
Hepatic Proteomic Analysis Reveals That Enhanced Carboxylic Acid Metabolism and Oxidoreduction Promote Muscle and Fat Deposition in Muscovy Duck

 ,
,
Abstract
:Simple Summary
Abstract
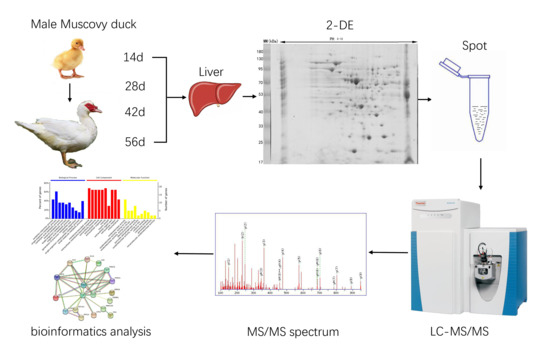
1. Introduction
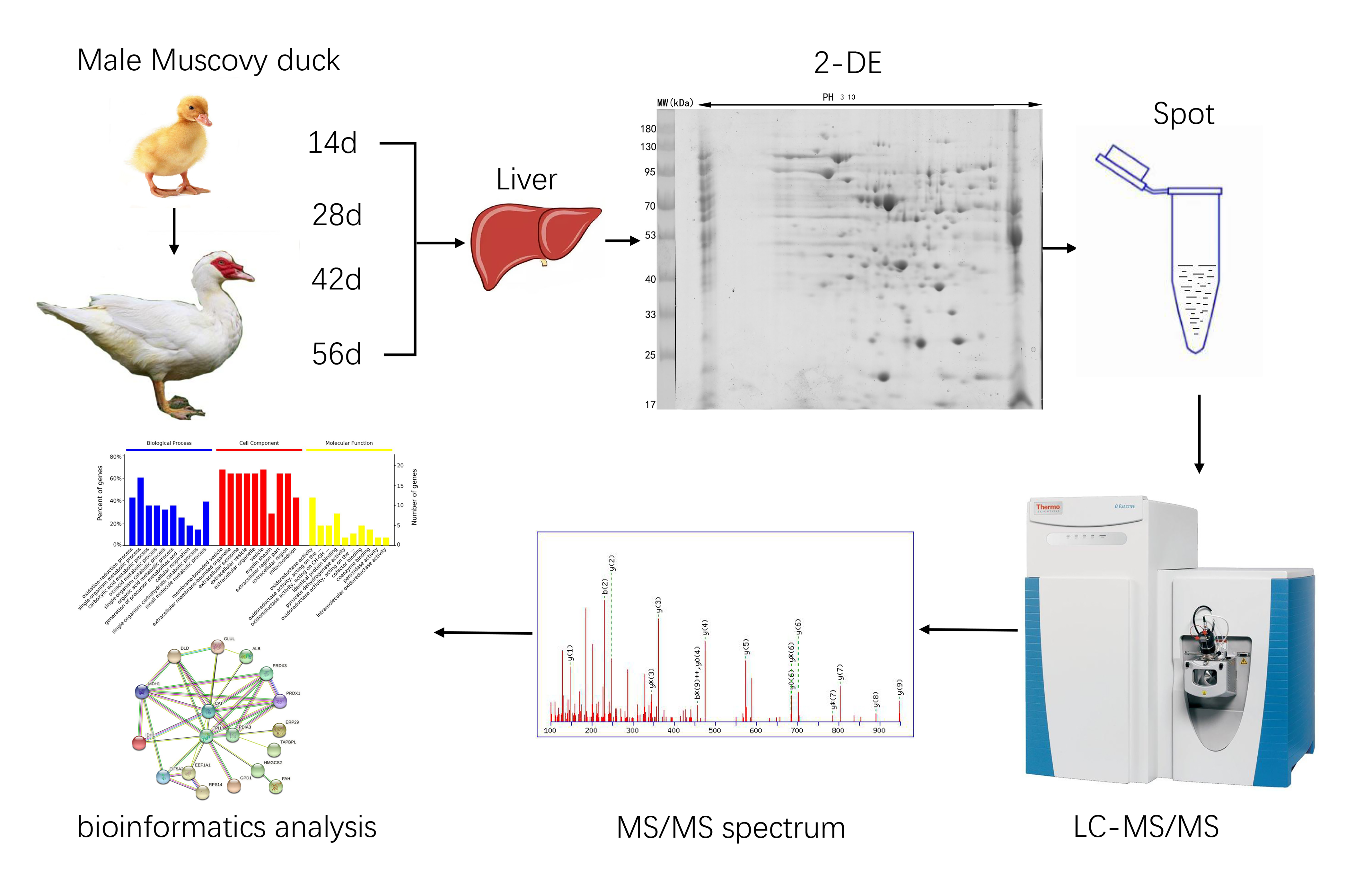
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Animals