
Maternal Nutrition and Developmental Programming of Male Progeny


Simple Summary
Abstract
1. Introduction
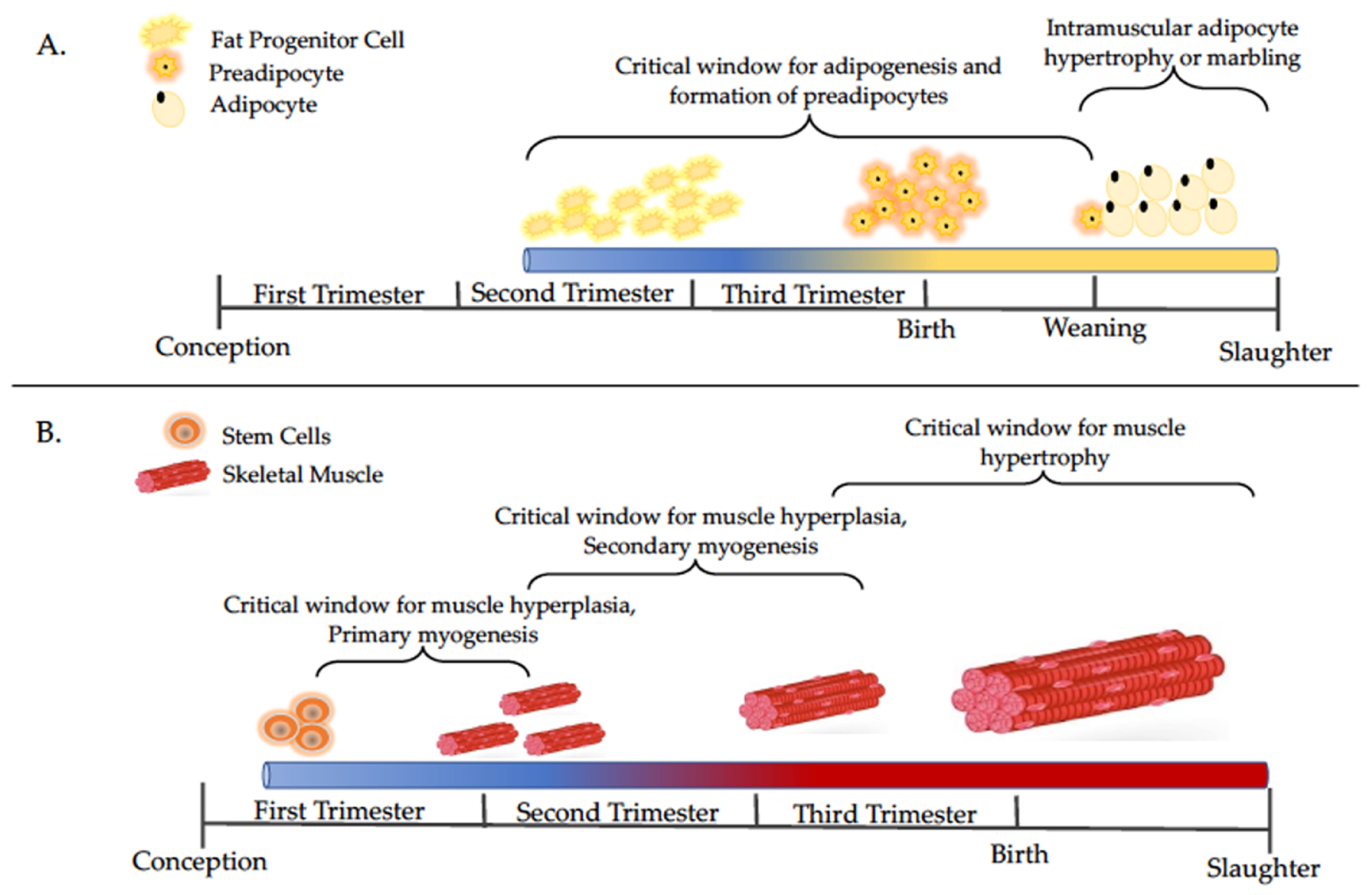
2. Maternal Under- and Over-nutrition Impacts on Skeletal Muscle and Adipose Tissue Development
3. Maternal Dietary Energy
3.1. Maternal Dietary Energy and Developmental Programming
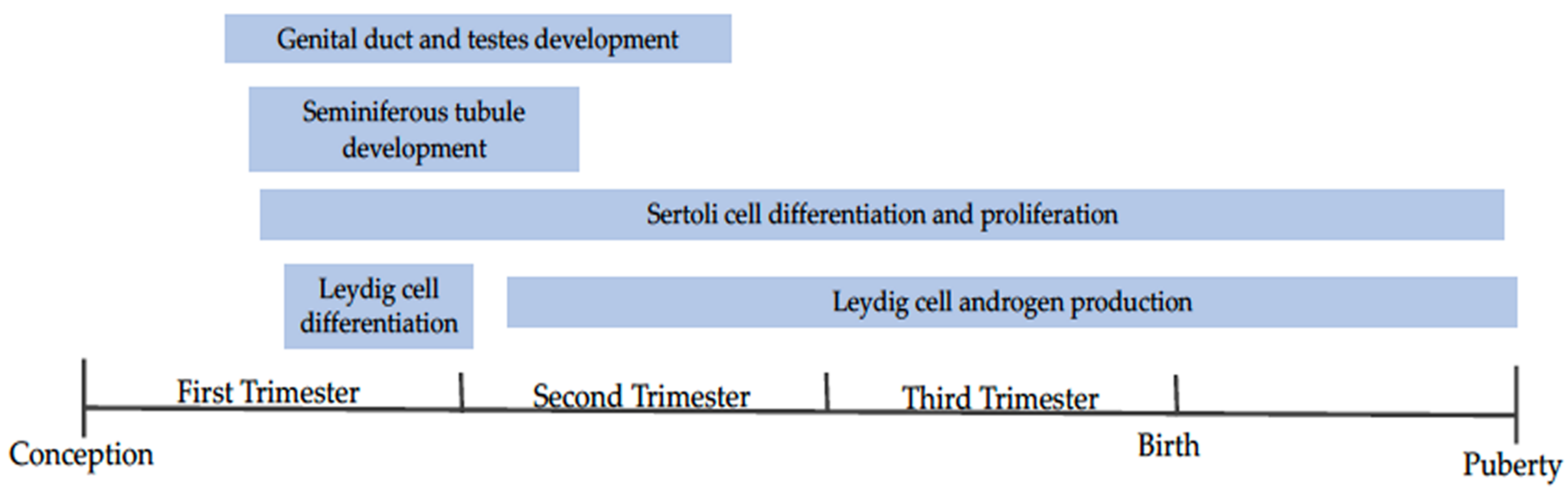
3.2. Dietary Energy In Utero and Male Reproductive Capacity
4. Maternal Dietary Protein
4.1. Dietary Protein and Developmental Programming
4.2. Dietary Protein In Utero and Male Reproductive Capacity
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Barker, D.J.; Clark, P.M. Fetal undernutrition and disease in later life. Rev. Reprod. 1997, 2, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Wallace, J.M.; Spencer, T.E. Board-invited review: Intrauterine growth retardation: Implications for the animal sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef]
- Bell, A.W. Prenatal programming of postnatal productivity and health of livestock: A brief review. Aust. J. Exp. Agric. 2006, 46, 725–732. [Google Scholar] [CrossRef]
- Funston, R.N.; Larson, D.M.; Vonnahme, K.A. Effects of maternal nutrition on conceptus growth and offspring performance: Implications for beef cattle production. J. Anim. Sci. 2010, 88, E205–E215. [Google Scholar] [CrossRef]
- Caton, J.S.; Crouse, M.S.; McLean, K.J.; Dahlen, C.R.; Ward, A.K.; Cushman, R.A.; Grazul-Bilska, A.T.; Neville, B.W.; Borowicz, P.P.; Reynolds, L.P. Maternal periconceptual nutrition, early pregnancy, and developmental outcomes in beef cattle. J. Anim. Sci. 2020, 98. [Google Scholar] [CrossRef] [PubMed]
- National Academies of Sciences, Engineering, and Medicine. Nutrient Requirements of Beef Cattle: Eighth Revised Edition; The National Academies Press: Washington, DC, USA, 2016; p. 494. [Google Scholar]
- Greenwood, P.; Clayton, E.; Bell, A. Developmental programming and beef production. Anim. Front. 2017, 7, 38–47. [Google Scholar] [CrossRef][Green Version]
- Micke, G.C.; Sullivan, T.M.; McMillen, I.C.; Gentili, S.; Perry, V.E. Heifer nutrient intake during early- and mid-gestation programs adult offspring adiposity and mRNA expression of growth-related genes in adipose depots. Reproduction 2011, 141, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Micke, G.C.; Sullivan, T.M.; Kennaway, D.J.; Hernandez-Medrano, J.; Perry, V.E. Maternal endocrine adaptation throughout pregnancy to nutrient manipulation: Consequences for sexually dimorphic programming of thyroid hormones and development of their progeny. Theriogenology 2015, 83, 604–615. [Google Scholar] [CrossRef]
- Copping, K.J.; Hoare, A.; Callaghan, M.; McMillen, I.C.; Rodgers, R.J.; Perry, V.E.A. Fetal programming in 2-year-old calving heifers: Peri-conception and first trimester protein restriction alters fetal growth in a gender-specific manner. Anim. Prod. Sci. 2014, 54, 1333–1337. [Google Scholar] [CrossRef]
- Hinde, K.; Carpenter, A.J.; Clay, J.S.; Bradford, B.J. Holsteins favor heifers, not bulls: Biased milk production programmed during pregnancy as a function of fetal sex. PLoS ONE 2014, 9, e86169. [Google Scholar] [CrossRef]
