
Isotopic Niche of Syntopic Granivores in Commercial Orchards and Meadows

 ,
, 
 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Small Mammal Trapping and Sample Size
2.3. Stable Isotope Analysis
2.4. Statistical Analyses
3. Results
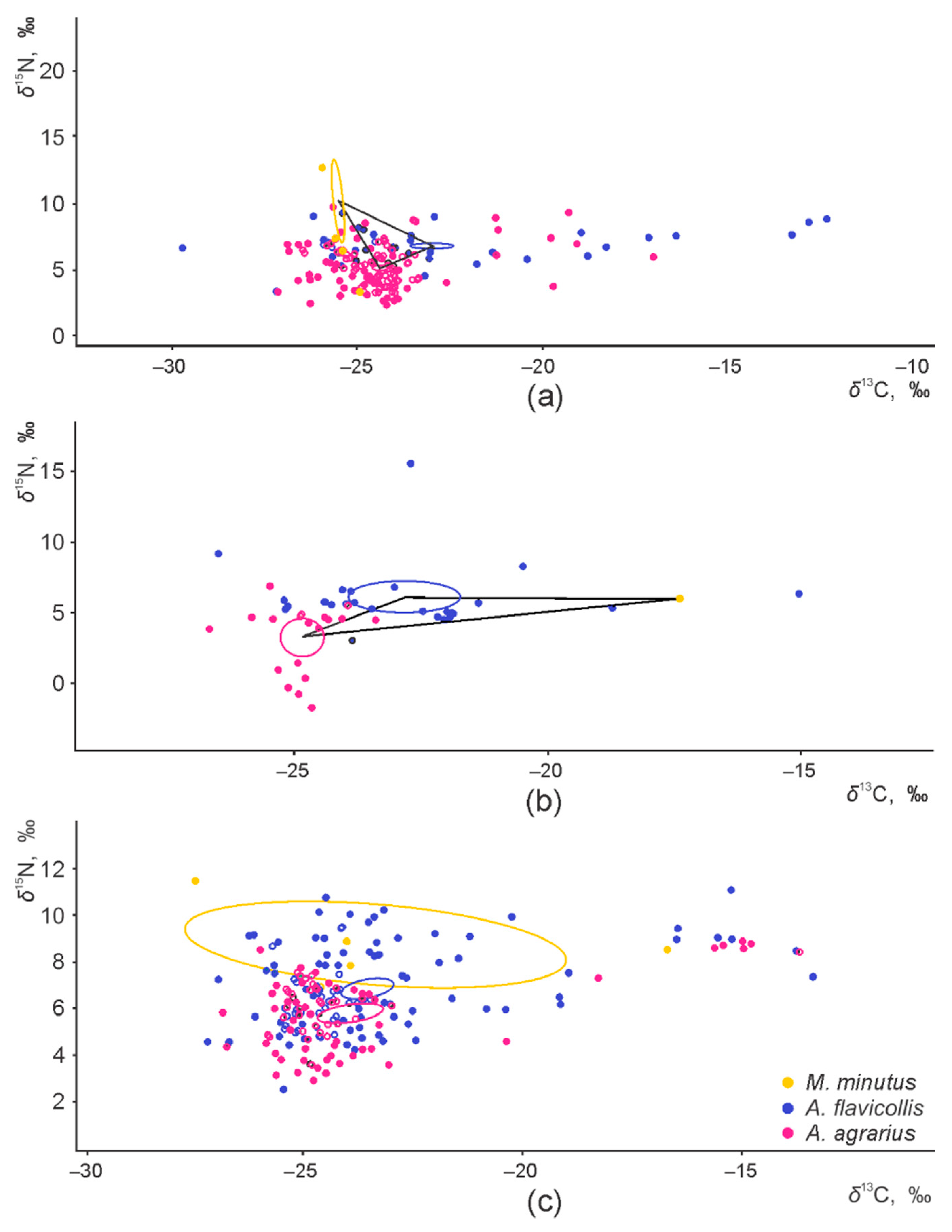
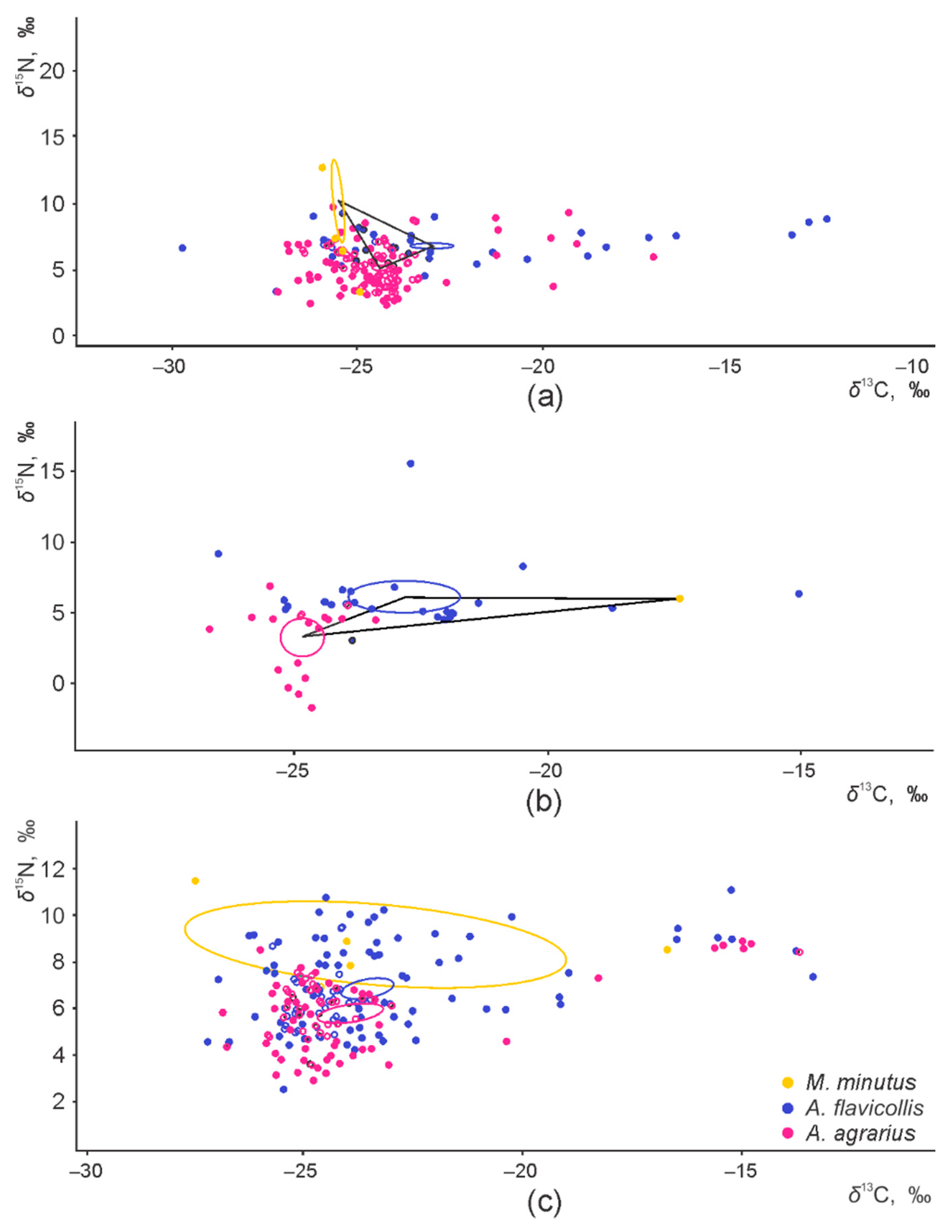
3.1. Sympatry and Syntopy of the Granivore Species
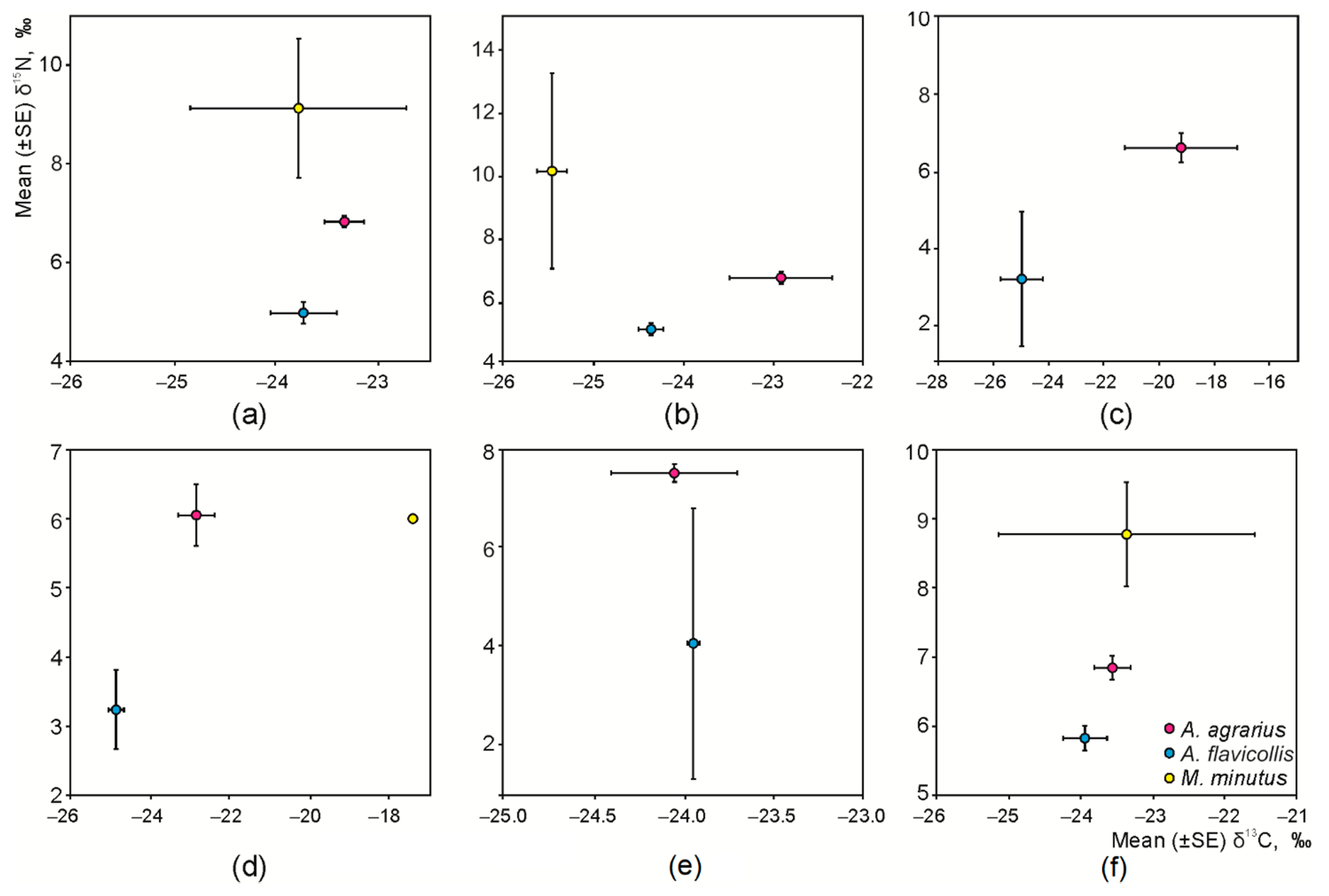
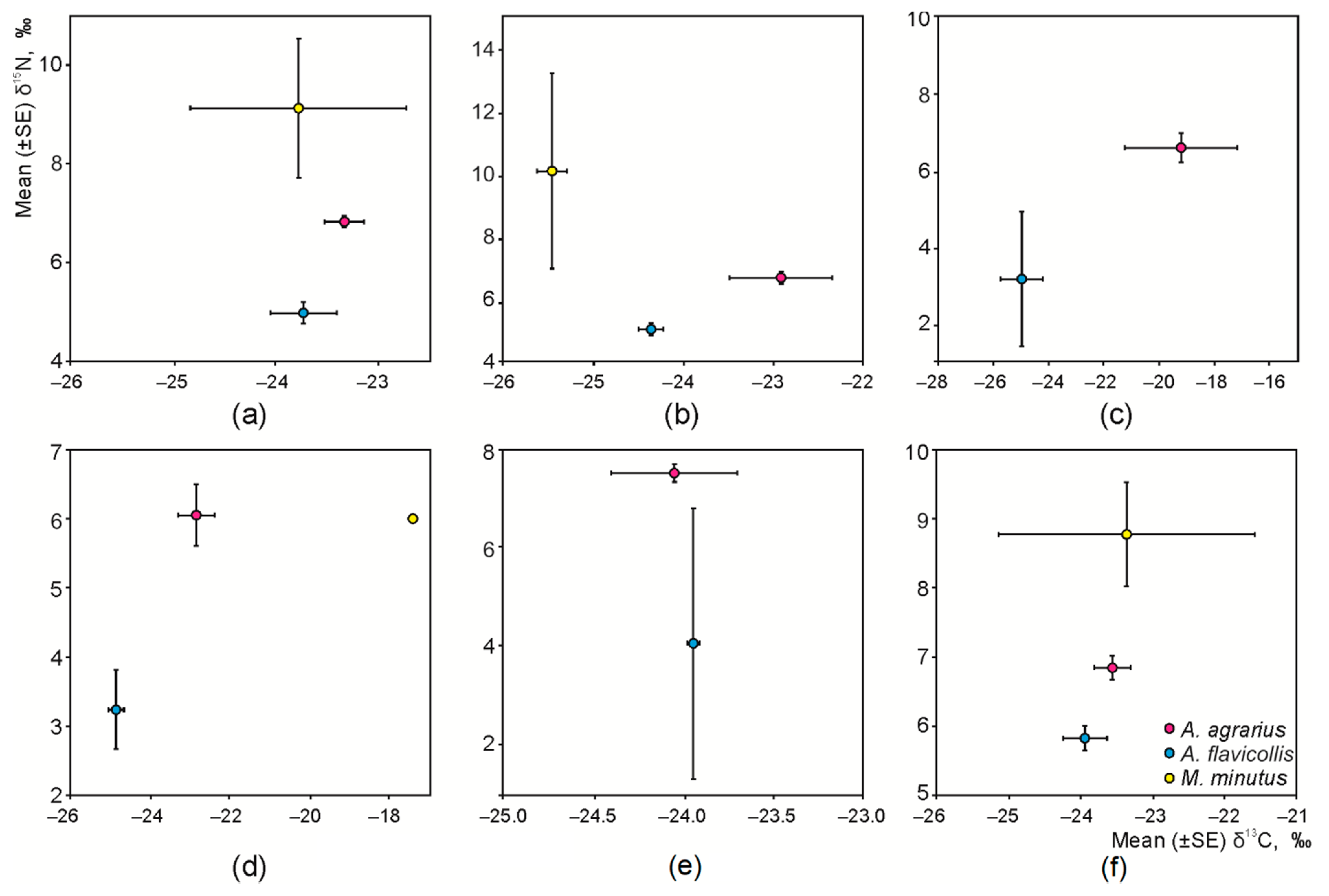