and Sample Collection
2.3. The Measurement of Growth Trait
2.4. Liver Protein Extraction
2.5. Two-Dimensional Electrophoresis
2.6. Gel Analysis and Protein Spots Identification by LC-MS/MS
2.7. Bioinformatic Analysis and Statistical Analysis
3. Results
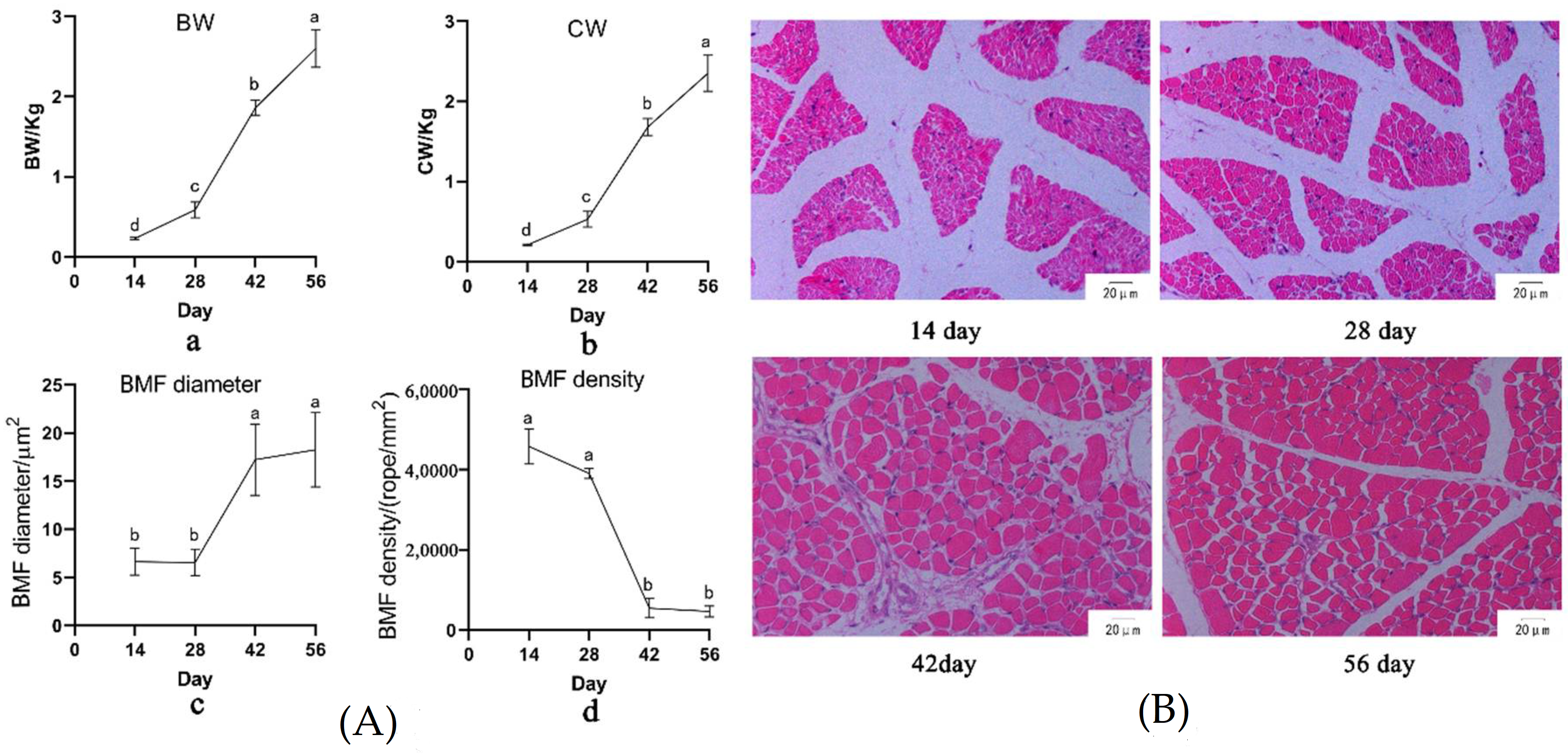
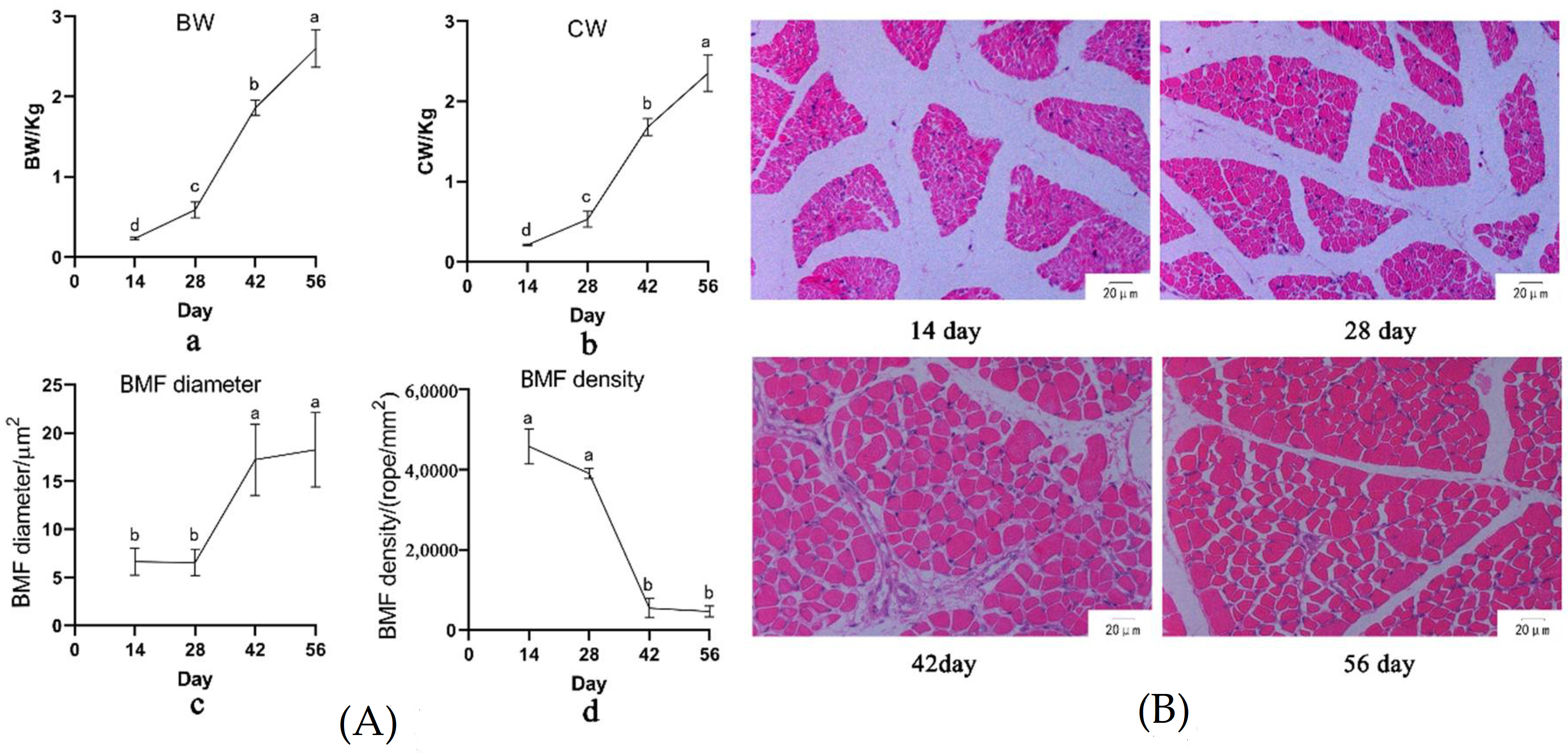
