
Insight View on the Role of in Ovo Feeding of Clenbuterol on Hatched Chicks: Hatchability, Growth Efficiency, Serum Metabolic Profile, Muscle, and Lipid-Related Markers

 ,
,  , , , and
, , , and 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Growth Efficiency and Yield of Carcass
2.3. Blood and Tissue Sampling
2.4. Hematological Analysis
2.5. Blood Biochemical Analysis
2.6. Antioxidant Activity in Breast Muscles
2.7. Immunity Markers
2.8. Amino Acid and Muscle Fatty Acid Profiles
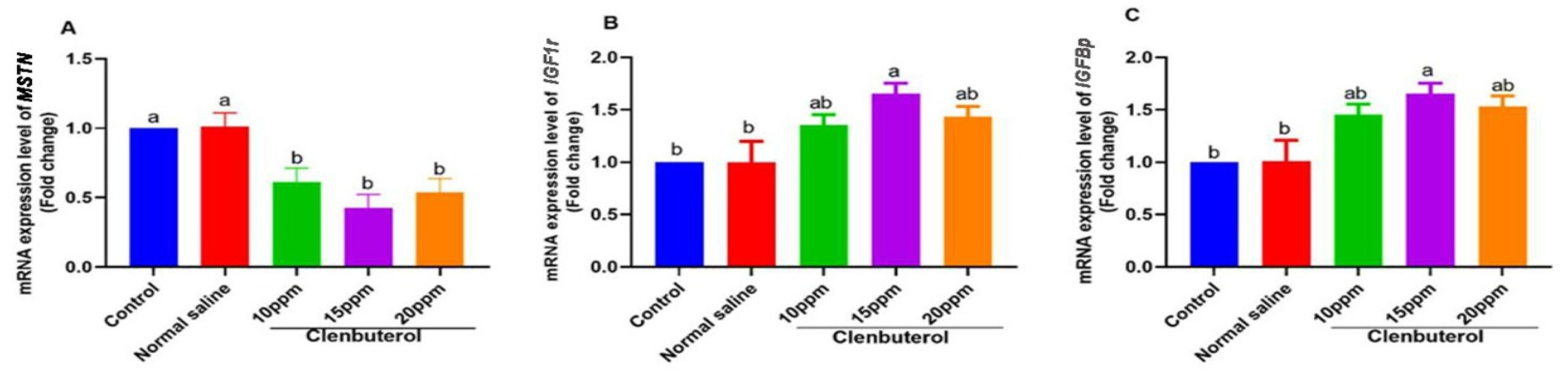
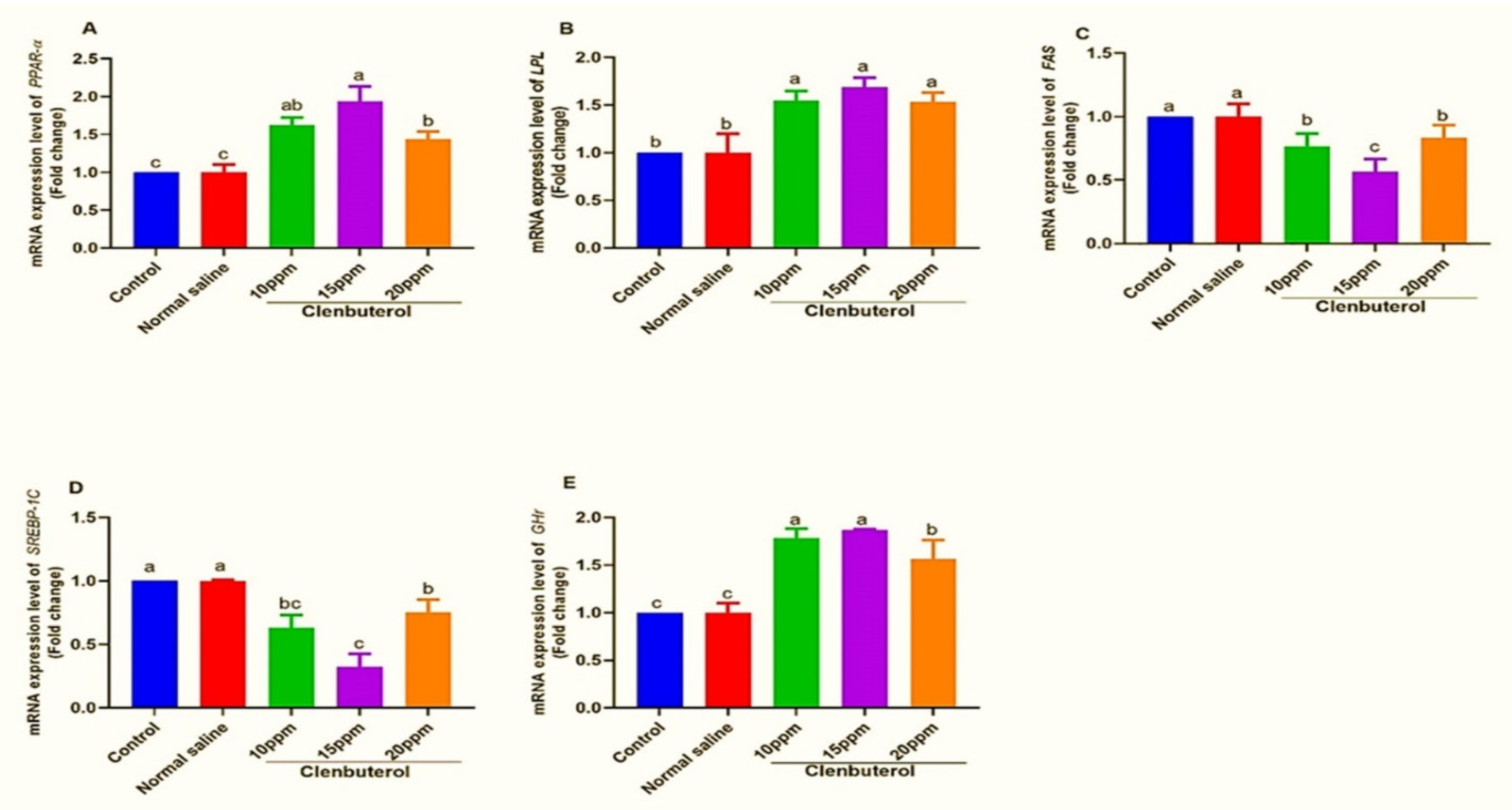
2.9. Gene Expression Analysis
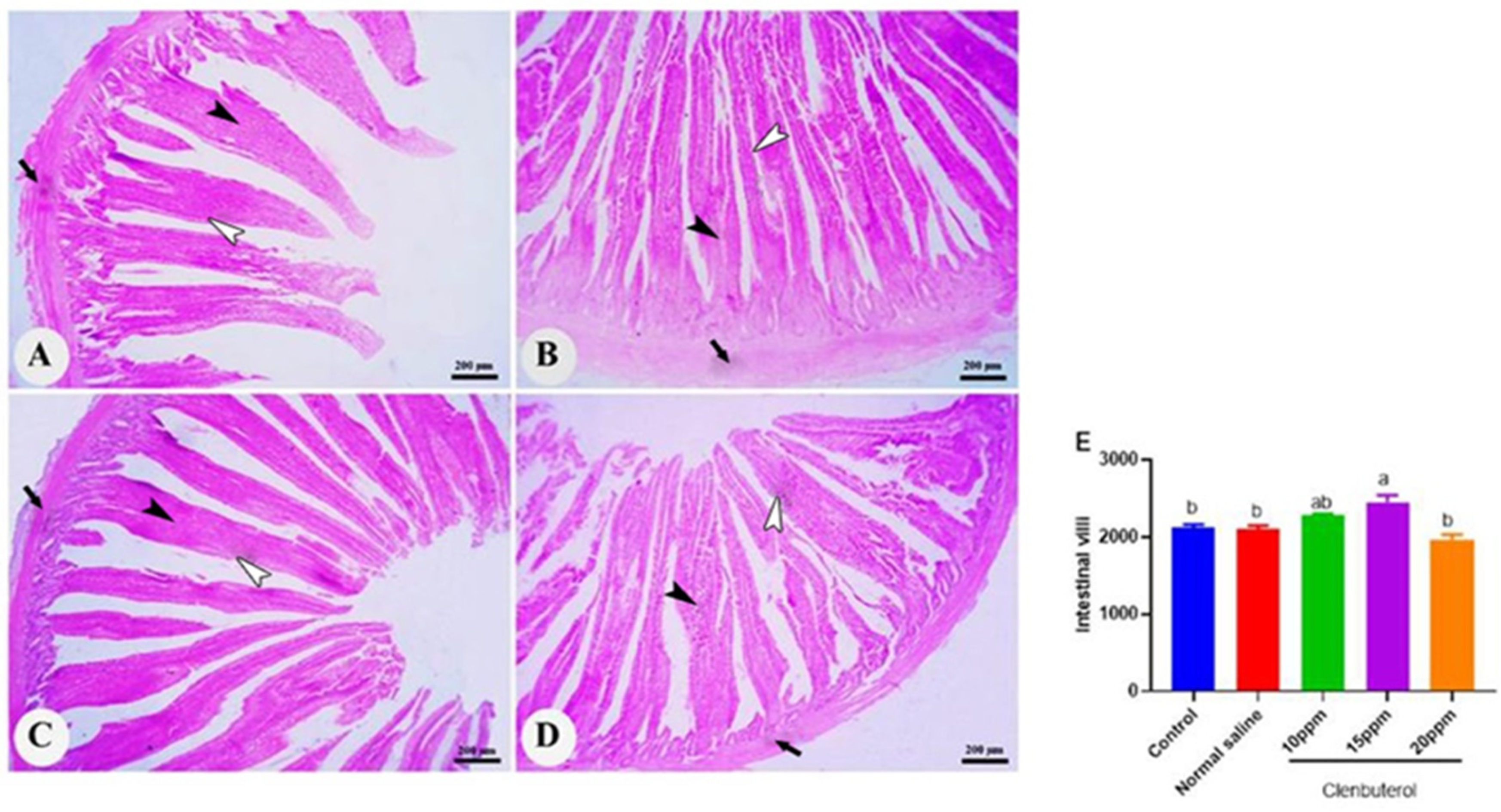
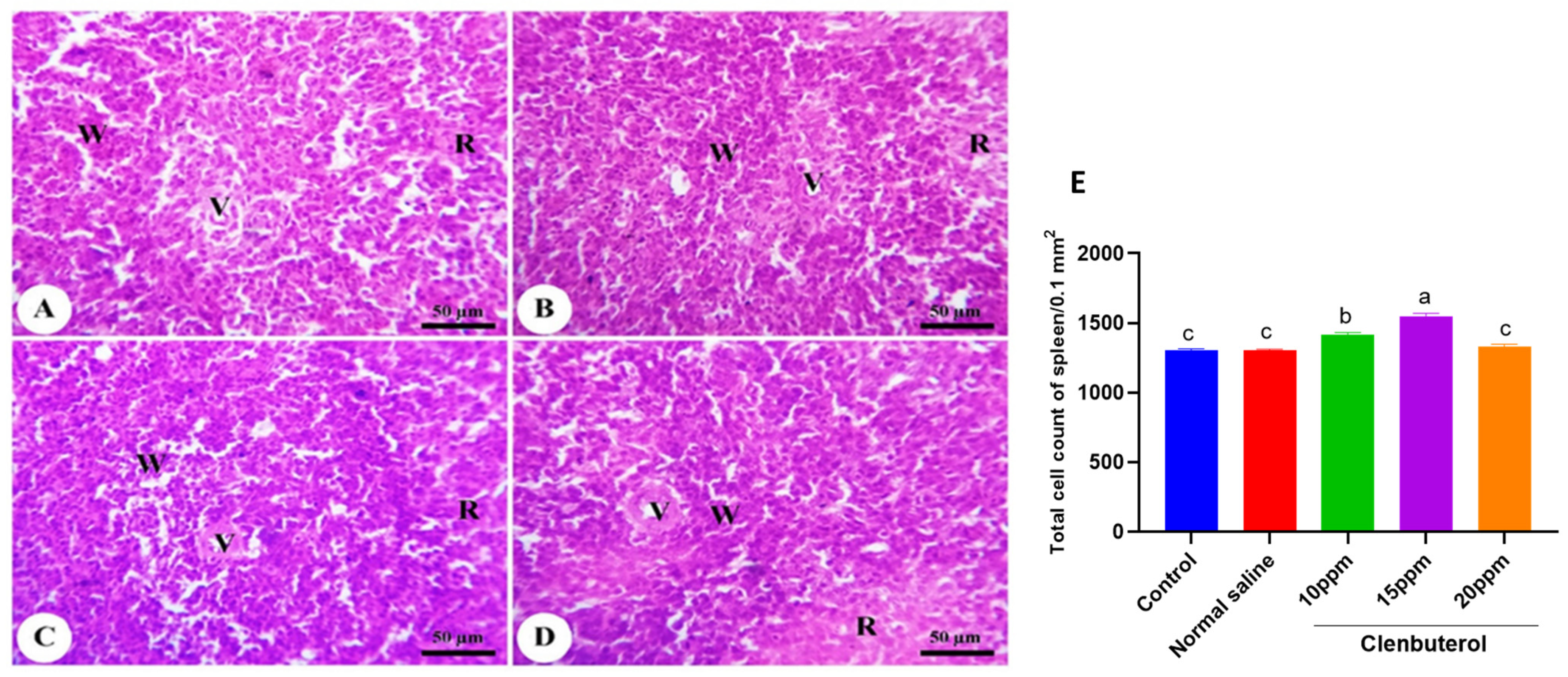
2.10. Histomorphometric Examination
2.11. Data Analysis
3. Results
3.1. Growth Performance Analysis and Carcass Traits
3.2. Blood Biochemical and Hematological Markers
3.3. Immunity and Antioxidant Activity
3.4. Muscle Fatty and Amino Acid Profiles
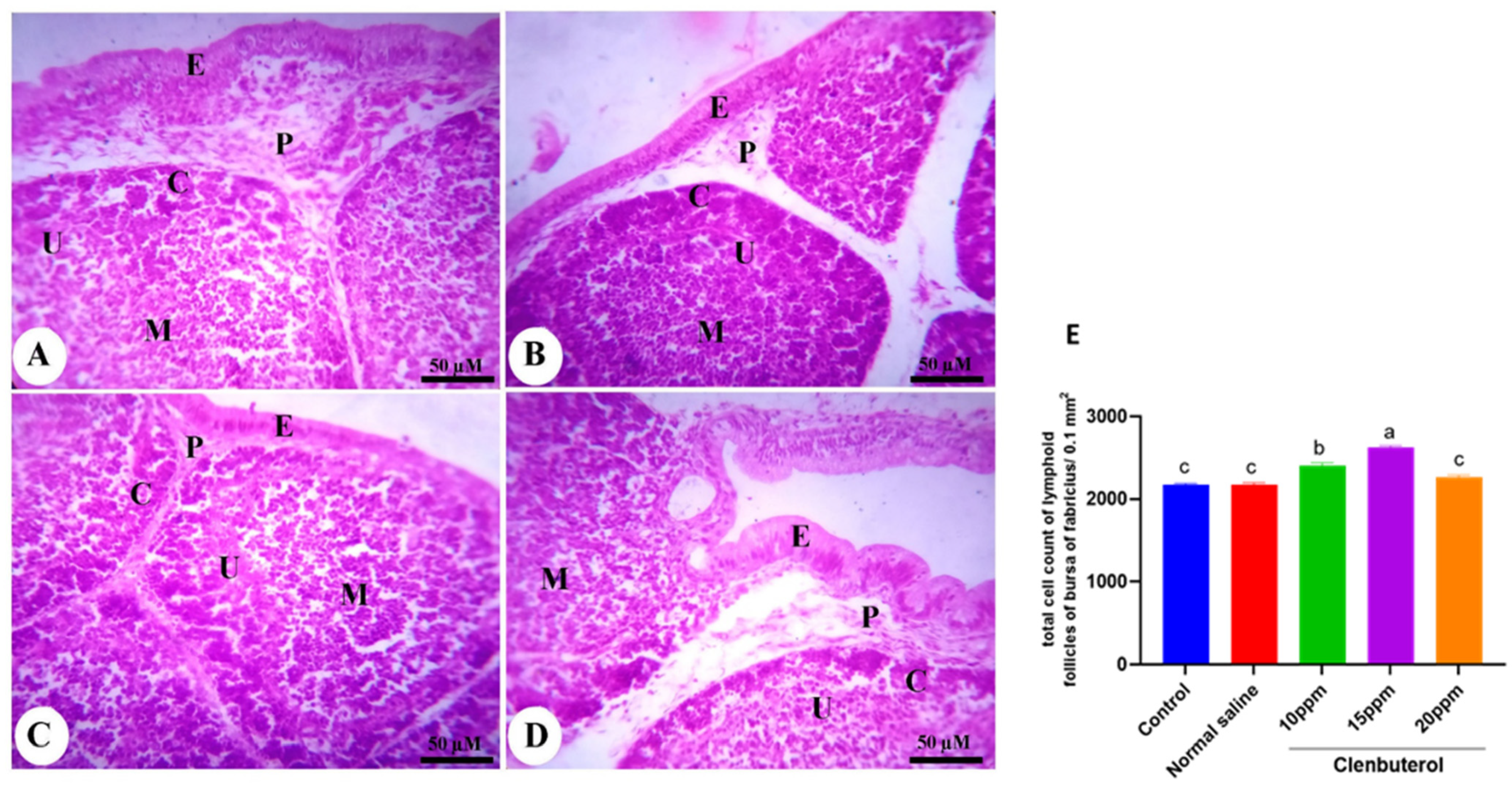
3.5. Histomorphometry of the Duodenum, Spleen and Bursa of Fabricius
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scanes, C. The global importance of poultry. Poult. Sci. 2007, 86, 1057. [Google Scholar] [CrossRef] [PubMed]
- Abd El, A.E.-M.E.; El-Wardany, I.; Abu-Taleb, A.M.; Wakwak, M.M.; Ebeid, T.A.; Saleh, A.A. Assessment of in ovo administration of Bifidobacterium bifidum and Bifidobacterium longum on performance, ileal histomorphometry, blood hematological, and biochemical parameters of broilers. Probiotics Antimicrob. Proteins 2020, 12, 439–450. [Google Scholar]
- Bhanja, S.; Mandal, A. Effect of in ovo injection of critical amino acids on pre-and post-hatch growth, immunocompetence and development of digestive organs in broiler chickens. Asian Australas. J. Anim. Sci. 2005, 18, 524–531. [Google Scholar] [CrossRef]
- Ferket, P. In ovo feeding and the promise of perinatal nutrition. In Proceedings of the Alltech International Nutrition Symposium, Lexington, KY, USA, 22–25 May 2011. [Google Scholar]
- Hassan, A.M. Effect of IN-OVO injection with Nano Iron -Particles on Physiological Responses and Performance of Broiler Chickens under Saini Conditions. Int. J. Environ. Agric. Biotechnol. 2018, 3, 855–863. [Google Scholar] [CrossRef]
- Sogunle, O.; Elangovan, A.; David, C.; Ghosh, J.; Awachat, V. Response of broiler chicken to in ovo administration of inorganic salts of Zinc, Selenium and Copper or their combination. Slovak J. Anim. Sci. 2018, 51, 8–19. [Google Scholar]
- Dawood, M.A.; Koshio, S.; Ishikawa, M.; Yokoyama, S. Immune responses and stress resistance in red sea bream, Pagrus major, after oral administration of heat-killed Lactobacillus plantarum and vitamin C. Fish Shellfish Immunol. 2016, 54, 266–275. [Google Scholar] [CrossRef]
- Burniston, J.G.; McLean, L.; Beynon, R.J.; Goldspink, D.F. Anabolic effects of a non-myotoxic dose of the β2-adrenergic receptor agonist clenbuterol on rat plantaris muscle. Muscle Nerve 2007, 35, 217–223. [Google Scholar] [CrossRef]
- Zhou, L.; Li, Y.; Nie, T.; Feng, S.; Yuan, J.; Chen, H.; Yang, Z. Clenbuterol inhibits SREBP-1c expression by activating CREB1. BMB Rep. 2007, 40, 525–531. [Google Scholar] [CrossRef]
- Hesketh, J.E.; Campbell, G.P.; Lobley, G.E.; Maltin, C.A.; Acamovic, F.; Palmer, R.M. Stimulation of actin and myosin synthesis in rat gastrocnemius muscle by clenbuterol; evidence for translational control. Comp. Biochem. Physiol. C Comp. Pharmacol. 1992, 102, 23–27. [Google Scholar] [CrossRef]
- Navegantes, L.C.C.; Resano, N.M.; Baviera, A.M.; Migliorini, R.H.; Kettelhut, I.C. Effect of sympathetic denervation on the rate of protein synthesis in rat skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E642–E647. [Google Scholar] [CrossRef]
