
Annual Long-Distance Migration Strategies and Home Range of Chinese Sparrowhawk (Accipiter soloensis) from South China


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Capture of Individuals and Transmitter Attachment
2.2. Identifying Stop-Over Sites
2.3. Defining Migration Parameters
2.4. Statistical Analysis
3. Results
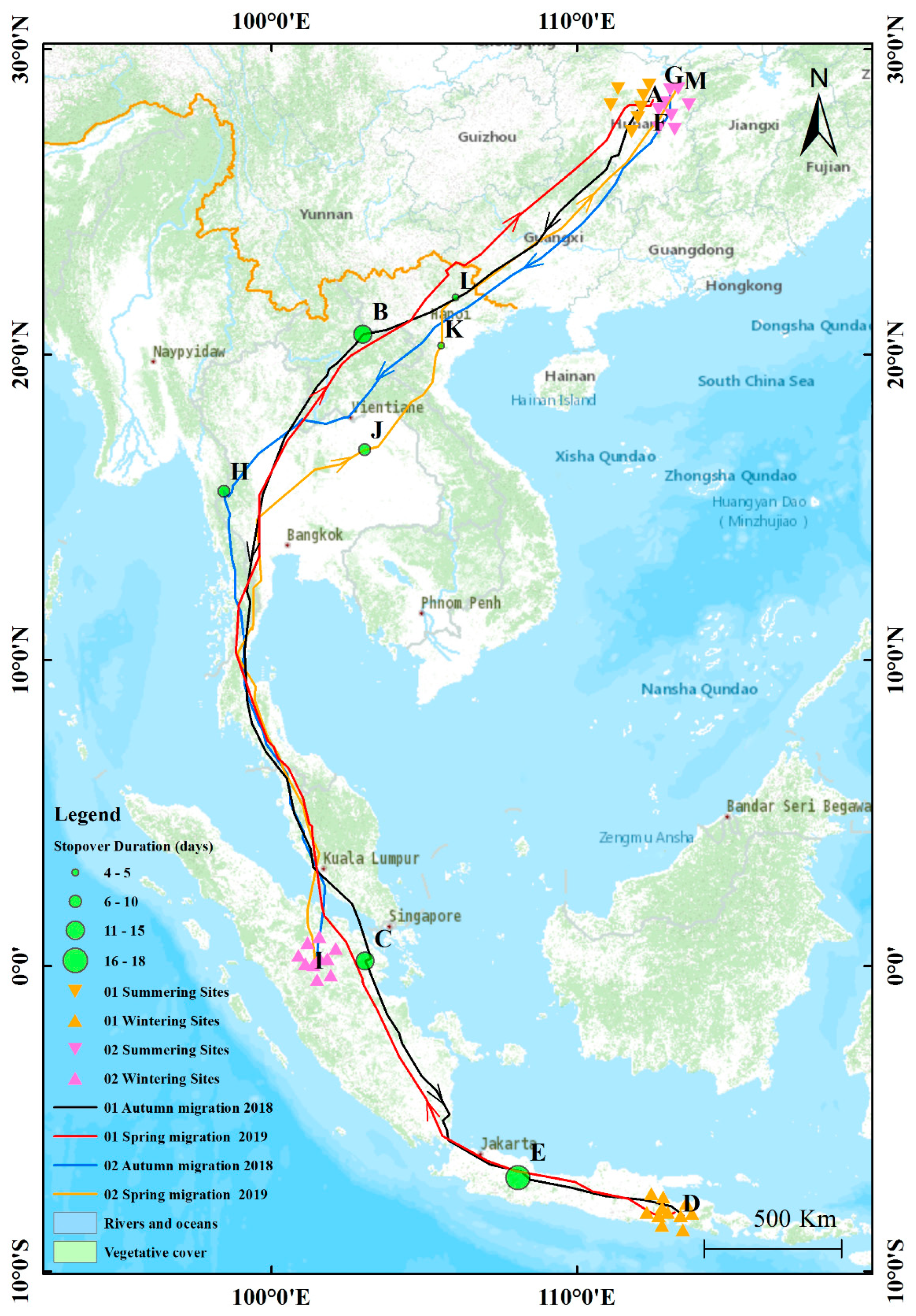
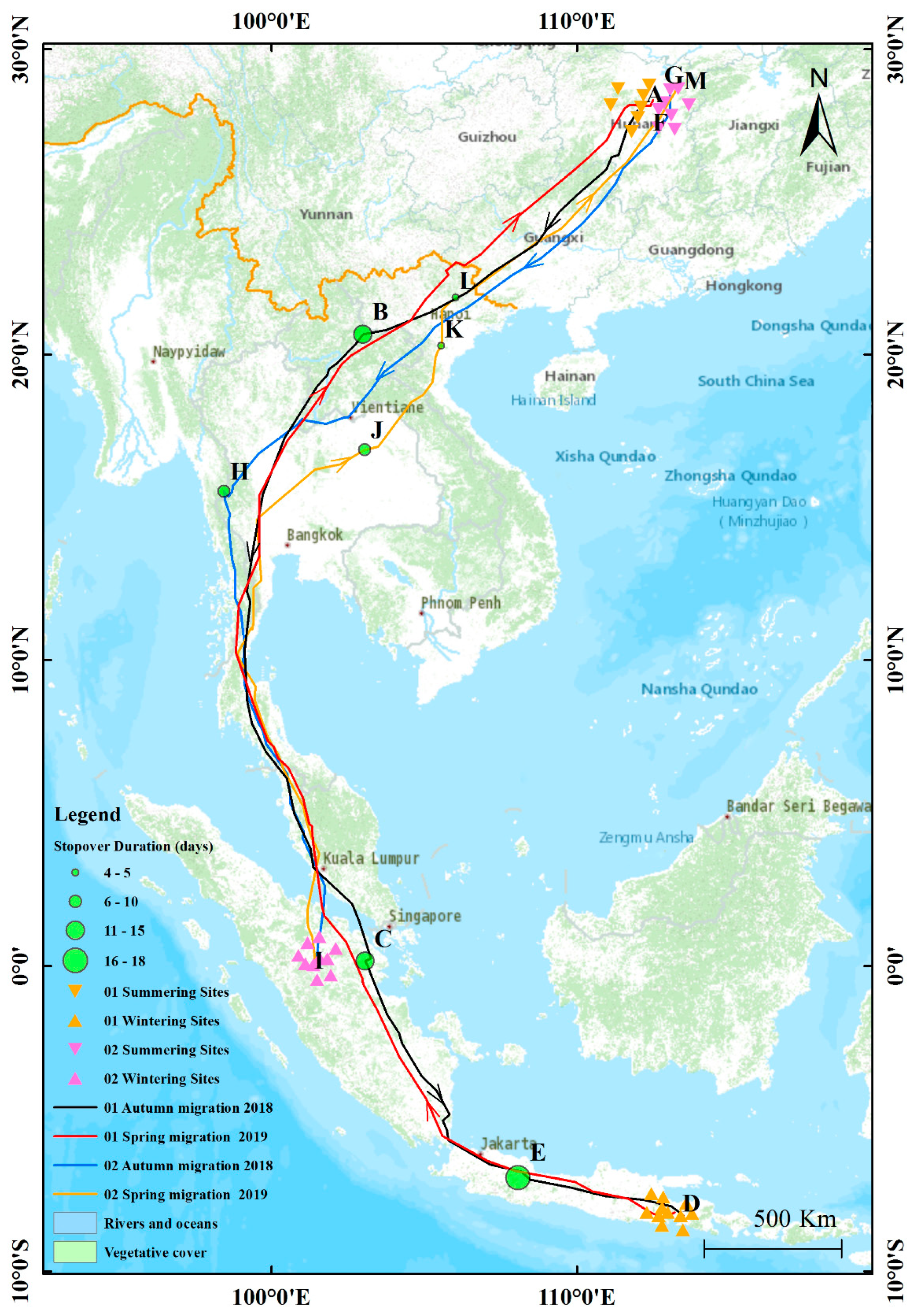
3.1. Migration Routes
3.2. Summering Sites, Wintering Sites and Stop-Over Sites
3.3. Migration Strategies
3.4. Home Range
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yamaç, E.; Bilgin, C.C. Post-fledging movements of Cinereous Vultures Aegypius monachus in Turkey revealed by GPS telemetry. Ardea 2012, 100, 149–156. [Google Scholar] [CrossRef]
- Germi, F.; Waluyo, D. Additional information on the autumn migration of raptors in east Bali, Indonesia. FORKTAIL 2006, 22, 71–76. [Google Scholar]
- Ferguson-Lees, J.; Christie, D.A. Raptors of the World; Houghton Mifflin Harcourt: Boston, MA, USA, 2001. [Google Scholar]
- Krone, O.; Berger, A.; Schulte, R. Recording movement and activity pattern of a White-tailed Sea Eagle (Haliaeetus albicilla) by a GPS datalogger. J. Ornithol. 2009, 150, 273–280. [Google Scholar] [CrossRef]
- Wattel, J. Geographical Differentiation in the Genus Accipiter; Nuttall Ornithological Club: Cambridge, MA, USA, 1973. [Google Scholar]
- White, C.M.N. Migration of Palearctic non-passerine birds in Wallacea. EMU 1976, 76, 79–82. [Google Scholar] [CrossRef]
- Beehler, B.M.; Pratt, T.K.; Zimmerman, D.A. Birds of New Guinea; Princeton University Press: Princeton, NJ, USA, 1986. [Google Scholar]
- Germi, F. Raptor migration in East Bali, Indonesia: Observations from a bottleneck watch site. Forktail 2005, 21, 93–98. [Google Scholar]
- Martín, B.; Torralvo, C.A.; Elias, G.; Tomás, J.; Onrubia1, A.; Ferrer, M. Are Western European ospreys (Pandion haliaetus) shortening their migration distances? Evidence from trends of the wintering population in the Iberian Peninsula. Eur. J. Wildl. Res. 2019, 65, 72. [Google Scholar] [CrossRef]