3.2. Interspecific and Habitat-Based Differences of Isotopic Niche in Syntopic Granivores
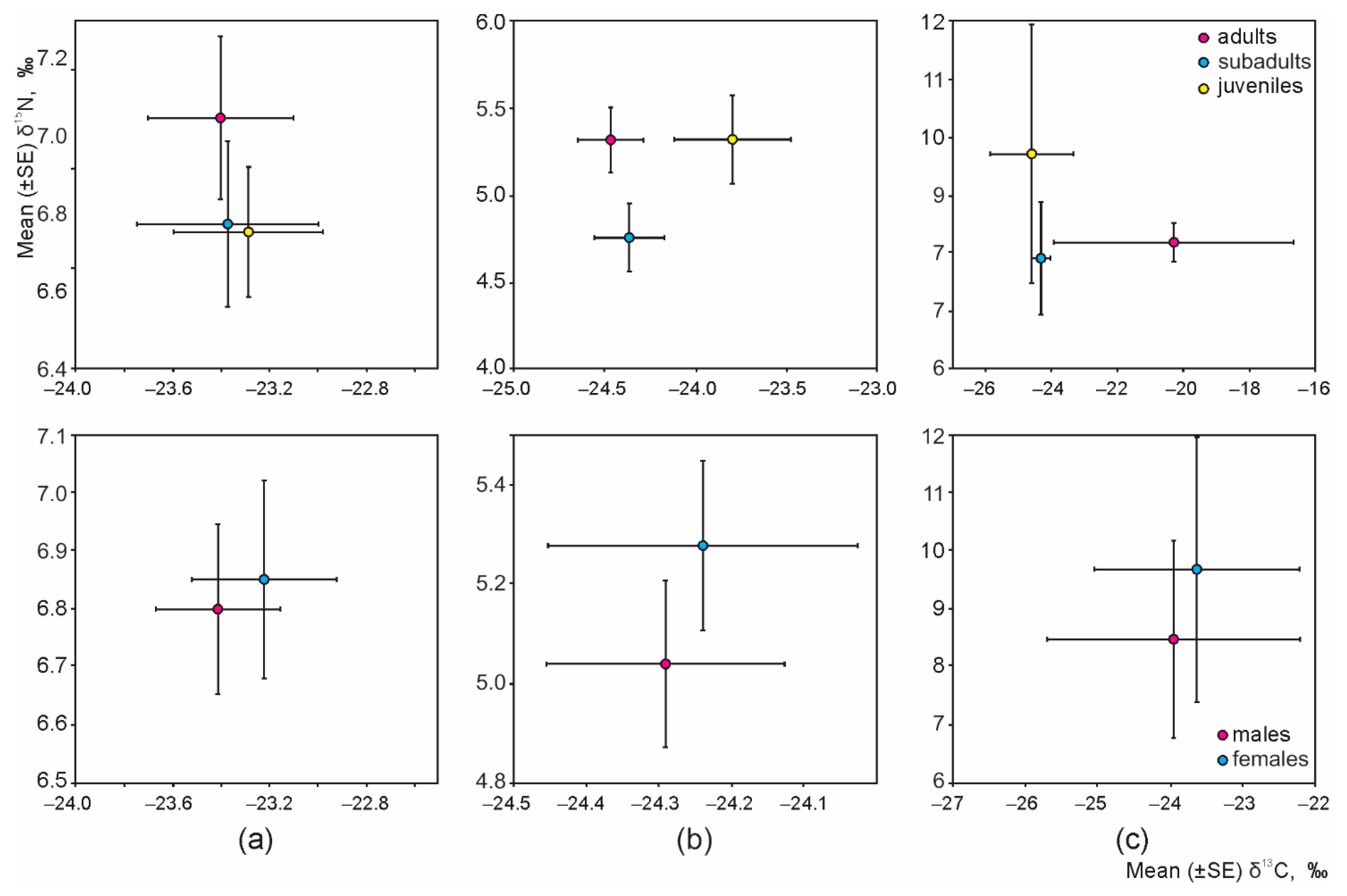
3.3. Intraspecific Differences of the Isotopic Niche in Syntopic Granivores
4. Discussion
5. Conclusions
- Syntopic living determines the separation of granivores in the isotopic space with full separation of species according to δ15N values, this presuming different amounts of food of animal origin in their diet.