- Kuiper, H.A.; Noordam, M.Y.; van Dooren-Flipsen, M.; Schilt, R.; Roos, A. Illegal use of β-adrenergic agonists: European Community. J. Anim. Sci. 1998, 76, 195–207. [Google Scholar] [CrossRef]
- Buttery, P.J.; Dawson, J.M. Growth promotion in farm animals. Proc. Nutr. Soc. 1990, 49, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Cini, M. The European Commission: Leadership, Organisation, and Culture in the EU Administration; Manchester University Press: Manchester, UK, 1996. [Google Scholar]
- Smith, D.; Paulson, G. Distribution, elimination, and residues of [14C] clenbuterol HCl in Holstein calves. J. Anim. Sci. 1997, 75, 454–461. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sauer, M.J.; Anderson, S.P. In vitro and in vivo studies of drug residue accumulation in pigmented tissues. Analyst 1994, 119, 2553–2556. [Google Scholar] [CrossRef] [PubMed]
- Boenisch, B.; Quirke, J. Safety Assessment of β-Agonists. In Vitro Toxicological Studies and Real Time Analysis of Residues in Food; RIKILT–DLO-Wageningen: Wageningen, The Netherlands, 1992; pp. 102–124. [Google Scholar]
- Jessen, S.; Solheim, S.A.; Jacobson, G.A.; Eibye, K.; Bangsbo, J.; Nordsborg, N.B.; Hostrup, M. Beta2-adrenergic agonist clenbuterol increases energy expenditure and fat oxidation, and induces mTOR phosphorylation in skeletal muscle of young healthy men. Drug Test. Anal. 2020, 12, 610–618. [Google Scholar] [CrossRef]
- Emili, M.; Stagni, F.; Salvalai, M.E.; Uguagliati, B.; Giacomini, A.; Albac, C.; Potier, M.-C.; Grilli, M.; Bartesaghi, R.; Guidi, S. Neonatal therapy with clenbuterol and salmeterol restores spinogenesis and dendritic complexity in the dentate gyrus of the Ts65Dn model of Down syndrome. Neurobiol. Dis. 2020, 140, 104874. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, R.A.; Elbialy, Z.I.; Abd El Latif, A.S.; Shukry, M.; Assar, D.H.; El Nokrashy, A.M.; Elsheshtawy, A.; Dawood, M.A.; Paray, B.A.; Van Doan, H. Dietary clenbuterol modifies the expression of genes involved in the regulation of lipid metabolism and growth in the liver, skeletal muscle, and adipose tissue of Nile tilapia (Oreochromis niloticus). Aquac. Rep. 2020, 17, 100319. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Ijiri, D.; Shimamoto, S.; Ishitani, K.; Nojima, T.; Ohtsuka, A. Single injection of the β2-adrenergic receptor agonist, clenbuterol, into newly hatched chicks alters abdominal fat pad mass in growing birds. Gen. Comp. Endocrinol. 2015, 211, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Ijiri, D.; Ishitani, K.; Shimamoto, S.; Ishimaru, Y.; Ohtsuka, A. The effects of intraperitoneal clenbuterol injection on protein degradation and myostatin expression differ between the sartorius and pectoral muscles of neonatal chicks. Gen. Comp. Endocrinol. 2014, 206, 111–117. [Google Scholar] [CrossRef]
- Buyse, J.; Decuypere, E.; Huyghebaert, G.; Herremans, M. The effect of clenbuterol supplementation on growth performance and on plasma hormone and metabolite levels of broilers. Poult. Sci. 1991, 70, 993–1002. [Google Scholar] [CrossRef]
- Ocampo, L.; Cortez, U.; Sumano, H.; Avila, E. Use of low doses of clenbuterol to reduce incidence of ascites syndrome in broilers. Poult. Sci. 1998, 77, 1297–1299. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Han, Z. Effects of dietary supplementation of (β2-adrenergic agonist clenbuterol on carcase characteristics and some metabolites in ducks. Br. Poult. Sci. 1994, 35, 355–361. [Google Scholar] [CrossRef]
- Dupont-Versteegden, E.E.; Katz, M.S.; McCarter, R.J. Beneficial versus adverse effects of long-term use of clenbuterol in mdx mice. Muscle Nerve 1995, 18, 1447–1459. [Google Scholar] [CrossRef] [PubMed]