- Trivers, R.L.; Willard, D.E. Natural selection of parental ability to vary the sex ratio of offspring. Science 1973, 179, 90–92. [Google Scholar] [CrossRef]
- Zhu, M.J.; Ford, S.P.; Nathanielsz, P.W.; Du, M. Effect of maternal nutrient restriction in sheep on the development of fetal skeletal muscle. Biol. Reprod. 2004, 71, 1968–1973. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Tong, J.; Zhao, J.; Underwood, K.R.; Zhu, M.; Ford, S.P.; Nathanielsz, P.W. Fetal programming of skeletal muscle development in ruminant animals. J. Anim. Sci. 2010, 88, E51–E60. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhu, M.J.; Dodson, M.V.; Du, M. Developmental programming of fetal skeletal muscle and adipose tissue development. J. Genom. 2013, 1, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Zhu, M.J.; Underwood, K.R.; Hess, B.W.; Ford, S.P.; Du, M. AMP-activated protein kinase and adipogenesis in sheep fetal skeletal muscle and 3T3-L1 cells1. J. Anim. Sci. 2008, 86, 1296–1305. [Google Scholar] [CrossRef]
- Wang, Q.A.; Tao, C.; Gupta, R.K.; Scherer, P.E. Tracking adipogenesis during white adipose tissue development, expansion and regeneration. Nat. Med. 2013, 19, 1338–1344. [Google Scholar] [CrossRef]
- Du, M.; Zhao, J.X.; Yan, X.; Huang, Y.; Nicodemus, L.V.; Yue, W.; McCormick, R.J.; Zhu, M.J. Fetal muscle development, mesenchymal multipotent cell differentiation, and associated signaling pathways. J. Anim. Sci. 2011, 89, 583–590. [Google Scholar] [CrossRef]
- Blasio, M.J.D.; Gatford, K.L.; Robinson, J.S.; Owens, J.A. Placental restriction of fetal growth reduces size at birth and alters postnatal growth, feeding activity, and adiposity in the young lamb. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2007, 292, R875–R886. [Google Scholar] [CrossRef]
- Desai, M.; Crowther, N.J.; Lucas, A.; Hales, C.N. Organ-selective growth in the offspring of protein-restricted mothers. Br. J. Nutr. 1996, 76, 591–603. [Google Scholar] [CrossRef]
- Ramirez, M.; Testa, L.M.; Lopez Valiente, S.; Latorre, M.E.; Long, N.M.; Rodriguez, A.M.; Pavan, E.; Maresca, S. Maternal energy status during late gestation: Effects on growth performance, carcass characteristics and meat quality of steers progeny. Meat. Sci. 2020, 164, 108095. [Google Scholar] [CrossRef]
- Ford, S.P.; Zhang, L.; Zhu, M.; Miller, M.M.; Smith, D.T.; Hess, B.W.; Moss, G.E.; Nathanielsz, P.W.; Nijland, M.J. Maternal obesity accelerates fetal pancreatic beta-cell but not alpha-cell development in sheep: Prenatal consequences. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R835–R843. [Google Scholar] [CrossRef] [PubMed]
- George, L.A.; Uthlaut, A.B.; Long, N.M.; Zhang, L.; Ma, Y.; Smith, D.T.; Nathanielsz, P.W.; Ford, S.P. Different levels of overnutrition and weight gain during pregnancy have differential effects on fetal growth and organ development. Reprod. Biol. Endocrinol. 2010, 8, 75. [Google Scholar] [CrossRef]
- Sandoval, C.; Askelson, K.; Lambo, C.A.; Dunlap, K.A.; Satterfield, M.C. Effect of maternal nutrient restriction on expression of glucose transporters (SLC2A4 and SLC2A1) and insulin signaling in skeletal muscle of SGA and Non-SGA sheep fetuses. Domest. Anim. Endocrinol. 2021, 74, 106556. [Google Scholar] [CrossRef]