- The separation of the core dietary niches in the apple orchards and raspberry plantations proved that these agricultural habitats are resource-scarce for granivores.
- Intraspecific differences of the isotopic niche were not present in any of the three granivorous species.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dickman, C.R. Habitat utilization and diet of the harvest mouse Micromys minutus, in an urban environment. Acta Theriol. 1986, 31, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Liro, A.; Szacki, J. Movements of field mice Apodemus agrarius (Pallas) in a suburban mosaic of habitats. Oecologia 1987, 74, 438–440. [Google Scholar] [CrossRef]
- Herczeg, R.; Horvath, G.F. Species composition and nestedness of small mammal assemblages in two disturbed marshlands. North.-West. J. Zool. 2015, 11, 183–193. [Google Scholar]
- Vecsernyés, F. Autumn habitat selection of the harvest mouse (Micromys minutus Pallas, 1771) in a rural and fragmented landscape. Rev. Suisse Zool. 2020, 126, 111–125. [Google Scholar] [CrossRef]
- Amori, G.; Hutterer, R.; Kryštufek, B.; Yigit, N.; Mitsainas, G.; Palomo, L. Apodemus flavicollis (amended version of 2016 assessment). IUCN Red List Threat. Species 2021, e.T1892A197269879. [Google Scholar] [CrossRef]
- Kaneko, Y.; Kryštufek, B.; Zagarondnyuk, I.; Vohralík, V.; Batsaikhan, N.; Avirmed, D.; Sukhchuluun, G. Apodemus agrarius (errata version published in 2017). IUCN Red List Threat. Species 2016, e.T1888A115057408. [Google Scholar] [CrossRef]
- Kryštufek, B.; Lunde, D.P.; Meinig, H.; Aplin, K.; Batsaikhan, N.; Henttonen, H. Micromys minutus. IUCN Red List Threat. Species 2019, e.T13373A119151882. [Google Scholar] [CrossRef]
- Stefke, K.; Landler, L. Long-term monitoring of rodent and shrew communities in a biodiversity hot-spot in Austria using barn owl (Tyto alba) pellets. Acta Oecol. 2020, 109, 103660. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Skipitytė, R.; Balčiauskienė, L.; Jasiulionis, M. Resource partitioning confirmed by isotopic signatures allows small mammals to share seasonally flooded meadows. Ecol. Evol. 2019, 9, 5479–5489. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Balčiauskienė, L.; Stirkė, V. Mow the Grass at the Mouse’s Peril: Diversity of Small Mammals in Commercial Fruit Farms. Animals 2019, 9, 334. [Google Scholar] [CrossRef] [Green Version]
- Walsh, R.E.; Aprígio Assis, A.P.; Patton, J.L.; Marroig, G.; Dawson, T.E.; Lacey, E.A. Morphological and dietary responses of chipmunks to a century of climate change. Glob. Change Biol. 2016, 22, 3233–3252. [Google Scholar] [CrossRef] [PubMed]
- Abt, K.F.; Bock, W.F. Seasonal variations of diet composition in farmland field mice Apodemus spp. and bank voles Clethrionomys glareolus. Acta Theriol. 1998, 43, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Butet, A.; Delettre, Y.R. Diet differentiation between European arvicoline and murine rodents. Acta Theriol. 2011, 56, 297. [Google Scholar] [CrossRef]
- Pineda-Munoz, S.; Alroy, J. Dietary characterization of terrestrial mammals. P. Roy. Soc. B-Biol. Sci. 2014, 281, 20141173. [Google Scholar] [CrossRef] [PubMed]
- Dayan, T.; Simberloff, D. Morphological relationships among coexisting Heteromyids: An incisive dental character. Am. Nat. 1994, 143, 462–477. [Google Scholar] [CrossRef]
- Brylski, P.; Hall, B. Ontogeny of a macroevolutionary phenotype: The external cheek pouches of geomyoid rodents. Evolution 1988, 42, 391–395. [Google Scholar] [CrossRef]