- Sun, Y.-H.; Deng, T.-W.; Lan, C.-Y.; Chen, C.-C. Spring Migration of Chinese Goshawks (Accipiter soloensis) in Taiwan. J. Raptor Res. 2010, 44, 188–195. [Google Scholar] [CrossRef]
- Seegar, W.S.; Cutchis, P.N.; Fuller, M.R. Fifteen years of satellite tracking developmentand application towildlife research and conservation. Johns Hopkins APL Tech. Dig. 1996, 17, 401–411. [Google Scholar]
- Moore, F.R.; Smith, R.J.; Sandberg, R. Stopover ecology of intercontinental migrants. In Birds of Two Worlds: The Ecology and Evolution of Migration; Greenberg, R., Marra, P.P., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2005; pp. 251–261. [Google Scholar]
- Gradimir, G.G.; Victor, G. Matarranz, Evgeniya, H. Dobreva, Dimitar, V. Popov, Ivelin, I. Ivanov, Ivailo, I. Klisurov, Elena, V. Kmetova, Dimitris, P. Vasilakis. First Result of the Tracking of an Eastern Imperial Eagle (Aquila heliaca) Tagged with a GPS/GSM Transmitter in Bulgaria. Acta Zool. Bulg. 2011, 3, 15–20. [Google Scholar]
- Gu, Z.; Pan, S.; Lin, Z.; Hu, L.; Dal, X.; Chang, J.; Xue, Y.; Su, H.; Long, J.; Sun, M.; et al. Climate-driven flyway changes and memory-based long-distance migration. Nature 2021, 591, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hahn, S.; Emmenegger, T.; Lisovski, S.; Amrhein, V.; Zehtindjiev, P.; Liechti, F. Variable detours in long-distance migration across ecological barriers and their relation to habitat availability at ground. Ecol. Evol. 2014, 4, 4150–4160. [Google Scholar] [CrossRef] [PubMed]
- Van Der Winden, J.; Poot, M.J.M.; Van Horssen, P.W. Large birds can migrate fast: The post-breeding flight of the Purple Heron Ardea purpurea to the Sahel. Ardea 2010, 98, 395–402. [Google Scholar] [CrossRef]
- Ye, X.; Xu, Z.; Aharon-Rotman, Y.; Yu, H.; Cao, L. First Description of Grey Heron Ardea cinerea Migration Recorded by GPS/GSM Transmitter. Ornithol. Sci. 2018, 17, 223–228. [Google Scholar] [CrossRef]
- Beekman, J.H.; Nolet, B.A.; Klaassen, M. Skipping swans: Fuelling rates and wind conditions determine differential use of migratory stopover sites of Bewick’s swans Cygnus bewickii. Ardea 2002, 90, 437–460. [Google Scholar]