- Sandoval, C.; Lambo, C.A.; Beason, K.; Dunlap, K.A.; Satterfield, M.C. Effect of maternal nutrient restriction on skeletal muscle mass and associated molecular pathways in SGA and Non-SGA sheep fetuses. Domest. Anim. Endocrinol. 2020, 72, 106443. [Google Scholar] [CrossRef]
- Ford, S.P.; Hess, B.W.; Schwope, M.M.; Nijland, M.J.; Gilbert, J.S.; Vonnahme, K.A.; Means, W.J.; Han, H.; Nathanielsz, P.W. Maternal undernutrition during early to mid-gestation in the ewe results in altered growth, adiposity, and glucose tolerance in male offspring. J. Anim. Sci. 2007, 85, 1285–1294. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.S.; Lang, A.L.; Grant, A.R.; Nijland, M.J. Maternal nutrient restriction in sheep: Hypertension and decreased nephron number in offspring at 9 months of age. J. Physiol. 2005, 565, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Long, N.M.; Rule, D.C.; Zhu, M.J.; Nathanielsz, P.W.; Ford, S.P. Maternal obesity upregulates fatty acid and glucose transporters and increases expression of enzymes mediating fatty acid biosynthesis in fetal adipose tissue depots. J. Anim. Sci. 2012, 90, 2201–2210. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, J.M.; Camacho, L.E.; Ebarb, S.M.; Swanson, K.C.; Vonnahme, K.A.; Stelzleni, A.M.; Johnson, S.E. Realimentation of nutrient restricted pregnant beef cows supports compensatory fetal muscle growth. J. Anim. Sci. 2013, 91, 4797–4806. [Google Scholar] [CrossRef]
- Duarte, M.S.; Gionbelli, M.P.; Paulino, P.V.; Serao, N.V.; Nascimento, C.S.; Botelho, M.E.; Martins, T.S.; Filho, S.C.; Dodson, M.V.; Guimaraes, S.E.; et al. Maternal overnutrition enhances mRNA expression of adipogenic markers and collagen deposition in skeletal muscle of beef cattle fetuses. J. Anim. Sci. 2014, 92, 3846–3854. [Google Scholar] [CrossRef]
- Moisa, S.J.; Shike, D.W.; Shoup, L.; Rodriguez-Zas, S.L.; Loor, J.J. Maternal Plane of Nutrition during Late Gestation and Weaning Age Alter Angus x Simmental Offspring Longissimus Muscle Transcriptome and Intramuscular Fat. PLoS ONE 2015, 10, e0131478. [Google Scholar] [CrossRef] [PubMed]
- Paradis, F.; Wood, K.M.; Swanson, K.C.; Miller, S.P.; McBride, B.W.; Fitzsimmons, C. Maternal nutrient restriction in mid-to-late gestation influences fetal mRNA expression in muscle tissues in beef cattle. BMC Genom. 2017, 18, 632. [Google Scholar] [CrossRef] [PubMed]
- Hewison, A.J.; Gaillard, J.M. Successful sons or advantaged daughters? The Trivers-Willard model and sex-biased maternal investment in ungulates. Trends Ecol. Evol. 1999, 14, 229–234. [Google Scholar] [CrossRef]
- Gionbelli, T.R.S.; Veloso, C.M.; Rotta, P.P.; Valadares Filho, S.C.; Carvalho, B.C.; Marcondes, M.I.; Cunha, S.C.; Novaes, M.A.S.; Prezotto, L.D.; Duarte, M.S.; et al. Foetal development of skeletal muscle in bovines as a function of maternal nutrition, foetal sex and gestational age. J. Anim. Physiol. Anim. Nutr. 2018, 102, 545–556. [Google Scholar] [CrossRef]
- Gionbelli, T.R.S.; Rotta, P.P.; Veloso, C.M.; Valadares Filho, S.C.; Carvalho, B.C.; Marcondes, M.I.; Ferreira, M.F.L.; Souza, J.V.F.; Santos, J.; Lacerda, L.C.; et al. Intestinal development of bovine foetuses during gestation is affected by foetal sex and maternal nutrition. J. Anim. Physiol. Anim. Nutr. 2017, 101, 493–501. [Google Scholar] [CrossRef]