- Witkowska-Piłaszewicz, O.; Pingwara, R.; Szczepaniak, J.; Winnicka, A. The Effect of the Clenbuterol—β2-Adrenergic Receptor Agonist on the Peripheral Blood Mononuclear Cells Proliferation, Phenotype, Functions, and Reactive Oxygen Species Production in Race Horses In Vitro. Cells 2021, 10, 936. [Google Scholar] [CrossRef] [PubMed]
- Pandit, R.J.; Hinsu, A.T.; Patel, N.V.; Koringa, P.G.; Jakhesara, S.J.; Thakkar, J.R.; Shah, T.M.; Limon, G.; Psifidi, A.; Guitian, J. Microbial diversity and community composition of caecal microbiota in commercial and indigenous Indian chickens determined using 16s rDNA amplicon sequencing. Microbiome 2018, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Havenstein, G.; Ferket, P.; Qureshi, M. Growth, livability, and feed conversion of 1957 versus 2001 broilers when fed representative 1957 and 2001 broiler diets. Poult. Sci. 2003, 82, 1500–1508. [Google Scholar] [CrossRef]
- Homma, H.; Shinohara, T. Effects of probiotic Bacillus cereus toyoi on abdominal fat accumulation in the Japanese quail (Coturnix japonica). Anim. Sci. J. 2004, 75, 37–41. [Google Scholar] [CrossRef]
- Nikolova, N.; Pavlovski, Z.; Milošević, N.; Perić, L. The quantity of abdominal fat in broiler chicken of different genotypes from fifth to seventh week of age. Biotechnol. Anim. Husb. 2007, 23, 331–338. [Google Scholar] [CrossRef]
- Taha, A.; El-Tahawy, A.; Abd El-Hack, M.; Swelum, A.; Saadeldin, I. Impacts of various storage periods on egg quality, hatchability, post-hatching performance, and economic benefit analysis of two breeds of quail. Poult. Sci. 2019, 98, 777–784. [Google Scholar] [CrossRef]
- Uni, Z.; Ferket, P.; Tako, E.; Kedar, O. In ovo feeding improves energy status of late-term chicken embryos. Poult. Sci. 2005, 84, 764–770. [Google Scholar] [CrossRef]
- Foye, O.; Uni, Z.; Ferket, P. Effect of in ovo feeding egg white protein, β-hydroxy-β-methylbutyrate, and carbohydrates on glycogen status and neonatal growth of turkeys. Poult. Sci. 2006, 85, 1185–1192. [Google Scholar] [CrossRef]
- Zulkifli, I.; Htin, N.N.; Alimon, A.; Loh, T.; Hair-Bejo, M. Dietary selection of fat by heat-stressed broiler chickens. Asian Australas. J. Anim. Sci. 2007, 20, 245–251. [Google Scholar] [CrossRef]
- Rodriguez, M.; Ortiz, L.; Alzueta, C.; Rebole, A.; Trevino, J. Nutritive value of high-oleic acid sunflower seed for broiler chickens. Poult. Sci. 2005, 84, 395–402. [Google Scholar] [CrossRef]
- Taha, A.E.; AbdAllah, O.A.; Attia, K.M.; El-Karim, A.; Ragaa, E.; El-Hack, A.; Mohamed, E.; El-Edel, M.A.; Saadeldin, I.M.; Hussein, E.O. Does in Ovo Injection of Two Chicken Strains with Royal Jelly Impact Hatchability, Post-Hatch Growth Performance and Haematological and Immunological Parameters in Hatched Chicks? Animals 2019, 9, 486. [Google Scholar] [CrossRef] [PubMed]
- Omar, E.N.; Jaafar, H.S. Halal supply chain in the food industry—A conceptual model. In Proceedings of the 2011 IEEE Symposium on Business, Engineering and Industrial Applications (ISBEIA), Langkawi, Malaysia, 25–28 September 2011; pp. 384–389. [Google Scholar]
- Baziz, H.A.; Geraert, P.; Padilha, J.; Guillaumin, S. Chronic heat exposure enhances fat deposition and modifies muscle and fat partition in broiler carcasses. Poult. Sci. 1996, 75, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Dein, F. Hematology in Avian Medicine and Surgery; WB Saunders: Philadelphia, PA, USA, 1986; pp. 174–191. [Google Scholar]
- Gross, W.B.; Siegel, H.S. Evaluation of the Heterophil/Lymphocyte Ratio as a Measure of Stress in Chickens. Avian Dis. 1983, 27, 972. [Google Scholar] [CrossRef]
- Akiba, Y.; Jensen, L.; Barb, C.; Kraeling, R. Plasma estradiol, thyroid hormones, and liver lipid content in laying hens fed different isocaloric diets. J. Nutr. 1982, 112, 299–308. [Google Scholar] [CrossRef]