- Gudmundsson, G.A.; Lindström, A.; Alerstam, T. Optimal fat loads and long-distance flight by migrating Knots Calidris canutus, Sanderlings, C. alba and Turnstones Arenaria interpres. Ibis 1991, 133, 140–152. [Google Scholar] [CrossRef]
- Henny, C.J.; Martell, M.S. Satellite-tagged Osprey Nearly Sets Longevity Record and Productivity Response to Initial Captures. J. Raptor Res. 2017, 51, 180–183. [Google Scholar] [CrossRef]
- Klaassen, R.H.G.; Ens, B.J.; Shamoun-Baranes, J.; Exo, K.M.; Bairlein, F. Migration strategy of a flight generalist, the Lesser Black-backed Gull Larus fuscus. Behav. Ecol. 2012, 23, 58–68. [Google Scholar] [CrossRef] [Green Version]
- Heim, W.; Pedersen, L.; Heim, R.; Kamp, J.; Smirenski, S.M.; Thomas, A.; Tøttrup, A.P.; Thorup, K. Full annual cycle tracking of a small songbird, the Siberian Rubythroat Calliope calliope, along the East Asian flyway. J. Ornithol. 2018, 159, 893–899. [Google Scholar] [CrossRef]
- Väli, Ü.; Bergmanis, U. Apparent survival rates of adult Lesser Spotted Eagle Clanga pomarina estimated by GPS-tracking, colour rings and wing-tags. Bird Study 2017, 64, 104–107. [Google Scholar] [CrossRef]
- Paganini, S.T.; Stafford, A.; von Hirschheydt, J.; Kéry, M. A Large Aggregation of 50 Peregrine Falcons (Falco peregrinus) During Migration in the Western Gulf of Mexico. J. Raptor Res. 2018, 52, 500–502. [Google Scholar] [CrossRef]
- Väli, Ü. Timing of breeding events of the Lesser Spotted Eagle Clanga pomarina as revealed by remote cameras and GPS-tracking. ARDEA 2017, 105, 77–81. [Google Scholar] [CrossRef]
- Sørensen, I.H.; Schlaich, A.E.; Klaassen, R.H.G.; Heldbjerg, H.; Koks, B.J. Rare case of an adult male Montagu’s Harrier Circus pygargus over-summering in West Africa, as revealed by GPS tracking. J. Ornithol. 2017, 158, 753–760. [Google Scholar] [CrossRef]
- Massa, C.; Gabelli, F.M.; Cueto, G.R. Using GPS tracking to determine movement patterns and foraging habitat selection of the Common Barn-owl (Tyto alba). Horner 2015, 30, 7–12. [Google Scholar]