- Long, N.M.; Rule, D.C.; Tuersunjiang, N.; Nathanielsz, P.W.; Ford, S.P. Maternal obesity in sheep increases fatty acid synthesis, upregulates nutrient transporters, and increases adiposity in adult male offspring after a feeding challenge. PLoS ONE 2015, 10, e0122152. [Google Scholar] [CrossRef]
- Nicholas, L.M.; Rattanatray, L.; MacLaughlin, S.M.; Ozanne, S.E.; Kleemann, D.O.; Walker, S.K.; Morrison, J.L.; Zhang, S.; Muhlhausler, B.S.; Martin-Gronert, M.S.; et al. Differential effects of maternal obesity and weight loss in the periconceptional period on the epigenetic regulation of hepatic insulin-signaling pathways in the offspring. FASEB J 2013, 27, 3786–3796. [Google Scholar] [CrossRef]
- Caton, J.S.; Crouse, M.S.; Reynolds, L.P.; Neville, T.L.; Dahlen, C.R.; Ward, A.K.; Swanson, K.C. Maternal nutrition and programming of offspring energy requirements. Transl. Anim. Sci. 2019, 3, 976–990. [Google Scholar] [CrossRef]
- Vonnahme, K.A.; Hess, B.W.; Hansen, T.R.; McCormick, R.J.; Rule, D.C.; Moss, G.E.; Murdoch, W.J.; Nijland, M.J.; Skinner, D.C.; Nathanielsz, P.W.; et al. Maternal undernutrition from early- to mid-gestation leads to growth retardation, cardiac ventricular hypertrophy, and increased liver weight in the fetal sheep. Biol. Reprod. 2003, 69, 133–140. [Google Scholar] [CrossRef]
- Wallace, L.R. The growth of lambs before and after birth in relation to the level of nutrition. J. Agric. Sci. 1948, 38, 243–302. [Google Scholar] [CrossRef]
- Baumann, M.U.; Deborde, S.; Illsley, N.P. Placental glucose transfer and fetal growth. Endocrine 2002, 19, 13–22. [Google Scholar] [CrossRef]
- Bell, A.; Greenwood, P.; Ehrhardt, R. Chapter 1 Regulation of metabolism and growth during prenatal life. Biol. Grow. Anim. 2005, 3, 3–34. [Google Scholar]
- Sanglard, L.P.; Nascimento, M.; Moriel, P.; Sommer, J.; Ashwell, M.; Poore, M.H.; Duarte, M.S.; Serao, N.V.L. Impact of energy restriction during late gestation on the muscle and blood transcriptome of beef calves after preconditioning. BMC Genom. 2018, 19, 702. [Google Scholar] [CrossRef] [PubMed]
- Corah, L.R.; Dunn, T.G.; Kaltenbach, C.C. Influence of prepartum nutrition on the reproductive performance of beef females and the performance of their progeny. J. Anim. Sci. 1975, 41, 819–824. [Google Scholar] [CrossRef]
- Loerch, S.C. Limit-feeding corn as an alternative to hay for gestating beef cows. J. Anim. Sci. 1996, 74, 1211–1216. [Google Scholar] [CrossRef]
- Radunz, A.E.; Fluharty, F.L.; Relling, A.E.; Felix, T.L.; Shoup, L.M.; Zerby, H.N.; Loerch, S.C. Prepartum dietary energy source fed to beef cows: II. Effects on progeny postnatal growth, glucose tolerance, and carcass composition. J. Anim. Sci. 2012, 90, 4962–4974. [Google Scholar] [CrossRef]
- Harmon, D.L. Impact of nutrition on pancreatic exocrine and endocrine secretion in ruminants: A review. J. Anim. Sci. 1992, 70, 1290–1301. [Google Scholar] [CrossRef]
