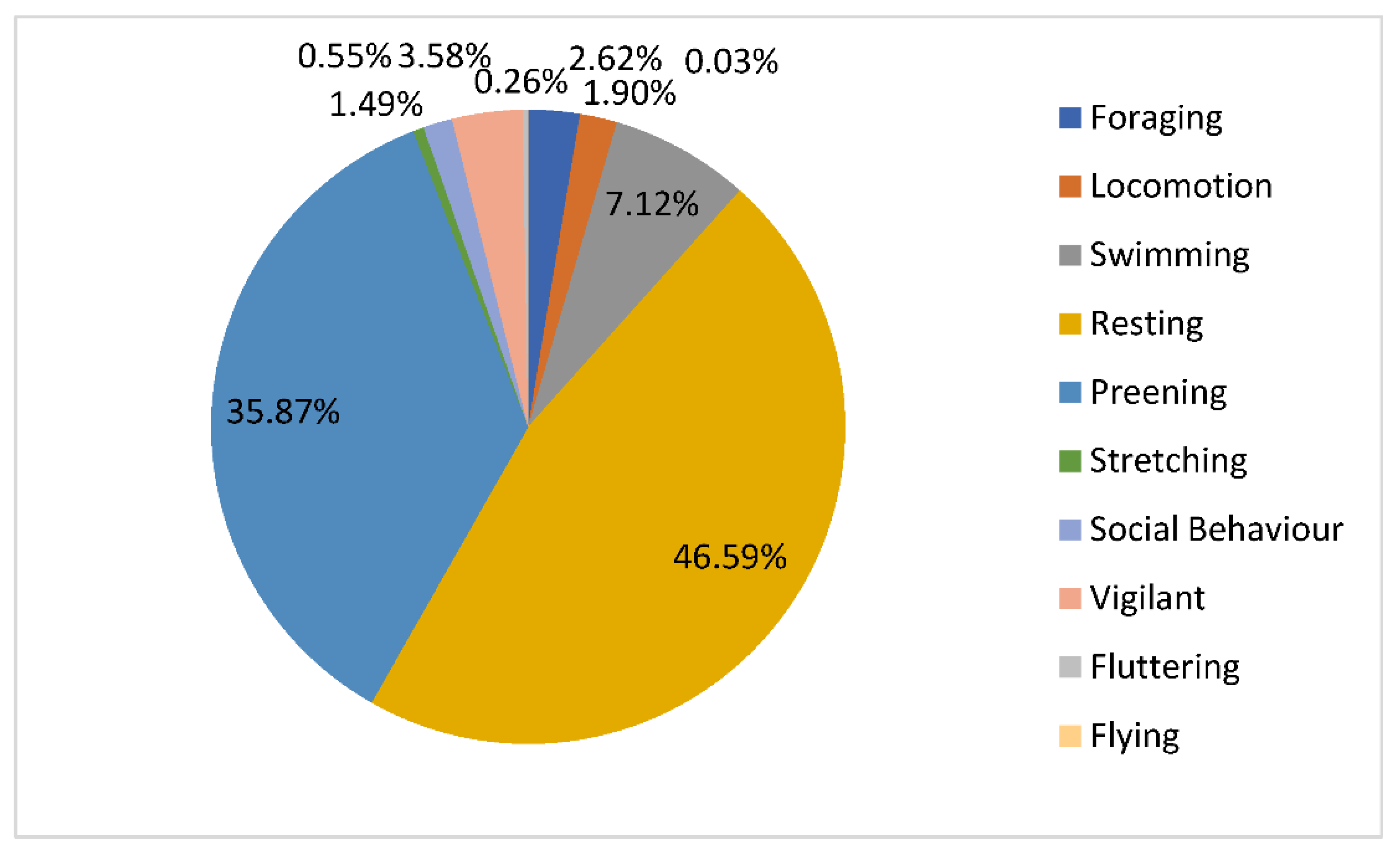
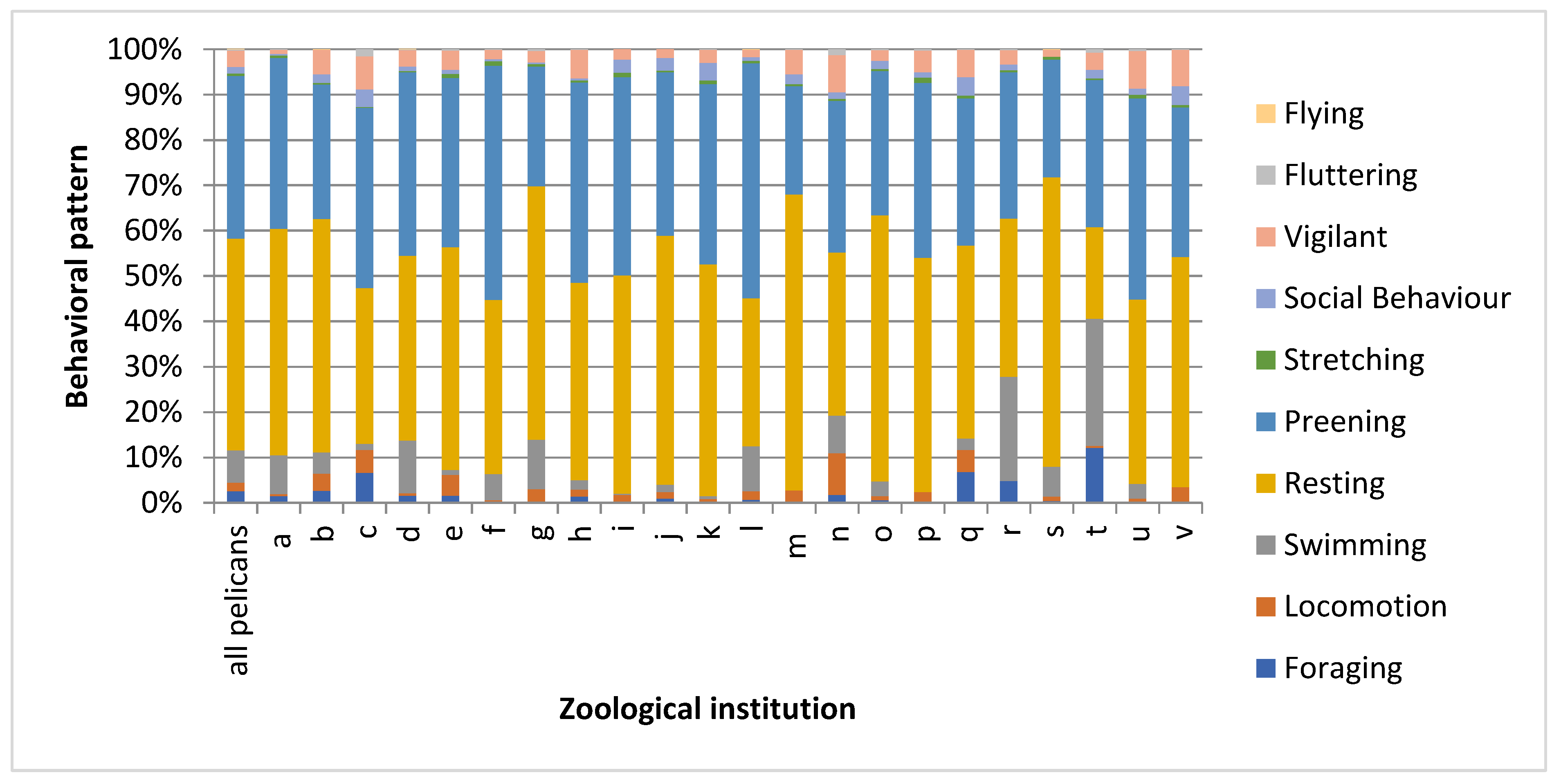
3.1. Behavioral Observations
3.2. Feather Corticosterone Concentrations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Council Directive 1999/22/EC of 29 March 1999 Relating to the Keeping of Wild Animals in Zoos. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=uriserv:OJ.L_.1999.094.01.0024.01.ENG (accessed on 7 July 2021).