- Park, K.-H.; Shin, D.-G.; Kim, J.-R.; Cho, K.-H. Senescence-Related Truncation and Multimerization of Apolipoprotein A-I in High-Density Lipoprotein with an Elevated Level of Advanced Glycated End Products and Cholesteryl Ester Transfer Activity. J. Gerontol. Ser. A 2010, 65A, 600–610. [Google Scholar] [CrossRef]
- Havel, R.J.; Eder, H.A.; Bragdon, J.H. The distribution and chemical composition of ultracentrifugally separated lipoproteins in human serum. J. Clin. Investig. 1955, 34, 1345–1353. [Google Scholar] [CrossRef]
- Reitman, S.; Frankel, S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef]
- Rudkin, F.M.; Bain, J.M.; Walls, C.; Lewis, L.E.; Gow, N.A.; Erwig, L.P. Altered dynamics of Candida albicans phagocytosis by macrophages and PMNs when both phagocyte subsets are present. MBio 2013, 4, e00810–e00813. [Google Scholar] [CrossRef]
- Schultz, L. Methods in Clinical Chemistry; The CV Mosby Co.: St Louis, MO, USA, 1987; pp. 742–746. [Google Scholar]
- Radwan, O.K.; Ahmed, R.F. Amendment effect of resveratrol on diclofenac idiosyncratic toxicity: Augmentation of the anti-inflammatory effect by assessment of Arachidonic acid and IL-1β levels. J. Pharm. Sci. 2016, 6, 170–177. [Google Scholar] [CrossRef]
- Hughes, M.; Kerry, J.; Arendt, E.; Kenneally, P.; McSweeney, P.; O’neill, E. Characterization of proteolysis during the ripening of semi-dry fermented sausages. Meat Sci. 2002, 62, 205–216. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Huang, J.B.; Zhang, Y.; Zhou, Y.B.; Wan, X.C.; Zhang, J.S. Effects of epigallocatechin gallate on lipid metabolism and its underlying molecular mechanism in broiler chickens. J. Anim. Physiol. Anim. Nutr. 2015, 99, 719–727. [Google Scholar] [CrossRef]
- Penha, A.M.; Schaeffel, F.; Feldkaemper, M. Insulin, insulin-like growth factor–1, insulin receptor, and insulin-like growth factor–1 receptor expression in the chick eye and their regulation with imposed myopic or hyperopic defocus. Mol. Vis. 2011, 17, 1436. [Google Scholar]
- Flees, J.; Rajaei-Sharifabadi, H.; Greene, E.; Beer, L.; Hargis, B.M.; Ellestad, L.; Porter, T.; Donoghue, A.; Bottje, W.G.; Dridi, S. Effect of Morinda citrifolia (Noni)-Enriched Diet on Hepatic Heat Shock Protein and Lipid Metabolism-Related Genes in Heat Stressed Broiler Chickens. Front. Physiol. 2017, 8, 919. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.J.; Wang, L.N.; Shu, G.; Wang, S.B.; Zhu, X.T.; Gao, P.; Xi, Q.Y.; Zhang, Y.L.; Zhang, Z.Q.; Jiang, Q.Y. Decorin-induced proliferation of avian myoblasts involves the myostatin/Smad signaling pathway. Poult. Sci. 2014, 93, 138–146. [Google Scholar] [CrossRef]
- Hausman, G.J.; Barb, C.R.; Fairchild, B.D.; Gamble, J.; Lee-Rutherford, L. Expression of genes for interleukins, neuropeptides, growth hormone receptor, and leptin receptor in adipose tissue from growing broiler chickens. Domest. Anim. Endocrinol. 2012, 43, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Jiang, Z.; Wang, X.; Luo, Y.; Li, X.; Liu, H. Role of the insulin-like growth factor system in epiphyseal cartilage on the development of Langshan and Arbor Acres chickens, Gallus domesticus. Poult. Sci. 2010, 89, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier Health Sciences: Amsterdam, The Netherlands, 2008. [Google Scholar]
- IBM Corp. IBM SPSS Statistics for Windows, Version 22.0; IBM Corp.: Armonk, NY, USA, 2013. [Google Scholar]
- Yossa, R.; Verdegem, M. Misuse of multiple comparison tests and underuse of contrast procedures in aquaculture publications. Aquaculture 2015, 437, 344–350. [Google Scholar] [CrossRef]
- Hinkle, R.T.; Hodge, K.M.; Cody, D.B.; Sheldon, R.J.; Kobilka, B.K.; Isfort, R.J. Skeletal muscle hypertrophy and anti-atrophy effects of clenbuterol are mediated by the β2-adrenergic receptor. Muscle Nerve 2002, 25, 729–734. [Google Scholar] [CrossRef]