- Gardner, D.S.; Van Bon, B.W.; Dandrea, J.; Goddard, P.J.; May, S.F.; Wilson, V.; Stephenson, T.; Symonds, M.E. Effect of periconceptional undernutrition and gender on hypothalamic-pituitary-adrenal axis function in young adult sheep. J. Endocrinol. 2006, 190, 203–212. [Google Scholar] [CrossRef]
- Radunz, A.E.; Fluharty, F.L.; Day, M.L.; Zerby, H.N.; Loerch, S.C. Prepartum dietary energy source fed to beef cows: I. Effects on pre- and postpartum cow performance. J. Anim. Sci. 2010, 88, 2717–2728. [Google Scholar] [CrossRef] [PubMed]
- Willett, E.L.; Ohms, J.I. Measurement of Testicular Size and Its Relation to Production of Spermatozoa by Bulls. J. Dairy Sci. 1957, 40, 1559–1567. [Google Scholar] [CrossRef]
- Hahn, J.; Foote, R.H.; Seidel, G.E., Jr. Testicular growth and related sperm output in dairy bulls. J. Anim. Sci. 1969, 29, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Abella, D.; Becu-Villalobos, D.; Lacau-Mengido, I.M.; Villegas, N.; Bentancur, O. Sperm production, testicular size, serum gonadotropins and testosterone levels in Merino and Corriedale breeds. Reprod. Nutr. Dev. 1999, 39, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Zamboni, L.; Bezard, J.; Mauleon, P. Role of the Mesonephros in the Development of the Sheep Fetal Ovary. Ann. Biol. Anim. Bioch. 1979, 19, 1153–1178. [Google Scholar] [CrossRef]
- Vigier, B.; Prepin, J.; Jost, A. Chronology of development of the genital tract of the calf fetus. Arch. Anat. Microsc. Morphol. Exp. 1976, 65, 77–101. [Google Scholar] [PubMed]
- Alejandro, B.; Perez, R.; Pedrana, G.; Milton, J.T.; Lopez, A.; Blackberry, M.A.; Duncombe, G.; Rodriguez-Martinez, H.; Martin, G.B. Low maternal nutrition during pregnancy reduces the number of Sertoli cells in the newborn lamb. Reprod. Fertil. Dev. 2002, 14, 333–337. [Google Scholar] [CrossRef]
- Hoffman, F.; Boretto, E.; Vitale, S.; Gonzalez, V.; Vidal, G.; Pardo, M.F.; Flores, M.F.; Garcia, F.; Bagnis, G.; Queiroz, O.C.M.; et al. Maternal nutritional restriction during late gestation impairs development of the reproductive organs in both male and female lambs. Theriogenology 2018, 108, 331–338. [Google Scholar] [CrossRef]
- Rae, M.T.; Kyle, C.E.; Miller, D.W.; Hammond, A.J.; Brooks, A.N.; Rhind, S.M. The effects of undernutrition, in utero, on reproductive function in adult male and female sheep. Anim. Reprod. Sci. 2002, 72, 63–71. [Google Scholar] [CrossRef]
- Brooks, A.N.; Hagan, D.M.; Sheng, C.; McNeilly, A.S.; Sweeney, T. Prenatal gonadotrophins in the sheep. Anim. Reprod. Sci. 1996, 42, 471–481. [Google Scholar] [CrossRef]
- Hochereau-de Reviers, M.T.; Monet-Kuntz, C.; Courot, M. Spermatogenesis and Sertoli cell numbers and function in rams and bulls. J. Reprod. Fertil. Suppl. 1987, 34, 101–114. [Google Scholar] [CrossRef]
- Senger, P.L. Pathways to Pregnancy & Parturition, 3rd ed.; Current Conceptions: Redmond, OR, USA, 2012; 381p. [Google Scholar]
- Orth, J.M.; Gunsalus, G.L.; Lamperti, A.A. Evidence from Sertoli cell-depleted rats indicates that spermatid number in adults depends on numbers of Sertoli cells produced during perinatal development. Endocrinology 1988, 122, 787–794. [Google Scholar] [CrossRef]