- Ward, S.J.; Sherwen, S.; Clark, F.E. Advances in Applied Zoo Animal Welfare Science. J. Appl. Anim. Welf. Sci. 2018, 21, 23–33. [Google Scholar] [CrossRef]
- Binding, S.; Farmer, H.; Krusin, L.; Cronin, K. Status of Animal Welfare Research in Zoos and Aquariums: Where Are We, Where to Next? J. Zoo Aquar. Res. 2020, 8, 166–174. [Google Scholar] [CrossRef]
- Dollinger, P.; Pagel, T.; Baumgartner, K.; Encke, D.; Engel, H.; Filz, A. Flugunfähigmachen von Vögeln–Für und Wider. Zool. Gart. 2013, 82, 293–339. [Google Scholar] [CrossRef]
- Peng, S.J.-L.; Chang, F.-C.; Sheng-Ting, J.; Fei, A. Welfare Assessment of Flight-Restrained Captive Birds: Effects of Inhibition of Locomotion. Thai J. Vet. Med. 2013, 43, 235–241. [Google Scholar]
- Maisack, C.; Schmidt, T. Zum Flugunfähigmachen von Vögeln in Zoos und privaten Geflügelhaltungen. Nat. Recht 2017, 39, 734–741. [Google Scholar] [CrossRef]
- Tyson For an End to Pinioning: The Case Against the Legal Mutilation of Birds in Captivity. J. Anim. Ethic 2014, 4, 1–4. [CrossRef]
- Hesterman, H.; Gregory, N.G.; Boardman, W.S.J. Deflighting Procedures and Their Welfare Implications in Captive Birds. Anim. Welf. 2001, 10, 405–419. [Google Scholar]
- Reese, L.; Ladwig-Wiegard, M.; Von Fersen, L.; Haase, G.; Will, H.; Merle, R.; Encke, D.; Maegdefrau, H.; Baumgartner, K.; Thöne-Reineke, C. Deflighting zoo birds and its welfare considerations. Anim. Welf. 2020, 29, 69–80. [Google Scholar] [CrossRef]
- Tierschutzgesetz in der Fassung der Bekanntmachung vom 18. Mai 2006 (BGBl. I S. 1206, 1313), das Zuletzt Durch Artikel 1 des Gesetzes vom 18. Juni 2021 (BGBl. I S. 1828) Geändert Worden ist. Available online: https://www.gesetze-im-internet.de/tierschg/BJNR012770972.html (accessed on 7 July 2021).
- Djurskyddsmyndighetens Föreskrifter om Djurhållning i Djurparker 2004 Djurskyddsmyndigheten L: Sweden. Available online: https://www.jordbruksverket.se/download/18.26424bf71212ecc74b08000877/1370040445226/DFS_2004-19.pdf (accessed on 7 July 2021).
- Hosey, G.; Melfi, V.; Pankhurst, S. Zoo Animals: Behaviour, Management, and Welfare; OUP Oxford: Oxford, UK, 2013; ISBN 978-0-19-969352-8. [Google Scholar]
- Häffelin, K.; Lindenwald, R.; Kaufmann, F.; Döhring, S.; Spindler, B.; Preisinger, R.; Rautenschlein, S.; Kemper, N.; Andersson, R. Corticosterone in feathers of laying hens: An assay validation for evidence-based assessment of animal welfare. Poult. Sci. 2020, 99, 4685–4694. [Google Scholar] [CrossRef] [PubMed]
- Johns, D.W.; Marchant, T.A.; Fairhurst, G.D.; Speakman, J.R.; Clark, R.G. Biomarker of burden: Feather corticosterone reflects energetic expenditure and allostatic overload in captive waterfowl. Funct. Ecol. 2018, 32, 345–357. [Google Scholar] [CrossRef]
- Paul-Murphy, J. Foundations in Avian Welfare. In Current Therapy in Avian Medicine and Surgery; Speer, B.L., Ed.; Elsevier: St. Louis, MO, USA, 2016. [Google Scholar]
- Palme, R.; Rettenbacher, S.; Touma, C.; El-Bahr, S.M.; Möstl, E. Stress Hormones in Mammals and Birds: Comparative Aspects Regarding Metabolism, Excretion, and Noninvasive Measurement in Fecal Samples. Ann. N. Y. Acad. Sci. 2005, 1040, 162–171. [Google Scholar] [CrossRef]
- Harris, C.M.; Madliger, C.L.; Love, O.P. Temporal overlap and repeatability of feather corticosterone levels: Practical considerations for use as a biomarker. Conserv. Physiol. 2016, 4, cow051. [Google Scholar] [CrossRef]