- Kelly, L. A Critical Rew on Home Range Studies. J. Wildl. Manag. 2008, 72, 290–298. [Google Scholar]
- García-Ripollés, C.; López-López, P.; Urios, V. First description of migration and wintering of adult Egyptian Vultures Neophron percnopterus tracked by GPS satellite telemetry. Bird Study 2010, 57, 261–265. [Google Scholar] [CrossRef] [Green Version]
- Gustin, M.; Giglio, G.; Pellegrino, S.C.; Frassanito, A.; Ferrarini, A. Space use and flight attributes of breeding Lesser Kestrels Falco naumanni revealed by GPS tracking. Bird Study 2017, 64, 274–277. [Google Scholar] [CrossRef]
- Germi, F.; Salim, A.; Minganti, A. First records of Chinese Sparrowhawk Accipiter soloensis wintering in Papua (Indonesian New Guinea). FORKTAIL 2013, 29, 43–47. [Google Scholar]
- Eichhorn, G.; Drent, R.H.; Stahl, J.; Leito, A.; Alerstam, T. Skipping the Baltic: The emergence of a dichotomy of alternative spring migration strategies in Russian barnacle geese. J. Anim. Ecol. 2009, 78, 63–72. [Google Scholar] [CrossRef]
- Henningsson, S.S.; Alerstam, T. Barriers and distances as determinants for the evolution of bird migration links: The arctic shorebird system. Proc. R. Soc. B Biol. Sci. 2005, 272, 2251–2258. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, C.; Klaassen, R.H.G.; Alerstam, T. Differences in speed and duration of bird migration between spring and autumn. Am. Nat. 2013, 181, 837–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krüger, S.; Amar, A. Insights into post-fledging dispersal of Bearded Vultures Gypaetus barbatus in southern Africa from GPS satellite telemetry. Bird Study 2017, 64, 125–131. [Google Scholar] [CrossRef]
- Burt, W.H. Territoriality and home range as applied to mammals. J. Mammal. 1943, 24, 346–352. [Google Scholar] [CrossRef]
- Powell, R.A. Animal Home Ranges and Territories and Home Range Estimators. In Research Techniques in Animal Ecology: Controversies and Consequences; Luiqi, B., Todd, K.F., Eds.; Columbia University Press: New York, NY, USA, 2000; pp. 65–110. [Google Scholar]