- Copping, K.J.; Hernandez-Medrano, J.; Hoare, A.; Hummitzsch, K.; McMillen, I.C.; Morrison, J.L.; Rodgers, R.J.; Perry, V.E.A. Maternal periconceptional and first trimester protein restriction in beef heifers: Effects on placental parameters and fetal and neonatal calf development. Reprod. Fertil. Dev. 2020, 32, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Tran, D.; Muesy-Dessole, N.; Josso, N. Anti-Mullerian hormone is a functional marker of foetal Sertoli cells. Nature 1977, 269, 411–412. [Google Scholar] [CrossRef] [PubMed]
- Rebourcet, D.; Darbey, A.; Monteiro, A.; Soffientini, U.; Tsai, Y.T.; Handel, I.; Pitetti, J.L.; Nef, S.; Smith, L.B.; O’Shaughnessy, P.J. Sertoli Cell Number Defines and Predicts Germ and Leydig Cell Population Sizes in the Adult Mouse Testis. Endocrinology 2017, 158, 2955–2969. [Google Scholar] [CrossRef] [PubMed]
- Meachem, S.J.; McLachlan, R.I.; de Kretser, D.M.; Robertson, D.M.; Wreford, N.G. Neonatal exposure of rats to recombinant follicle stimulating hormone increases adult Sertoli and spermatogenic cell numbers. Biol. Reprod. 1996, 54, 36–44. [Google Scholar] [CrossRef]
- Kotsampasi, B.; Balaskas, C.; Papadomichelakis, G.; Chadio, S.E. Reduced Sertoli cell number and altered pituitary responsiveness in male lambs undernourished in utero. Anim. Reprod. Sci. 2009, 114, 135–147. [Google Scholar] [CrossRef]
- Andrade, L.P.; Rhind, S.M.; Rae, M.T.; Kyle, C.E.; Jowett, J.; Lea, R.G. Maternal undernutrition does not alter Sertoli cell numbers or the expression of key developmental markers in the mid-gestation ovine fetal testis. J. Negat. Results Biomed. 2013, 12, 2. [Google Scholar] [CrossRef]
- Weller, M.; Fortes, M.R.S.; Marcondes, M.I.; Rotta, P.P.; Gionbeli, T.R.S.; Valadares Filho, S.C.; Campos, M.M.; Silva, F.F.; Silva, W.; Moore, S.; et al. Effect of maternal nutrition and days of gestation on pituitary gland and gonadal gene expression in cattle. J. Dairy Sci. 2016, 99, 3056–3071. [Google Scholar] [CrossRef]
- Cailleau, J.; Vermeire, S.; Verhoeven, G. Independent control of the production of insulin-like growth factor I and its binding protein by cultured testicular cells. Mol. Cell. Endocrinol. 1990, 69, 79–89. [Google Scholar] [CrossRef]
- Pitetti, J.L.; Calvel, P.; Zimmermann, C.; Conne, B.; Papaioannou, M.D.; Aubry, F.; Cederroth, C.R.; Urner, F.; Fumel, B.; Crausaz, M.; et al. An essential role for insulin and IGF1 receptors in regulating sertoli cell proliferation, testis size, and FSH action in mice. Mol. Endocrinol. 2013, 27, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Rae, M.T.; Rhind, S.M.; Kyle, C.E.; Miller, D.W.; Brooks, A.N. Maternal undernutrition alters triiodothyronine concentrations and pituitary response to GnRH in fetal sheep. J. Endocrinol. 2002, 173, 449–455. [Google Scholar] [CrossRef]
- Deligeorgis, S.G.; Chadio, S.; Menegatos, J. Pituitary responsiveness to GnRH in lambs undernourished during fetal life. Anim. Reprod. Sci. 1996, 43, 113–121. [Google Scholar] [CrossRef]
- Thomas, G.B.; McNeilly, A.S.; Gibson, F.; Brooks, A.N. Effects of pituitary-gonadal suppression with a gonadotrophin-releasing hormone agonist on fetal gonadotrophin secretion, fetal gonadal development and maternal steroid secretion in the sheep. J. Endocrinol. 1994, 141, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, P.; Aitken, R.P.; Rhind, S.M.; Racey, P.A.; Wallace, J.M. Influence of placentally mediated fetal growth restriction on the onset of puberty in male and female lambs. Reproduction 2001, 122, 375–383. [Google Scholar] [CrossRef]