- Bortolotti, G.R.; Marchant, T.A.; Blas, J.; German, T. Corticosterone in feathers is a long-term, integrated measure of avian stress physiology. Funct. Ecol. 2008, 22, 494–500. [Google Scholar] [CrossRef]
- Monclús, L.; Lopez-Bejar, M.; De La Puente, J.; Covaci, A.; Jaspers, V.L. First evaluation of the use of down feathers for monitoring persistent organic pollutants and organophosphate ester flame retardants: A pilot study using nestlings of the endangered cinereous vulture (Aegypius monachus). Environ. Pollut. 2018, 238, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Glucs, Z.E.; Smith, D.R.; Tubbs, C.W.; Scherbinski, J.J.; Welch, A.; Burnett, J.; Clark, M.; Eng, C.; Finkelstein, M.E. Glucocorticoid measurement in plasma, urates, and feathers from California condors (Gymnogyps californianus) in response to a human-induced stressor. PLoS ONE 2018, 13, e0205565. [Google Scholar] [CrossRef] [PubMed]
- Carbajal, A.; Tallo-Parra, O.; Sabes-Alsina, M.; Mular, I.; Lopez-Bejar, M. Feather corticosterone evaluated by ELISA in broilers: A potential tool to evaluate broiler welfare. Poult. Sci. 2014, 93, 2884–2886. [Google Scholar] [CrossRef] [PubMed]
- Reese, L.; Baumgartner, K.; Thöne-Reineke, C.; Von Fersen, L.; Merle, R.; Ladwig-Wiegard, M.; Will, H.; Haase, G.; Tallo-Parra, O.; Carbajal, A.; et al. Feather Corticosterone Measurements of Greater Flamingos Living under Different Forms of Flight Restraint. Animals 2020, 10, 605. [Google Scholar] [CrossRef]
- Crivelli, A.J.; Schreiber, R.W. Status of the Pelecanidae. Biol. Conserv. 1984, 30, 147–156. [Google Scholar] [CrossRef]
- Klausen, B. A mixed-species exhibit for African water birds (including pelicans, flamingos, spoonbills and storks) at Odense Zoo, Denmark: Breeding success, animal welfare and education. Int. Zoo Yearb. 2014, 48, 61–68. [Google Scholar] [CrossRef]
- Gokula, V. An ethogram of Spot-billed Pelican (Pelecanus philippensis). Chin. Birds 2012, 2, 183–192. [Google Scholar] [CrossRef][Green Version]
- Verband Deutscher Zoodirektoren; International Union of Directors of Zoological Gardens; World Zoo Organization. Der Zoologische Garten: Zeitschrift Für die Gesamte Tiergärtnerei; Offizielles Organ des Verbandes der Zoologischen Gärten-VdZ; Organ of the World Association of Zoos & Aquariums-WAZA. 1949. Available online: https://www.biodiversitylibrary.org/creator/233898/author?bpg1&ppg=1&psize=250#/titles (accessed on 7 July 2021).
- Harrison, C.J.O. Allopreening as Agonistic Behaviour. Behaviour 1965, 24, 161–208. [Google Scholar] [CrossRef]
- Monclús, L.; Carbajal, A.; Tallo-Parra, O.; Sabés-Alsina, M.; Darwich, L.; López, R.M.; Lopez-Bejar, M. Relationship between feather corticosterone and subsequent health status and survival in wild Eurasian Sparrowhawk. J. Ornithol. 2017, 158, 773–783. [Google Scholar] [CrossRef]
- del Hoyo, J.; Elliott, A.; Sargatal, J. (Eds.) Handbook of the Birds of the World; Lynx Edicions: Barcelona, Spain, 1992; Volume 1. [Google Scholar]
- Brown, L.H.; Urban, E.K. The breeding biology of the great white pelican Pelecanus onocrotalus roseus at Lake Shala, Ethiopia. Ibis 2008, 111, 199–237. [Google Scholar] [CrossRef]
- Schreiber, R.W.; Schreiber, E.A.; Anderson, D.W.; Bradley, D.W. Plumages and Molts of Brown Pelicans; Natural History Museum of Los Angeles County: Los Angeles, CA, USA, 1989. [Google Scholar]
- Monclús, L.; Tallo-Parra, O.; Carbajal, A.; Quevedo, M.A.; Lopez-Bejar, M. Feather corticosterone in Northern Bald Ibis Geronticus eremita: A stable matrix over time able to predict reproductive success. J. Ornithol. 2020, 161, 557–567. [Google Scholar] [CrossRef]