- Robinson, N.E. Clenbuterol and the horse. AAEP Proc. 2000, 46, 229–233. [Google Scholar]
- Abdulredha, W.S. Effect of Clenbuterol using as weight loose on liver enzymes and lipids profile. Iraq Med J. 2019, 3, 52–55. [Google Scholar]
- Takahashi, K.; Akiba, Y.; Horiguchi, M. Effects of a beta-adrenergic agonist clenbuterol) on performance, carcase composition, hepatic microsomal mixed function oxidase and antibody production in female broilers treated with or without corticosterone. Br. Poult. Sci. 1993, 34, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Bobowiec, R.; Radymska-Wawrzyniak, K. Effect of -2 adrenergic agonist clenbuterol on plasma lipoprotein fractions, fatty acid content and muscle fiber types in chickens. Med. Weter. 1994, 50, 457. [Google Scholar]
- Hamano, Y.; Kobayashi, S.; Terashima, Y. Effect of beta-adrenergic agonist (Clenbuterol) on skeletal muscle protein turnover in broiler chicks fed either low or adequate protein diet. Anim. Sci. Technol. 1994, 65, 1105–1110. [Google Scholar]
- Spurlock, D.M.; McDaneld, T.G.; McIntyre, L.M. Changes in skeletal muscle gene expression following clenbuterol administration. BMC Genom. 2006, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hamano, Y. Influence of lipoic acid on lipid metabolism and β-adrenergic response to intravenous or oral administration of clenbuterol in broiler chickens. Reprod. Nutr. Dev. 2002, 42, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Yao, T.; Qin, Y.; Yang, X.; Li, J.; Li, J.; Gu, X. Clenbuterol distribution and residues in goat tissues after the repeated administration of a growth-promoting dose. J. Anal. Toxicol. 2015, 39, 465–471. [Google Scholar] [CrossRef][Green Version]
- Muramatsu, T.; Kakita, M.; Aoyagi, Y.; Okumura, J. Research note: β-adrenergic agonist effects on liver and breast muscle protein synthesis in female chicks. Poult. Sci. 1991, 70, 1630–1632. [Google Scholar] [CrossRef]
- Liu, P.; XIANG, J.-Z.; Zhao, L.; Yang, L.; Hu, B.-R.; Fu, Q. Effect of β2-adrenergic agonist clenbuterol on ischemia/reperfusion injury in isolated rat hearts and cardiomyocyte apoptosis induced by hydrogen peroxide. Acta Pharmacol. Sin. 2008, 29, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Webb, E.; Casey, N. Fatty acids in carcass fat of steers treated with a β-adrenergic agonist individually or in combination with trenbolone acetate+ oestradiol-17β. Meat Sci. 1995, 41, 69–76. [Google Scholar] [CrossRef]
- Kheiri, F.; Alibeyghi, M. Effect of different levels of lysine and threonine on carcass characteristics, intestinal microflora and growth performance of broiler chicks. Ital. J. Anim. Sci. 2017, 16, 580–587. [Google Scholar] [CrossRef]
- Abo, T.; Iida, R.h.; Kaneko, S.; Suga, T.; Yamada, H.; Hamada, Y.; Yamane, A. IGF and myostatin pathways are respectively induced during the earlier and the later stages of skeletal muscle hypertrophy induced by clenbuterol, a β2-adrenergic agonist. Cell Biochem. Funct. 2012, 30, 671–676. [Google Scholar] [CrossRef]
- Lalani, R.; Bhasin, S.; Byhower, F.; Tarnuzzer, R.; Grant, M.; Shen, R.; Asa, S.; Ezzat, S.; Gonzalez-Cadavid, N. Myostatin and insulin-like growth factor-I and-II expression in the muscle of rats exposed to the microgravity environment of the NeuroLab space shuttle flight. J. Endocrinol. 2000, 167, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Akutsu, S.; Wakana, N.; Morito, M.; Shimada, A.; Yamane, A. The expressions of insulin-like growth factors, their receptors, and binding proteins are related to the mechanism regulating masseter muscle mass in the rat. Arch. Oral Biol. 2006, 51, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Awede, B.L.; Thissen, J.-P.; Lebacq, J. Role of IGF-I and IGFBPs in the changes of mass and phenotype induced in rat soleus muscle by clenbuterol. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E31–E37. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Murray, R.D.; Shalet, S.M. Impact of growth hormone status on body composition and the skeleton. Horm. Res. Paediatr. 2004, 62, 35–41. [Google Scholar] [CrossRef]