- DelCurto, T.; Hess, B.W.; Huston, J.E.; Olson, K.C. Optimum supplementation strategies for beef cattle consuming low-quality roughages in the western United States. J. Anim. Sci. 2000, 77, 1–16. [Google Scholar] [CrossRef]
- Bohnert, D.W.; Stalker, L.A.; Mills, R.R.; Nyman, A.; Falck, S.J.; Cooke, R.F. Late gestation supplementation of beef cows differing in body condition score: Effects on cow and calf performance. J. Anim. Sci. 2013, 91, 5485–5491. [Google Scholar] [CrossRef]
- Herring, C.M.; Bazer, F.W.; Johnson, G.A.; Wu, G. Impacts of maternal dietary protein intake on fetal survival, growth, and development. Exp. Biol. Med. 2018, 243, 525–533. [Google Scholar] [CrossRef]
- Larson, D.M.; Martin, J.L.; Adams, D.C.; Funston, R.N. Winter grazing system and supplementation during late gestation influence performance of beef cows and steer progeny. J. Anim. Sci. 2009, 87, 1147–1155. [Google Scholar] [CrossRef]
- Stalker, L.A.; Adams, D.C.; Klopfenstein, T.J.; Feuz, D.M.; Funston, R.N. Effects of pre- and postpartum nutrition on reproduction in spring calving cows and calf feedlot performance. J. Anim. Sci. 2006, 84, 2582–2589. [Google Scholar] [CrossRef]
- Peine, J.L.; Jia, G.; Van Emon, M.L.; Neville, T.L.; Kirsch, J.D.; Hammer, C.J.; O’Rourke, S.T.; Reynolds, L.P.; Caton, J.S. Effects of maternal nutrition and rumen-protected arginine supplementation on ewe performance and postnatal lamb growth and internal organ mass. J. Anim. Sci. 2018, 96, 3471–3481. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Cudd, T.A.; Meininger, C.J.; Spencer, T.E. Maternal nutrition and fetal development. J. Nutr. 2004, 134, 2169–2172. [Google Scholar] [CrossRef]
- Kwon, H.; Wu, G.; Bazer, F.W.; Spencer, T.E. Developmental changes in polyamine levels and synthesis in the ovine conceptus. Biol. Reprod. 2003, 69, 1626–1634. [Google Scholar] [CrossRef] [PubMed]
- Martín, M.J.; Jiménez, M.D.; Motilva, V. New issues about nitric oxide and its effects on the gastrointestinal tract. Curr. Pharm. Des. 2001, 7, 881–908. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Davis, T.A.; Kim, S.W.; Li, P.; Marc Rhoads, J.; Carey Satterfield, M.; Smith, S.B.; Spencer, T.E.; Yin, Y. Arginine metabolism and nutrition in growth, health and disease. Amino Acids 2009, 37, 153–168. [Google Scholar] [CrossRef]
- Long, N.M.; Vonnahme, K.A.; Hess, B.W.; Nathanielsz, P.W.; Ford, S.P. Effects of early gestational undernutrition on fetal growth, organ development, and placentomal composition in the bovine. J. Anim. Sci. 2009, 87, 1950–1959. [Google Scholar] [CrossRef] [PubMed]
- Long, N.; Cooper, M.P.; Krehbiel, C.; Wettemann, R. Effects of nutrient restriction during early gestation on postnatal growth, carcass and organ weights of beef steers. Biol. Reprod. 2007, 77, 161. [Google Scholar] [CrossRef]