- Patrick, E.M. Site Fidelity, Home Range, and Daily Movements of White Perch, Morone Americana, and Striped Bass, Morone Saxatilis, in Two Small Tributaries of the York River, Virginia. Ph.D. Thesis, The College of William and Mary, Williamsburg, VA, USA, 2005; pp. 16–20. [Google Scholar]
- Bengtsson, D.; Avril, A.; Gunnarsson, G.; Elmberg, J.; Söderquist, P.; Norevik, G. Movements, home range size and habitat selection of mallards during autmn migration. PLoS ONE 2014, 9, e100764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Christie, M.; Coleman, J.; Hassell, C.; Gosbell, K.; Lisovski, S.; Minton, C.; Klaassen, M. Time versus energy minimization migration strategy varies with body size and season in long-distance migratory shorebirds. Mov. Ecol. 2017, 5, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Bird Number | Bird 01 HQP589 21.05.2018 | Bird 02 HQP593 22.05.2018 | ||
|---|---|---|---|---|
| Equipment Serial Number | ||||
| Release Date | ||||
| Autumn Migration | Spring Migration | Autumn Migration | Springmigration | |
| Start Date | 02.09.2018 | 02.03.2019 | 03.09.2018 | 13.03.2019 |
| Ending Date | 24.11.2018 | 22.05.2019 | 23.10.2018 | 30.04.2019 |
| Days of Migration (Days) | 84 | 83 | 50 | 49 |
| Number of GPS Sites | 75 | 83 | 102 | 102 |
| Migration Distance (km) | 5550 | 5379 | 4110 | 4061 |
| Linear Distance (km) | 4103 | 4185 | 3470 | 3493 |
| Minimum and Maximum Daily Distance (km) | 36–212 | 32–252 | 18–198 | 16–262 |
| Number of Stops | 2 | 1 | 1 | 3 |
| Stop-Over Duration (Days) | 14, 15 | 18 | 9 | 10, 4, 5 |
| Bird Number | Type of Sites | Site | Longitude and Latitude | Location | Country | Arrival–Departure |
|---|---|---|---|---|---|---|
| Bird 01 | Summering sites | A | 112°37′9″ E, 28°9′51″ N | Ningxiang | China | 28.05.2018–02.09.2018 |
| Autumn migration stop-over sites | B | 102°59′29″ E, 20°39′58″ N | Oudomxay | Laos | 16.09.2018–29.09.2018 | |
| C | 103°3′19″ E, 0°9′3″ N | Riau | Indonesia | 21.10.2018–04.11.2018 | ||
| Wintering sites | D | 113°29′29″ E, 8°10′17″ S | Lawang | Indonesia | 24.11.2018–01.03.2019 | |
| Spring migration stop-over sites | E | 112°49′16″ E, 8°14′57″ S | Malang | Indonesia | 02.03.2019–19.03.2019 | |