- Jia, J.; Ahmed, I.; Liu, L.; Liu, Y.; Xu, Z.; Duan, X.; Li, Q.; Dou, T.; Gu, D.; Rong, H. Selection for growth rate and body size have altered the expression profiles of somatotropic axis genes in chickens. PLoS ONE 2018, 13, e0195378. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.; El Asely, A.; Abd El-Naby, A.S.; Samir, F.; El-Ashram, A.; Sudhakaran, R.; Dawood, M.A. Growth performance, intestinal histomorphology and growth-related gene expression in response to dietary Ziziphus mauritiana in Nile tilapia (Oreochromis niloticus). Aquaculture 2019, 512, 734301. [Google Scholar] [CrossRef]
- Zhang, B.; Yang, X.; Guo, Y.; Long, F. Effects of dietary lipids and Clostridium butyricum on serum lipids and lipid-related gene expression in broiler chickens. Animal 2011, 5, 1909–1915. [Google Scholar] [CrossRef] [PubMed]
- Finck, B.N.; Kelly, D.P. Peroxisome proliferator–activated receptor γ coactivator-1 (PGC-1) regulatory cascade in cardiac physiology and disease. Circulation 2007, 115, 2540–2548. [Google Scholar] [CrossRef]
- Kim, H.-K.; Della-Fera, M.A.; Hausman, D.B.; Baile, C.A. Effect of clenbuterol on apoptosis, adipogenesis, and lipolysis in adipocytes. J. Physiol. Biochem. 2010, 66, 197–203. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Experimental Diets | |
|---|---|---|
| Starter (0–4 weeks) | Grower (5–12 weeks) | |
| Yellow corn | 55.3 | 60.33 |
| Soybean meal, 48% | 38.47 | 33.1 |
| Soybean oil | 2.05 | 2.40 |
| Di-calcium phosphate | 1.73 | 1.75 |
| Limestone | 1.25 | 1.05 |
| Mineral and Vitamin premix 1 | 0.3 | 0.3 |
| Salt | 0.22 | 0.22 |
| Sodium bicarbonate | 0.32 | 0.25 |
| DL-methionine | 0.18 | 0.20 |
| Lysine | 0.19 | 0.40 |
| Nutrients Composition | ||
| Metabolizable energy (Kcal/kg) | 3000 | 3050 |
| Crude protein% | 23.04 | 21.05 |
| Crude fat% | 4.51 | 4.97 |
| Lysine% | 1.49 | 1.47 |
| Methionine% | 0.56 | 0.55 |
| Calcium% | 0.96 | 0.88 |
| Available phosphorous | 0.44 | 0.42 |
| Sodium | 0.2 | 0.18 |
| Gene | Forward | Reverse | Accession Number | Amplicon (pb) | References |
|---|---|---|---|---|---|
| GAPDH | ACATGGCATCCAAGGAGTGAG | GGGGAGACAGAAGGGAACAGA | NM_204305 | 158 | [51] |
| IGF1R | TTCAGGAACCAAAGGGCGA | TGTAATCTGGAGGGCGATACC | NM_205032 | 167 | [52] |
| PPARα | TGTGGAGATCGTCCTGGTCT | CGTCAGGATGGTTGGTTTGC | NM_001001464 | 103 | [51] |
| SREBP-1c | TCACCGCTTCTTCGTGGAC | CTGAAGGTACTCCAACGCATC | AY029224 | 220 | [51] |
| FAS | CAATGGACTTCATGCCTCGGT | GCTGGGTACTGGAAGACAAACA | NM_205155.2 | 119 | [51] |
| LPL | GTGACCAAGGTAGACCAGCC | GAAGAGACTTCAGGCAGCGT | NM_205282.1 | 62 | [53] |
| MSTN | TTACCCAAAGCTCCTCCACTG | AGGATCTGCACAAACACCGT | NM_001001461 | 120 | [54] |
| GHR | CATGGCCACCTTTTGCAGAC | ACCTTGGATTTCTGCCCTGG | NM_001001293 | 121 | [55] |
| IGFBP2 | CACAACCACGAGGACTCAAA | CATTCACCGACATCTTGCAC | NM_205359.1 | 299 | [56] |
| Parameters | Control | Normal Saline | Clenbuterol | p-Value | Polynomial Contrasts | |||
|---|---|---|---|---|---|---|---|---|
| 10 ppm | 15 ppm | 20 ppm | Linear | Quadratic | ||||
| Hatchability (%) | 84 ± 1.3 b | 82.6 ± 2.2 b | 89.9 ± 3.2 a | 90.2 ± 1.5 a | 90.3 ± 3.4 a | 0.046 | 0.012 | 0.001 |
| IBW (g/bird) | 31.1 ± 0.4 | 31.1 ± 1.4 | 30.3 ± 0.4 | 30.1 ± 0.8 | 30.2 ± 0.7 | 0.110 | 0.114 | 0.223 |
| FBW (g/bird) | 2680.1 ± 29.2 b | 2679.5 ± 33.5 b | 2815.5 ± 17.2 a | 2938.2 ± 25.7 a | 2655.4 ± 25.8 b | 0.021 | 0.001 | 0.024 |