3.1. Growth and Fat Deposition of Muscovy Duck at Different Ages
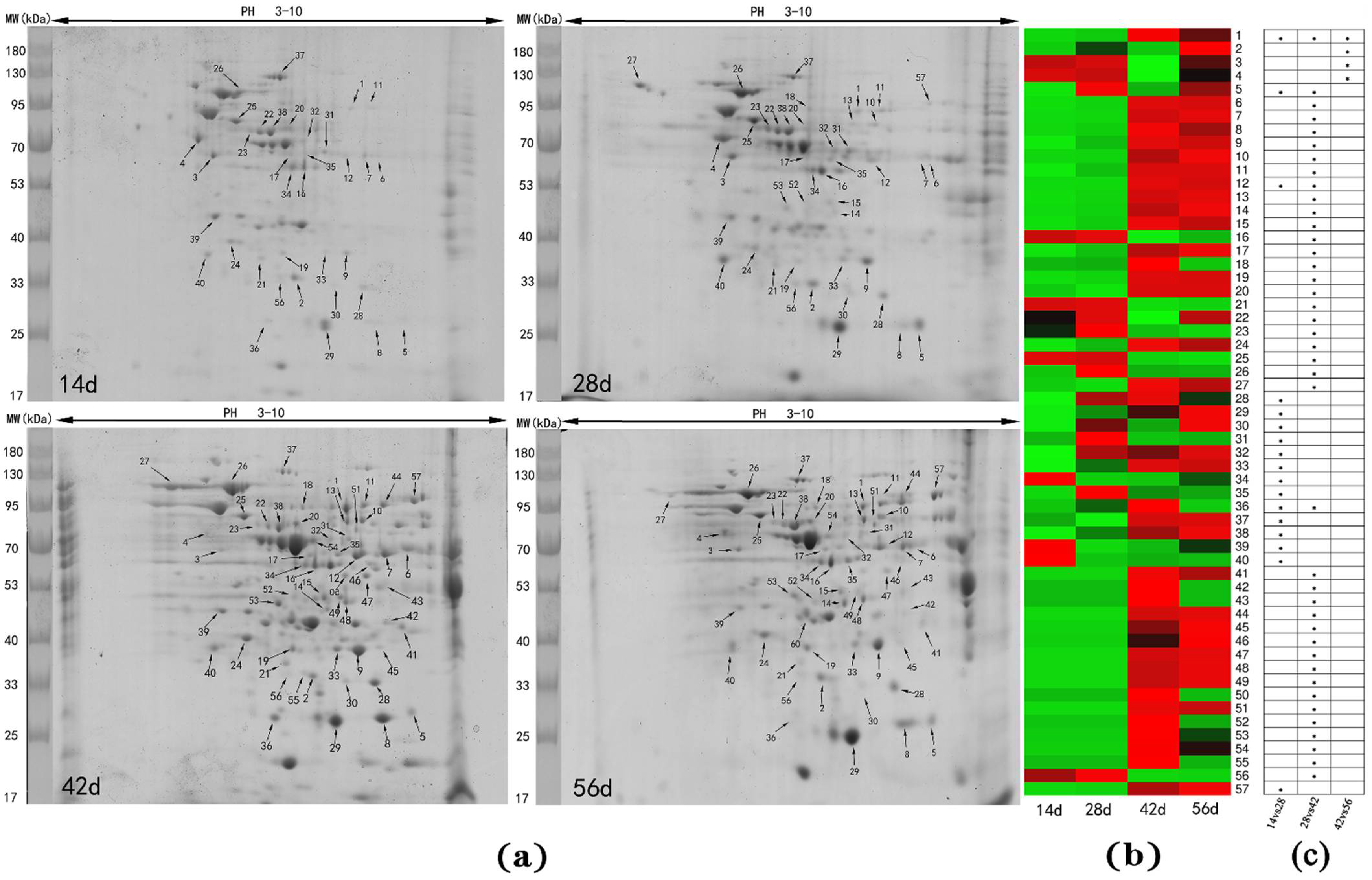
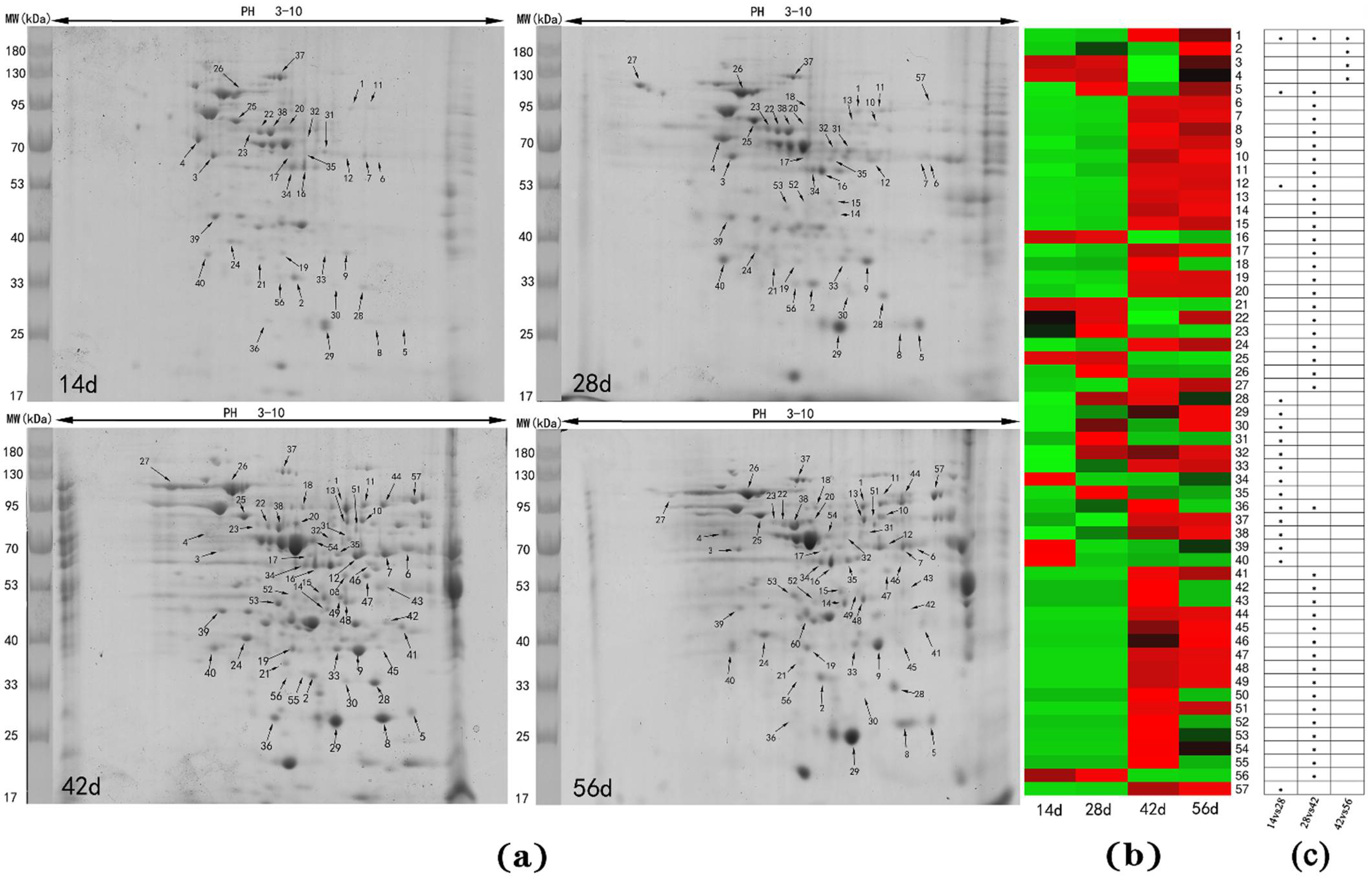