| Summering sites | F | 112°42′22″ E, 28°11′24″ N | Ningxiang | China | 22.05.2019–08.09.2019 | |
| Bird 02 | Summering sites | G | 113°1′54″ E, 28°31′4″ N | Changsha | China | 28.05.2018–03.09.2018 |
| Autumn migration stop-over sites | H | 98°26′7″ E, 15°31′54″ N | Thaton | Myanmar | 27.09.2018–05.10.2018 | |
| Wintering sites | I | 101°25′19″ E, 0°1′2″ S | Sumatra | Indonesia | 23.10.2018–13.03.2019 | |
| Spring migration stop-over sites | J | 103°2′12″ E, 16°52′44″ N | Non Sung | Thailand | 30.03.2019–08.04.2019 | |
| K | 105°35′26″ E, 20°33′23″ N | Hanoi | Vietnam | 14.04.2019–17.04.2019 | ||
| L | 105°36′3″ E, 21°29′29″ N | Quang Minh | Vietnam | 18.04.2019–22.04.2019 | ||
| Summering sites | M | 113°15′26″ E, 28°42′31″ N | Wushi | China | 01.05.2019–01.09.2019 |
| Bird Number | Season | Type of Site | Locations | 100% Minimum Convex Polygon (km2) | Fixed Kernel (km2) | ||
|---|---|---|---|---|---|---|---|
| 95% | 75% | 50% or Core Area | |||||
| Bird 01 | Autumn migration 2018 | Summering sites | A | 2617.2565 | 10.9326 | 3.6414 | 1.4716 |
| Stop-over sites | B | 0.0592 | 0.0563 | 0.0171 | 0.0032 | ||
| C | 0.1608 | 0.0867 | 0.0396 | 0.0196 | |||
| Wintering sites | D | 9.7193 | 1.0919 | 0.3925 | 0.1680 | ||
| Spring migration 2019 | Stop-over sites | E | 0.3090 | 0.0096 | 0.0021 | 0.0006 | |
| Summering sites | F | 6730.9273 | 36.5494 | 14.5752 | 7.7079 | ||
| Bird 02 | Autumn migration 2018 | Summering sites | G | 468.4247 | 3.3148 | 0.9811 | 0.3747 |
| Stop-over sites | H | 0.2338 | 0.1678 | 0.0473 | 0.0128 | ||
| Wintering sites | I | 7.8153 | 2.0470 | 0.8079 | 0.3892 | ||
| Spring migration 2019 | Stop-over sites | J | 0.3535 | 0.0298 | 0.0081 | 0.0038 | |
| K | 0.0440 | 0.0847 | 0.0447 | 0.0202 | |||
| L | 0.4575 | 0.0117 | 0.0026 | 0.0010 | |||
| Summering sites | M | 415.2083 | 28.9693 | 8.5827 | 2.6349 | ||
| n | 100% Minimum Convex Polygon (km2) | Fixed Kernel (km2) | ||||
|---|---|---|---|---|---|---|
| 95% | 75% | 50% or Core Area | ||||
| Summering sites | Mean ± SD | 2557.9543 ± 2965.0585 | 19.9415 ± 15.4373 | 6.9451 ± 5.9829 | 3.0473 ± 3.2412 | |