- Fischer, C.; Gayer, C.; Kurucz, K.; Riesch, F.; Tscharntke, T.; Batáry, P. Ecosystem services and disservices provided by small rodents in arable fields: Effects of local and landscape management. J. Appl. Ecol. 2018, 55, 548–558. [Google Scholar] [CrossRef] [Green Version]
- Luque-Larena, J.J.; Mougeot, F.; Roig, D.V.; Lambin, X.; Rodríguez-Pastor, R.; Rodríguez-Valín, E.; Anda, P.; Escudero, R. Tularemia outbreaks and common vole (Microtus arvalis) irruptive population dynamics in northwestern Spain, 1997–2014. Vector-Borne Zoonot. 2015, 15, 568–570. [Google Scholar] [CrossRef] [PubMed]
- Zwolak, R.; Crone, E.E. Quantifying the outcome of plant–granivore interactions. Oikos 2012, 121, 20–27. [Google Scholar] [CrossRef]
- Jansen, P.A.; Forget, P.M. Scatterhoarding rodents and tree regeneration. In Nouragues. Dynamics and Plant–Animal Interactions in a Neotropical Rainforest; Bongers, F., Charles-Dominique, P., Forget, P.M., Théry, M., Eds.; Kluver Academic Publishers: Dordrecht, The Netherlands, 2001; pp. 275–288. [Google Scholar]
- Reichman, O.J. Relation of desert rodent diets to available resources. J. Mamm. 1975, 56, 731–751. [Google Scholar] [CrossRef]
- Bartel, S.L.; Orrock, J.L. An omnivorous mesopredator modifies predation of omnivore-dispersed seeds. Ecosphere 2021, 12, e03369. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Skipitytė, R.; Garbaras, A.; Stirkė, V.; Balčiauskienė, L.; Remeikis, V. Stable Isotopes Reveal the Dominant Species to Have the Widest Trophic Niche of Three Syntopic Microtus Voles. Animals 2021, 11, 1814. [Google Scholar] [CrossRef]
- Lancaster, J.; Pillay, N. Behavioral interactions between a coexisting rodent Micaelamys namaquensis and macroscelid Elephantulus myurus. Curr. Zool. 2010, 56, 395–400. [Google Scholar] [CrossRef]
- Pianka, E.R. Niche overlap and diffuse competition. Proc. Natl. Acad. Sci. USA 1974, 71, 2141–2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, E.M.; Paise, G. Temporal niche overlap among insectivorous small mammals. Integr. Zool. 2011, 6, 375–386. [Google Scholar] [CrossRef]
- Castro-Arellano, I.; Lacher, T.E. Temporal niche segregation in two rodent assemblages of subtropical Mexico. J. Trop. Ecol. 2009, 25, 593–603. [Google Scholar] [CrossRef]
- Zhong, W.; Wang, G.; Zhou, Q.; Ma, L.; Wan, X.; Liu, W. Spatial niche partitioning of coexisting small mammals in sand dunes. Ital. J. Zool. 2016, 83, 248–254. [Google Scholar] [CrossRef]
- Magomedov, M.S. Microhabitat partitioning in a rodent community in the arid conditions of the South-western Caspian Lowland. J. Vertebr. Biol. 2021, 70, 20091. [Google Scholar] [CrossRef]
- Stephens, R.B.; Hobbie, E.A.; Lee, T.D.; Rowe, R.J. Pulsed resource availability changes dietary niche breadth and partitioning between generalist rodent consumers. Ecol. Evol. 2019, 9, 10681–10693. [Google Scholar] [CrossRef] [Green Version]
- Stirkė, V.; Balčiauskas, L.; Balčiauskienė, L. Common Vole as a Focal Small Mammal Species in Orchards of the Northern Zone. Diversity 2021, 13, 134. [Google Scholar] [CrossRef]
- Kronfeld, N.; Dayan, T. A new method of determining diets of rodents. J. Mamm. 1998, 79, 1198–1202. [Google Scholar] [CrossRef]
- Hansson, L. Methods of morphological diet microanalysis in rodents. Oikos 1970, 21, 255–266. [Google Scholar] [CrossRef]
- Franco-Trecu, V.; Drago, M.; Riet-Sapriza, F.G.; Parnell, A.; Frau, R.; Inchausti, P. Bias in diet determination: Incorporating traditional methods in Bayesian mixing models. PLoS ONE 2013, 8, e80019. [Google Scholar] [CrossRef] [Green Version]