| WG (g/bird) | 2649.2 ± 25.5 b | 2648.4 ± 3.4 b | 2785.2 ± 3.5 ab | 2908.0 ± 7.4 a | 2625.2 ± 3.7 b | 0.001 | 0.01 | 0.0254 |
| FCR (g feed/gain) | 1.6 ± 3.2 | 1.59 ± 4.8 | 1.52 ± 4.3 | 1.47 ± 3.4 | 1.65 ± 7.2 | 0.165 | 0.021 | 0.214 |
| Carcass (%) | 72.5 ± 1.2 b | 71.5 ± 0.4 b | 73.15 ± 3.1 b | 75.54 ± 2.1 a | 72.15 ± 1.4 b | 0.002 | 0.015 | 0.014 |
| Abdominal Fat (% of eviscerated weight) | 2.7 ± 0.1 a | 2.75 ± 0.1 a | 2.014 ± 0.2 b | 1.86 ± 0.1 c | 2.11 ± 0.1 b | 0.002 | 0.014 | 0.018 |
| Parameters | Control | Normal Saline | Clenbuterol | p-Value | Polynomial Contrasts | |||
|---|---|---|---|---|---|---|---|---|
| 10 ppm | 15 ppm | 20 ppm | Linear | Quadratic | ||||
| RBC (×106/μL) | 4.01 ± 0.32 | 3.91 ± 0.32 | 3.75 ± 0.14 | 3.71 ± 0.21 | 3.47 ± 0.14 | 0.125 | 0.214 | 0.31 |
| Hemoglobin (g/dL) | 12.05 ± 0.21 | 11.88 ± 0.3 | 11.52 ± 0.32 | 12.14 ± 0.5 | 11.50 ± 0.21 | 0.121 | 0.125 | 0.25 |
| H/L ratio% | 0.56 ± 0.01 | 0.57 ± 0.04 | 0.59 ± 0.14 | 0.60 ± 0.15 | 0.62 ± 0.32 | 0.11 | 0.31 | 0.45 |
| Cholesterol (mg/dL) | 110.54 ± 5.4 a | 112.5 ± 1.3 a | 82.14 ± 1.4 c | 79.15 ± 2.3 bc | 85.05 ± 3.2 b | 0.241 | 0.01 | 0.035 |
| TG (mg/dL) | 103.15 ± 6.2 a | 106.4 ± 4.4 a | 92.45 ± 3.4 c | 85.45 ± 2.4 d | 101 ± 2.1 b | 0.014 | 0.021 | 0.01 |
| Total lipids (mg/dL) | 198.9 ± 4.1 a | 200.1 ± 2.2 a | 156.36 ± 3.4 c | 150.05 ± 3.1 c | 161.56 ± 4.1 b | 0.018 | 0.015 | 0.021 |
| HDL-C (mg/dL) | 55.1 ± 2.1 | 54.2 ± 1.4 | 57.6 ± 2.1 | 59.45 ± 2.5 | 56.48 ± 1.1 | 0.144 | 0.144 | 0.21 |
| LDL-C (mg/dL) | 79.45 ± 3.4 a | 79.15 ± 2.1 a | 75.58 ± 2.1 c | 74.45 ± 2.1 c | 78.15 ± 1.4 b | 0.001 | 0.014 | 0.014 |
| Albumin (g/dL) | 1.92 ± 0.2 b | 1.99 ± 0.14 b | 2.12 ± 0.24 a | 2.14 ± 0.3 a | 1.95 ± 0.1 b | 0.001 | 0.001 | 0.01 |
| Globulin (g/dL) | 2.12 ± 0.14 | 2.10 ± 0.1 | 2.15 ± 0.5 | 2.20 ± 0.3 | 2.10 ± 0.4 | 0.125 | 0.14 | 0.12 |
| Total protein (g/dL) | 4.04 ± 0.15 b | 4.09 ± 0.14 b | 4.27 ± 0.3 a | 4.34 ± 0.4 a | 4.05 ± 0.5 b | 0.011 | 0.02 | 0.001 |
| AST (U/L) | 112.10 ± 4.1 | 110.15 ± 2.0 | 111.05 ± 3.0 | 115.45 ± 3.1 | 116.48 ± 4.1 | 0.125 | 0.45 | 0.32 |
| ALT (U/L) | 12.15 ± 0.4 | 12.23 ± 1.2 | 10.95 ± 0.8 | 11.02 ± 0.9 | 11.47 ± 0.4 | 0.154 | 0.158 | 0.28 |
| TAC (U/L) | 1.81 ± 0.2 | 1.83 ± 0.2 | 1.82 ± 0.1 | 1.83 ± 0.3 | 1.80 ± 0.4 | 0.121 | 0.147 | 0.14 |
| Parameters | Control | Normal Saline | Clenbuterol | p-Value | Polynomial Contrasts | |||
|---|---|---|---|---|---|---|---|---|
| 10 ppm | 15 ppm | 20 ppm | Linear | Quadratic | ||||
| GSH-PX(U/g) | 25.14 ± 1.2 | 25.22 ± 1.2 | 26.45 ± 2.1 | 26.48 ± 1.2 | 24.91 ± 1.8 | 0.114 | 0.112 | 0.51 |
| MDA (nmol/g) | 10.155 ± 1.1 | 10.78 ± 1.1 | 10.19 ± 1.1 | 10.48 ± 1.4 | 10.54 ± 0.5 | 0.125 | 0.102 | 0.32 |
| SOD (U/mg protein) | 45.45 ± 2.0 | 46.112 ± 2.1 | 45.47 ± 3.2 | 46.2 ± 2.1 | 44.78 ± 2.4 | 0.214 | 0.25 | 0.15 |
| Phagocytic activity (%) | 52.15 ± 2.4 | 51.14 ± 2.0 | 54.78 ± 1.4 | 55.78 ± 0.3 | 52.45 ± 1.4 | 0.101 | 0.45 | 0.21 |
| Phagocytic index | 3.42 ± 10.4 | 3.32 ± 0.1 | 3.30 ± 1.2 | 3.56 ± 0.3 | 3.22 ± 0.4 | 0.15 | 0.25 | 0.45 |
| Lysozyme-activity (μ/mL) | 0.89 ± 0.04 | 0.92 ± 0.1 | 0.93 ± 0.4 | 0.91 ± 0.4 | 0.90 ± 0.02 | 0.145 | 0.27 | 0.32 |
| Fatty Acids | Control | Normal Saline | Clenbuterol | p-Value | Polynomial Contrasts | |||
|---|---|---|---|---|---|---|---|---|
| 10 ppm | 15 ppm | 20 ppm | Linear | Quadratic | ||||
| SFA | ||||||||
| C16:0 | 29.24 ± 2.1 a | 29.12 ± 1.2 a | 25.3 ± 1.1 b | 20.14 ± 1.1 c | 25.4 ± 1.3 b | 0.214 | 0.001 | 0.21 |
| C16:1 | 3.75 ± 0.5 a | 3.74 ± 0.3 a | 3.53 ± 0.4 b | 3.62 ± 0.4 b | 3.60 ± 0.02 b | 0.001 | 0.01 | 0.023 |
| C18:0 | 9.45 ± 0.8 a | 9.42 ± 0.7 a | 7.48 ± 0.7 bc | 6.34 ± 0.3 c | 8.65 ± 0.5 b | 0.001 | 0.012 | 0.04 |