- Ford, S.P. Control of blood flow to the gravid uterus of domestic livestock species. J. Anim. Sci. 1995, 73, 1852–1860. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, L.P.; Redmer, D.A. Utero-placental vascular development and placental function. J. Anim. Sci. 1995, 73, 1839–1851. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Medrano, J.H.; Copping, K.J.; Hoare, A.; Wapanaar, W.; Grivell, R.; Kuchel, T.; Miguel-Pacheco, G.; McMillen, I.C.; Rodgers, R.J.; Perry, V.E. Gestational dietary protein is associated with sex specific decrease in blood flow, fetal heart growth and post-natal blood pressure of progeny. PLoS ONE 2015, 10, e0125694. [Google Scholar] [CrossRef] [PubMed]
- Osgerby, J.C.; Wathes, D.C.; Howard, D.; Gadd, T.S. The effect of maternal undernutrition on ovine fetal growth. J Endocrinol 2002, 173, 131–141. [Google Scholar] [CrossRef]
- Underwood, K.R.; Tong, J.F.; Price, P.L.; Roberts, A.J.; Grings, E.E.; Hess, B.W.; Means, W.J.; Du, M. Nutrition during mid to late gestation affects growth, adipose tissue deposition, and tenderness in cross-bred beef steers. Meat. Sci. 2010, 86, 588–593. [Google Scholar] [CrossRef]
- Cafe, L.M.; Hennessy, D.W.; Hearnshaw, H.; Morris, S.G.; Greenwood, P.L. Consequences of prenatal and preweaning growth for feedlot growth, intake and efficiency of Piedmontese- and Wagyu-sired cattle. Anim. Prod. Sci. 2009, 49, 461–467. [Google Scholar] [CrossRef]
- Funston, R.N.; Summers, A.F.; Roberts, A.J. Alpharma Beef Cattle Nutrition Symposium: Implications of nutritional management for beef cow-calf systems. J. Anim. Sci. 2012, 90, 2301–2307. [Google Scholar] [CrossRef] [PubMed]
- Stalker, L.A.; Ciminski, L.A.; Adams, D.C.; Klopfenstein, T.J.; Clark, R.T. Effects of Weaning Date and Prepartum Protein Supplementation on Cow Performance and Calf Growth. Rangel. Ecol. Manag. 2007, 60, 578–587. [Google Scholar] [CrossRef]
- Copping, K.J.; Ruiz-Diaz, M.D.; Rutland, C.S.; Mongan, N.P.; Callaghan, M.J.; McMillen, I.C.; Rodgers, R.J.; Perry, V.E.A. Peri-conception and first trimester diet modifies reproductive development in bulls. Reprod. Fertil. Dev. 2018, 30, 703–720. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, T.M.; Micke, G.C.; Greer, R.M.; Perry, V.E. Dietary manipulation of Bos indicusxheifers during gestation affects the prepubertal reproductive development of their bull calves. Anim. Reprod. Sci. 2010, 118, 131–139. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCoski, S.; Bradbery, A.; Marques, R.d.S.; Posbergh, C.; Sanford, C. Maternal Nutrition and Developmental Programming of Male Progeny. Animals 2021, 11, 2216. https://doi.org/10.3390/ani11082216
McCoski S, Bradbery A, Marques RdS, Posbergh C, Sanford C. Maternal Nutrition and Developmental Programming of Male Progeny. Animals. 2021; 11(8):2216. https://doi.org/10.3390/ani11082216
Chicago/Turabian StyleMcCoski, Sarah, Amanda Bradbery, Rodrigo da Silva Marques, Christian Posbergh, and Carla Sanford. 2021. "Maternal Nutrition and Developmental Programming of Male Progeny" Animals 11, no. 8: 2216. https://doi.org/10.3390/ani11082216
APA StyleMcCoski, S., Bradbery, A., Marques, R. d. S., Posbergh, C., & Sanford, C. (2021). Maternal Nutrition and Developmental Programming of Male Progeny. Animals, 11(8), 2216. https://doi.org/10.3390/ani11082216