- Zarzoso-Lacoste, D.; Bonnaud, E.; Corse, E.; Gilles, A.; Meglecz, E.; Costedoat, C.; Gouni, A.; Vidal, E. Improving morphological diet studies with molecular ecology: An application for invasive mammal predation on island birds. Biol. Conserv. 2016, 193, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Missagia, R.V.; Patterson, B.D.; Perini, F.A. Stable isotope signatures and the trophic diversification of akodontine rodents. Evol. Ecol. 2019, 33, 855–872. [Google Scholar] [CrossRef]
- Verde Arregoitia, L.D.; D’Elía, G. Classifying rodent diets for comparative research. Mammal. Rev. 2021, 51, 51–65. [Google Scholar] [CrossRef]
- Shiels, A.B.; Flores, C.A.; Khamsing, A.; Krushelnycky, P.D.; Mosher, S.M.; Drake, D.R. Dietary niche differentiation among three species of invasive rodents (Rattus rattus, R. exulans, Mus musculus). Biol. Invasions 2013, 15, 1037–1048. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Skipitytė, R.; Jasiulionis, M.; Trakimas, G.; Balčiauskienė, L.; Remeikis, V. The impact of Great Cormorants on biogenic pollution of land ecosystems: Stable isotope signatures in small mammals. Sci. Total Environ. 2016, 565, 376–383. [Google Scholar] [CrossRef] [PubMed]
- Balčiauskas, L. Methods of Investigation of Terrestrial Ecosystems; Part. I. Animal Surveys; VU Leidykla: Vilnius, Lithuania, 2004; p. 183. [Google Scholar]
- Rezzani, R.; Nardo, L.; Favero, G.; Peroni, M.; Rodella, L.F. Thymus and aging: Morphological, radiological, and functional overview. Age 2014, 36, 313–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balciauskas, L.; Balciauskiene, L.; Janonyte, A. Reproduction of the root vole (Microtus oeconomus) at the edge of its distribution range. Turk. J. Zool. 2012, 36, 668–675. [Google Scholar]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER—Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef]
- Codron, J.; Duffy, K.J.; Avenant, N.L.; Sponheimer, M.; Leichliter, J.; Paine, O.; Sandberg, P.; Codron, D. Stable isotope evidence for trophic niche partitioning in a South African savanna rodent community. Curr. Zool. 2015, 61, 397–411. [Google Scholar] [CrossRef]
- Polis, G.A. Age structure component of niche width and intraspecific resource partitioning: Can age groups function as ecological species? Am. Nat. 1984, 123, 541–564. [Google Scholar] [CrossRef]
- Sheppard, C.E.; Inger, R.; McDonald, R.A.; Barker, S.; Jackson, A.L.; Thompson, F.J.; Vitikainen, E.I.K.; Cant, M.A.; Marshall, H.H. Intragroup competition predicts individual foraging specialisation in a group-living mammal. Ecol. Lett. 2018, 21, 665–673. [Google Scholar] [CrossRef]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Symes, C.T.; Wilson, J.W.; Woodborne, S.M.; Shaikh, Z.S.; Scantlebury, M. Resource partitioning of sympatric small mammals in an African forest-grassland vegetation mosaic. Austral. Ecol. 2013, 38, 721–729. [Google Scholar] [CrossRef]
- Navarro, A.B.; Magioli, M.; Bogoni, J.A.; Moreira, M.Z.; Silveira, L.F.; Alexandrino, R.; Apolinario da Luz, D.T.; Pizo, M.A.; Silva, W.R.; Cristina de Oliveira, V.; et al. Human-modified landscapes narrow the isotopic niche of neotropical birds. Oecologia 2021, 196, 171–184. [Google Scholar] [CrossRef]
- Ribeiro, J.F.; Guaraldo, A.; Nardoto, G.B.; Santoro, G.; Vieira, E.M. Habitat type and seasonality influence the isotopic trophic niche of small mammals in a neotropical savanna. Hystrix It. J. Mamm. 2019, 30, 30–38. [Google Scholar] [CrossRef]