- Adámková, M.; Bílková, Z.; Tomášek, O.; Šimek, Z.; Albrecht, T. Feather steroid hormone concentrations in relation to age, sex, and molting time in a long-distance migratory passerine. Ecol. Evol. 2019, 9, 9018–9026. [Google Scholar] [CrossRef] [PubMed]
- Marinov, M.; Tamiris, P.; Dorosencu, A.; Nichersu, I.; Vasile, A.; Trifanov, C.; Bozagievici, R.; Tošić, K.; Botond, J. Monitoring the Great White Pelican (Pelecanus Onocrotalus Linnaeus, 1758) Breeding Population Using Drones in 2016-the Danube Delta (Romania). Sci. Ann. Danube Delta Inst. 2016, 22, 41–52. [Google Scholar]
- Breed, M. 1973 The Many Eyes Hypothesis. In Conceptual Breakthroughs in Ethology and Animal Behavior; Academic Press: Cambridge, MA, USA, 2017; pp. 121–122. ISBN 978-0-12-809265-1. [Google Scholar]
- Rubenstein, D.I.; Wrangham, R.W. Ecological Aspects of Social Evolution: Birds and Mammals; University Press: Princeton, NJ, USA, 1987; ISBN 978-0-691-61016-0. [Google Scholar]
- Kappeler, P. Verhaltensbiologie; Springer: Berlin/Heidelberg, Germany, 2017; ISBN 978-3-662-53144-0. [Google Scholar]
- Voit, M.; Merle, R.; Thöne-Reineke, C.; Baumgartner, K.; Von Fersen, L.; Reese, L.; Ladwig-Wiegard, M.; Will, H.; Tallo-Parra, O.; Carbajal, A.; et al. Validation of an Alternative Feather Sampling Method to Measure Corticosterone. Animals 2020, 10, 2054. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Zoo | Total Number of Observed Animals | Feather Samples for CORTf | Flight Status Airworthy | Flight Status Irreversibly Deflighted | Flight Status Reversibly Deflighted |
|---|---|---|---|---|---|
| a | 18 | 10 | 10 | ||
| b | 10 | 10 | 10 | ||
| c | 3 | 3 | 3 | ||
| d | 16 | 16 | 9 | 7 | |
| e | 14 | 13 | 13 | ||
| f | 5 | 5 | 5 | ||
| g | 25 | 10 | 10 | ||
| h | 16 | 11 | 11 | ||
| i | 8 | 8 | 8 | ||
| j | 4 | 4 | 4 | ||
| k | 10 | 10 | 6 | 4 | |
| l | 9 | 9 | 1 | 8 | |
| m | 4 | 4 | 4 | ||
| n | 4 | 4 | 4 | ||
| o | 16 | 12 | 12 | ||
| p | 10 | 10 | 6 | 4 | |
| q | 3 | 3 | 3 | ||
| r | 8 | 8 | 4 | 4 | |
| s | 9 | 9 | 7 | ||
| t | 10 | 10 | 2 | 6 | 4 |
| u | 5 | 5 | 3 | 2 | |
| v | 8 | 8 | 8 | ||
| Total | 215 | 182 | 7 | 134 | 41 |
| Behavior Pattern | |
|---|---|
| Locomotion | Pelicans shift their weight from 1 side to another, 1 foot is set on the ground by the other in sequence. They roll over from the heel to the toes. The speed depends on the circumstances; they can also run. The wings are sometimes opened for balance. |
| Stretching | There are 2 types of stretching: the pelicans stretch their head horizontally while the wings are pulled backwards, or the pelicans stretch one foot horizontally backward and the equilateral wing is pulled backward. |
| Preening | There are different types of preening, but commonly, “preening involves the contact between the bill and feathers” [25]. The feathers of the whole body can be erected and laid straight back. The wings are opened during sunbathing to dry their feathers. Scratching is also a type of preening. |
| Swimming | Pelicans swim by paddling with their feet in the water. |
| Resting | They stand on their feet on the ground, or they lay down and their eyes can be closed or be open. The head is placed vertically on their back. In this position they often put their beak in the feathers on the back. |
| Foraging | Pelicans try to catch fish by diving with their head or with only their beak under water while opening their beak. |