| C14:0 | 1.18 ± 0.1 a | 1.115 ± 0.3 a | 0.93 ± 0.02 bc | 0.75 ± 0.02 c | 1.02 ± 0.01 b | 0.001 | 0.01 | 0.01 |
| MUFA | ||||||||
| C18:1 | 18.24 ± 1.1 | 18.12 ± 0.7 | 18.11 ± 1.0 | 18.04 ± 1.1 | 18.36 ± 0.9 | 0.145 | 0.63 | 0.81 |
| PUFA | ||||||||
| C18:2 | 23.14 ± 1.2 | 23.22 ± 2.1 | 23.33 ±1.2 | 23.40 ± 1.1 | 23.45 ± 1.2 | 0.121 | 0.365 | 0.75 |
| C18:3n3 | 1.62 ± 0.3 | 1.60 ± 0.4 | 1.68 ± 0. 1 | 1.65 ± 0.4 | 1.64 ± 0.1 | 0.132 | 0.225 | 0.89 |
| C18:2n6 | 0.92 ± 0.04 | 0.90 ± 0.07 | 0.88 ± 0.1 | 0.86 ± 0.1 | 0.89 ± 0.1 | 0.116 | 0.23 | 0.52 |
| C22:6n3 | 0.84 ± 0.01 | 0.88 ± 0.08 | 0.82 ± 0.2 | 0.79 ± 0.1 | 0.80 ± 0.1 | 0.132 | 0.14 | 0.95 |
| Amino Acids | Control | Normal Saline | Clenbuterol | ||
|---|---|---|---|---|---|
| 10 ppm | 15 ppm | 20 ppm | |||
| Leucine | 6.76 ± 0.5 b | 6.77 ± 0.4 b | 6.85 ± 0.8 b | 6.89 ± 0.7 a | 6.79 ± 0.2 b |
| Isoleucine | 3.28 ± 0.1 b | 3.20 ± 0.1 b | 3.43 ± 0.1 a | 3.52 ± 0.1 a | 3.30 ± 0.1 b |
| Phenylalanine | 2.12 ± 0.3 c | 2.10 ± 0.1 c | 2.55 ± 0.1 a | 2.69 ± 0.1 a | 2.20 ± 0.1 b |
| Lysine | 7.30 ± 0.8 b | 7.29 ± 0.9 b | 7.45 ± 0.3 a | 7.50 ± 0.5 a | 7.42 ± 0.8 a |
| Methionine | 2.14 ± 0.1 b | 2.22 ± 0.1 b | 2.30 ± 0.2 a | 2.39 ± 0.3 a | 2.20 ± 0.1 b |
| Valine | 4.02 ± 0.2 c | 4.1 ± 0.1 c | 4.22 ± 0.1 a | 4. 33 ± 0.4 a | 4.11 ± 0.1 b |
| Threonine | 3.75 ± 0.1 b | 3.70 ± 0.2 b | 3.90 ± 0.2 a | 3.95 ± 0.1 a | 3.72 ± 0.1 b |
| Serine | 3.51 ± 0.1 c | 3.55 ± 0.2 c | 3.69 ± 0.2 b | 3.77 ± 0.1 a | 3.52 ± 0.1 c |
| Glycine | 3.42 ± 0.4 | 3.44 ± 0.2 | 3.44 ± 0.3 | 3.40 ± 0.1 | 3.33 ± 0.2 |
| Glutamic acid | 11.55 ± 0.4 | 11.45 ± 0.7 | 11.57 ± 0.8 | 11.53 ± 0.6 | 11.49 ± 0.4 |
| Aspartic acid | 8.3 ± 0.3 c | 7.98 ± 0.2 c | 8.15 ± 0.4 b | 8.20 ± 0.4 a | 8.10 ± 0.8 bc |
| Alanine | 4.92 ± 0.2 b | 4.88 ± 0.4 b | 4.95 ± 0.1 a | 5.05 ± 0.4 a | 4.89 ± 001 b |
| Tyrosine | 2.81 ± 0.1 | 2.80 ± 0.2 | 2.82 ± 0.3 | 2.89 ± 0.2 | 2.83 ± 0.5 |
| Cysteine | 1.69 ± 0.1 | 1.73 ± 0.1 | 1.72 ± 0.3 | 1.75 ± 0.3 | 1.68 ± 0.3 |
| Histidine | 3.15 ± 0.3 | 3.22 ± 0.4 | 3.12 ± 0.2 | 3.22 ± 0.2 | 3.19 ± 0.3 |
| Arginine | 5.14 ± 0.2 c | 5.12 ± 0.4 c | 5.22 ± 0.2 b | 5.35 ± 0.1 a | 5.12 ± 0.2 c |
| Proline | 2.19 ± 0.1 b | 2.24 ± 0.1 b | 2.21 ± 0.1 b | 2.41 ± 0.2 a | 2.18 ± 0.1 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saleh, A.A.; Alhotan, R.A.; Alharthi, A.S.; Nassef, E.; Kassab, M.A.; Farrag, F.A.; Hendam, B.M.; Abumnadour, M.M.A.; Shukry, M. Insight View on the Role of in Ovo Feeding of Clenbuterol on Hatched Chicks: Hatchability, Growth Efficiency, Serum Metabolic Profile, Muscle, and Lipid-Related Markers. Animals 2021, 11, 2429. https://doi.org/10.3390/ani11082429
Saleh AA, Alhotan RA, Alharthi AS, Nassef E, Kassab MA, Farrag FA, Hendam BM, Abumnadour MMA, Shukry M. Insight View on the Role of in Ovo Feeding of Clenbuterol on Hatched Chicks: Hatchability, Growth Efficiency, Serum Metabolic Profile, Muscle, and Lipid-Related Markers. Animals. 2021; 11(8):2429. https://doi.org/10.3390/ani11082429
Chicago/Turabian StyleSaleh, Ahmed A., Rashed A. Alhotan, Abdulrahman S. Alharthi, Eldsokey Nassef, Mohamed A. Kassab, Foad A. Farrag, Basma M. Hendam, Mohamed M. A. Abumnadour, and Mustafa Shukry. 2021. "Insight View on the Role of in Ovo Feeding of Clenbuterol on Hatched Chicks: Hatchability, Growth Efficiency, Serum Metabolic Profile, Muscle, and Lipid-Related Markers" Animals 11, no. 8: 2429. https://doi.org/10.3390/ani11082429
APA StyleSaleh, A. A., Alhotan, R. A., Alharthi, A. S., Nassef, E., Kassab, M. A., Farrag, F. A., Hendam, B. M., Abumnadour, M. M. A., & Shukry, M. (2021). Insight View on the Role of in Ovo Feeding of Clenbuterol on Hatched Chicks: Hatchability, Growth Efficiency, Serum Metabolic Profile, Muscle, and Lipid-Related Markers. Animals, 11(8), 2429. https://doi.org/10.3390/ani11082429