- Mori, E.; Ferretti, F.; Fattorini, N. Alien war: Ectoparasite load, diet and temporal niche partitioning in a multi-species assembly of small rodents. Biol. Invasions 2019, 21, 3305–3318. [Google Scholar] [CrossRef]
- Hope, A.G.; Gragg, S.F.; Nippert, J.B.; Combe, F.J. Consumer roles of small mammals within fragmented native tallgrass prairie. Ecosphere 2021, 12, e03441. [Google Scholar] [CrossRef]
- Eckrich, C.A.; Flaherty, E.A.; Ben-David, M. Functional and numerical responses of shrews to competition vary with mouse density. PLoS ONE 2018, 13, e0189471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, C.; Guo, Q.; Zhang, Z.; Li, X.; Man, D.; Yuan, S.; Fu, H.; Wu, X.; Jin, G.; Liu, J.; et al. Trophic niche of Brandt’s voles (Lasiopodomys brandtii) and their interspecific relationships with other common rodents in a typical steppe, Inner Mongolia. Acta Theriol. Sinica 2020, 40, 424. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat | TE | TS | NSM | NH | Number of Habitats Inhabited by | ||||
|---|---|---|---|---|---|---|---|---|---|
| Af | Aa | Mm | Af + Aa | All | |||||
| Apple orchards | 11,268 | 46 | 567 | 10 | 10 | 8 | 5 | 8 | 4 |
| Plum orchards | 600 | 8 | 26 | 2 | 2 | 1 | 0 | 1 | 0 |
| Raspberry plantations | 1450 | 15 | 76 | 3 | 3 | 3 | 1 | 2 | 1 |
| Currant plantations | 2950 | 14 | 180 | 3 | 2 | 2 | 0 | 2 | 0 |
| Control habitats | 8260 | 82 | 532 | 17 | 11 | 13 | 3 | 9 | 3 |
| Habitat | Apodemus flavicollis | Apodemus agrarius | Micromys minutus | ||||||
|---|---|---|---|---|---|---|---|---|---|
| N | m/f | a/s/j | N | m/f | a/s/j | N | m/f | a/s/j | |
| Apple orchards | 127 | 67/60 | 56/44/27 | 44 | 26/18 | 11/15/18 | 5 | 2/3 | 0/0/5 |
| Plum orchards | 4 | 1/3 | 1/0/3 | 5 | 3/2 | 0/0/5 | 0 | ||
| Raspberry plantations | 19 | 13/6 | 3/10/6 | 26 | 16/10 | 9/2/15 | 1 | 1/0 | 0/0/1 |
| Currant plantations | 2 | 2/0 | 1/1/0 | 35 | 16/19 | 9/9/17 | |||
| Control habitats | 82 | 41/41 | 21/26/35 | 114 | 62/52 | 26/34/54 | 5 | 2/3 | 2/2/1 |
| Habitat | TS | NAf | NAa | NMm | NAf + Aa | NAf + Mm | NAa + Mm | NAll |
|---|---|---|---|---|---|---|---|---|
| Apple orchards | 46 | 34 | 14 | 5 | 13 | 4 | 4 | 4 |
| Plum orchards | 8 | 3 | 1 | 1 | ||||
| Raspberry plantations | 15 | 5 | 5 | 1 | 3 | 1 | 1 | 1 |
| Currant plantations | 14 | 3 | 4 | 3 | ||||
| Control habitats | 82 | 32 | 26 | 5 | 14 | 4 | 5 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balčiauskas, L.; Skipitytė, R.; Garbaras, A.; Stirkė, V.; Balčiauskienė, L.; Remeikis, V. Isotopic Niche of Syntopic Granivores in Commercial Orchards and Meadows. Animals 2021, 11, 2375. https://doi.org/10.3390/ani11082375
Balčiauskas L, Skipitytė R, Garbaras A, Stirkė V, Balčiauskienė L, Remeikis V. Isotopic Niche of Syntopic Granivores in Commercial Orchards and Meadows. Animals. 2021; 11(8):2375. https://doi.org/10.3390/ani11082375
Chicago/Turabian StyleBalčiauskas, Linas, Raminta Skipitytė, Andrius Garbaras, Vitalijus Stirkė, Laima Balčiauskienė, and Vidmantas Remeikis. 2021. "Isotopic Niche of Syntopic Granivores in Commercial Orchards and Meadows" Animals 11, no. 8: 2375. https://doi.org/10.3390/ani11082375
APA StyleBalčiauskas, L., Skipitytė, R., Garbaras, A., Stirkė, V., Balčiauskienė, L., & Remeikis, V. (2021). Isotopic Niche of Syntopic Granivores in Commercial Orchards and Meadows. Animals, 11(8), 2375. https://doi.org/10.3390/ani11082375