| Social behavior | Social behavior is described as interactions between 2 or more pelicans. It can be antagonistic behavior or, for example, allopreening, when “it preens the plumage of another bird” [27]. |
| Vigilant | If pelicans are alert, the neck will be stretched upward and the eyes will be open. Pelicans stand or sit in this position. |
| Fluttering | Fluttering is expressed by walking, running, or standing combined with moving of the wings. |
| Flying | If there is no connection to the ground, the pelicans are flying by moving their wings up and down. |
| n | Mean | Minimum | Maximum | |
|---|---|---|---|---|
| Status | ||||
| Airworthy | 7 | 1.53 | 1.24 | 1.95 |
| Extirpated | 6 | 1.46 | 1.05 | 1.73 |
| Pinioned | 128 | 1.53 | 0.89 | 2.45 |
| Feather-clipped | 41 | 1.67 | 1.17 | 2.25 |
| Group size | ||||
| 1–5 pelicans | 28 | 1.71 | 1.18 | 2.45 |
| 6–10 pelicans | 82 | 1.57 | 1.07 | 2.09 |
| >10 pelicans | 72 | 1.48 | 0.89 | 2.29 |
| Sex | ||||
| Female | 98 | 1.58 | 0.89 | 2.45 |
| Male | 84 | 1.54 | 1.12 | 2.29 |
| Age | ||||
| <1 year | 3 | 1.72 | 1.50 | 2.02 |
| 1–3 years | 4 | 1.76 | 1.43 | 2.09 |
| >3 years | 175 | 1.55 | 0.89 | 2.45 |
| Parameter | p-Value Univariable | Effect Size | p-Value Multivariable | Effect Size |
|---|---|---|---|---|
| Foraging | 0.215 | 0.233 | ||
| Walking | 0.081 | 0.283 | ||
| Swimming | 0.611 | 0.103 | ||
| Resting | 0.241 | 0.222 | ||
| Preening | 0.757 | 0.058 | ||
| Stretching | 0.688 | 0.079 | 0.260 | 0.197 |
| Social behavior | 0.296 | 0.188 | ||
| Vigilant | 0.471 | 0.126 | ||
| Fluttering | 0.030 | 0.303 | 0.043 | 0.282 |
| Flying | 0.284 | 0.218 | ||
| group size | 0.154 | 001 | 0.083 | 0.1 |
| Age | 0.821 | 0.047/0.038 | 0.885 | 0.047/0.038 |
| Sex | 0.609 | 0.019 | ||
| Status | 0.946 | 0.021/0.020 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haase, G.; Baumgartner, K.; von Fersen, L.; Merle, R.; Wiegard, M.; Will, H.; Reese, L.; Tallo-Parra, O.; Carbajal, A.; Lopez-Bejar, M.; et al. Feather Corticosterone Measurements and Behavioral Observations in the Great White Pelican (Pelecanus onocrotalus) Living under Different Flight Restraint Conditions in German Zoos. Animals 2021, 11, 2522. https://doi.org/10.3390/ani11092522
Haase G, Baumgartner K, von Fersen L, Merle R, Wiegard M, Will H, Reese L, Tallo-Parra O, Carbajal A, Lopez-Bejar M, et al. Feather Corticosterone Measurements and Behavioral Observations in the Great White Pelican (Pelecanus onocrotalus) Living under Different Flight Restraint Conditions in German Zoos. Animals. 2021; 11(9):2522. https://doi.org/10.3390/ani11092522
Chicago/Turabian StyleHaase, Gudrun, Katrin Baumgartner, Lorenzo von Fersen, Roswitha Merle, Mechthild Wiegard, Hermann Will, Lukas Reese, Oriol Tallo-Parra, Annais Carbajal, Manel Lopez-Bejar, and et al. 2021. "Feather Corticosterone Measurements and Behavioral Observations in the Great White Pelican (Pelecanus onocrotalus) Living under Different Flight Restraint Conditions in German Zoos" Animals 11, no. 9: 2522. https://doi.org/10.3390/ani11092522
APA StyleHaase, G., Baumgartner, K., von Fersen, L., Merle, R., Wiegard, M., Will, H., Reese, L., Tallo-Parra, O., Carbajal, A., Lopez-Bejar, M., & Thöne-Reineke, C. (2021). Feather Corticosterone Measurements and Behavioral Observations in the Great White Pelican (Pelecanus onocrotalus) Living under Different Flight Restraint Conditions in German Zoos. Animals, 11(9), 2522. https://doi.org/10.3390/ani11092522