| 4 | Max | 6730.9275 | 36.5494 | 14.5752 | 7.7079 | |
| Min | 415.2083 | 3.3148 | 0.9811 | 0.3747 | ||
| Wintering sites | Mean ± SD | 8.7673 ± 1.3463 | 1.5695 ± 0.6754 | 0.6002 ± 0.2937 | 0.2786 ± 0.1564 | |
| 2 | Max | 9.7193 | 2.0470 | 0.8079 | 0.3892 | |
| Min | 7.8153 | 1.0919 | 0.3925 | 0.1680 | ||
| Stop-over sites | Mean ± SD | 0.2311 ± 0.1537 | 0.0638 ± 0.0557 | 0.0231 ± 0.0202 | 0.0087 ± 0.0086 | |
| 7 | Max | 0.4575 | 0.1678 | 0.0473 | 0.0202 | |
| Min | 0.0440 | 0.0096 | 0.0021 | 0.0006 | ||
| Autumn migration stop-over sites | Mean ± SD | 0.1513 ± 0.0877 | 0.1036 ± 0.0576 | 0.0347 ± 0.0157 | 0.0119 ± 0.0082 | |
| 3 | Max | 0.2338 | 0.1678 | 0.0473 | 0.0196 | |
| Min | 0.0592 | 0.0563 | 0.0171 | 0.0032 | ||
| Spring migration Stop-over sites | Mean ± SD | 0.2910 ± 0.1760 | 0.0340 ± 0.0350 | 0.0144 ± 0.0204 | 0.0064 ± 0.0093 | |
| 4 | Max | 0.4575 | 0.0847 | 0.0447 | 0.0202 | |
| Min | 0.0440 | 0.0096 | 0.0021 | 0.0006 | ||
| Between the three type sites | 13 | p | 0.007 | 0.007 | 0.007 | 0.009 |
| Between the autumn and spring migration stop-over sites | 7 | p | 0.289 | 0.077 | 0.157 | 0.480 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Min, X.; Gao, Z.; Lin, Y.; Lu, C.-H. Annual Long-Distance Migration Strategies and Home Range of Chinese Sparrowhawk (Accipiter soloensis) from South China. Animals 2021, 11, 2237. https://doi.org/10.3390/ani11082237
Min X, Gao Z, Lin Y, Lu C-H. Annual Long-Distance Migration Strategies and Home Range of Chinese Sparrowhawk (Accipiter soloensis) from South China. Animals. 2021; 11(8):2237. https://doi.org/10.3390/ani11082237
Chicago/Turabian StyleMin, Xiao, Zijing Gao, Yuanfeng Lin, and Chang-Hu Lu. 2021. "Annual Long-Distance Migration Strategies and Home Range of Chinese Sparrowhawk (Accipiter soloensis) from South China" Animals 11, no. 8: 2237. https://doi.org/10.3390/ani11082237
APA StyleMin, X., Gao, Z., Lin, Y., & Lu, C.-H. (2021). Annual Long-Distance Migration Strategies and Home Range of Chinese Sparrowhawk (Accipiter soloensis) from South China. Animals, 11(8), 2237. https://doi.org/10.3390/ani11082237