3.2. Differentially Abundant Proteins in Livers at Different Stages
3.3. LC-MS/MS Identification of Differentially Abundant Protein Spots
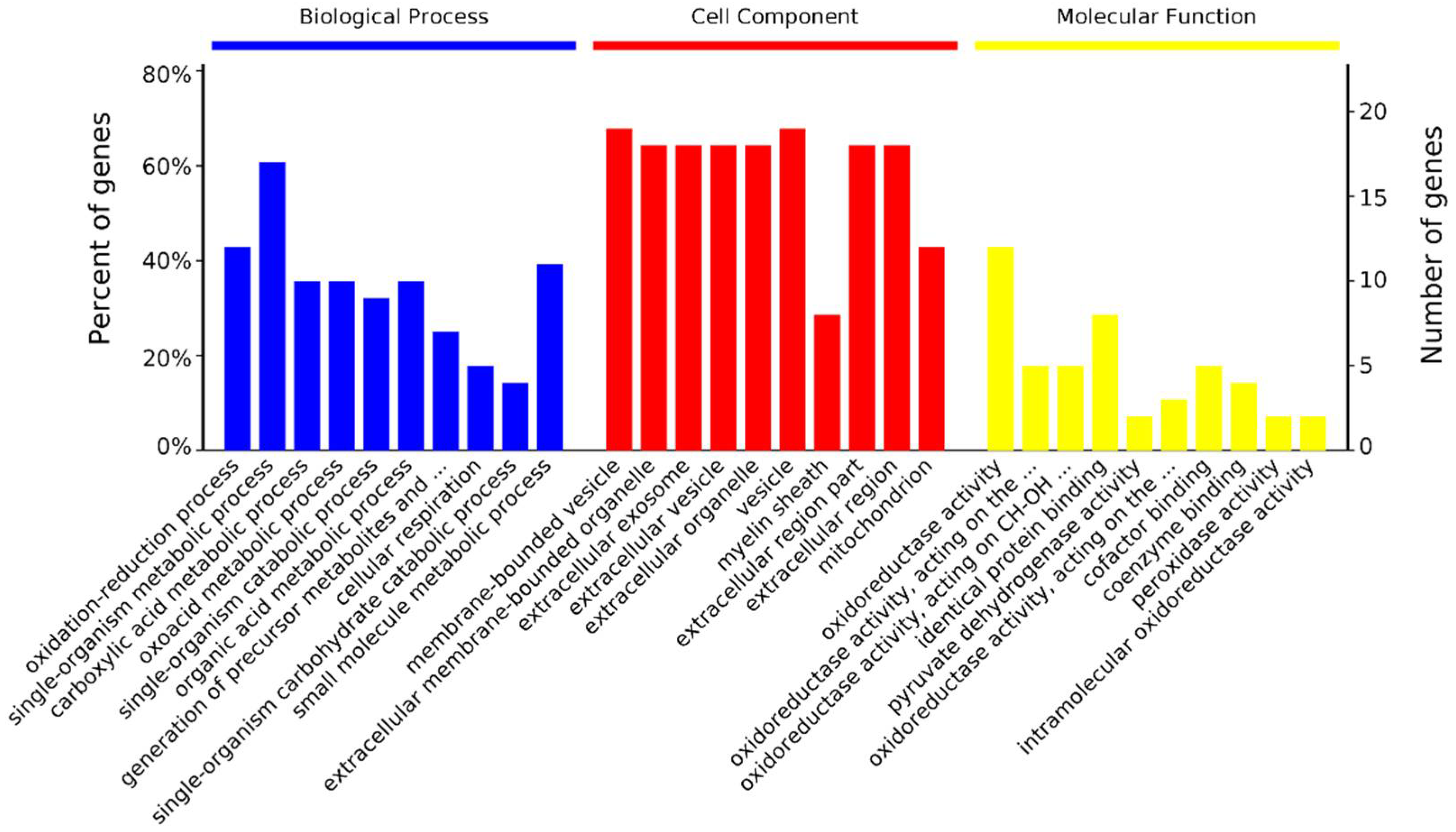
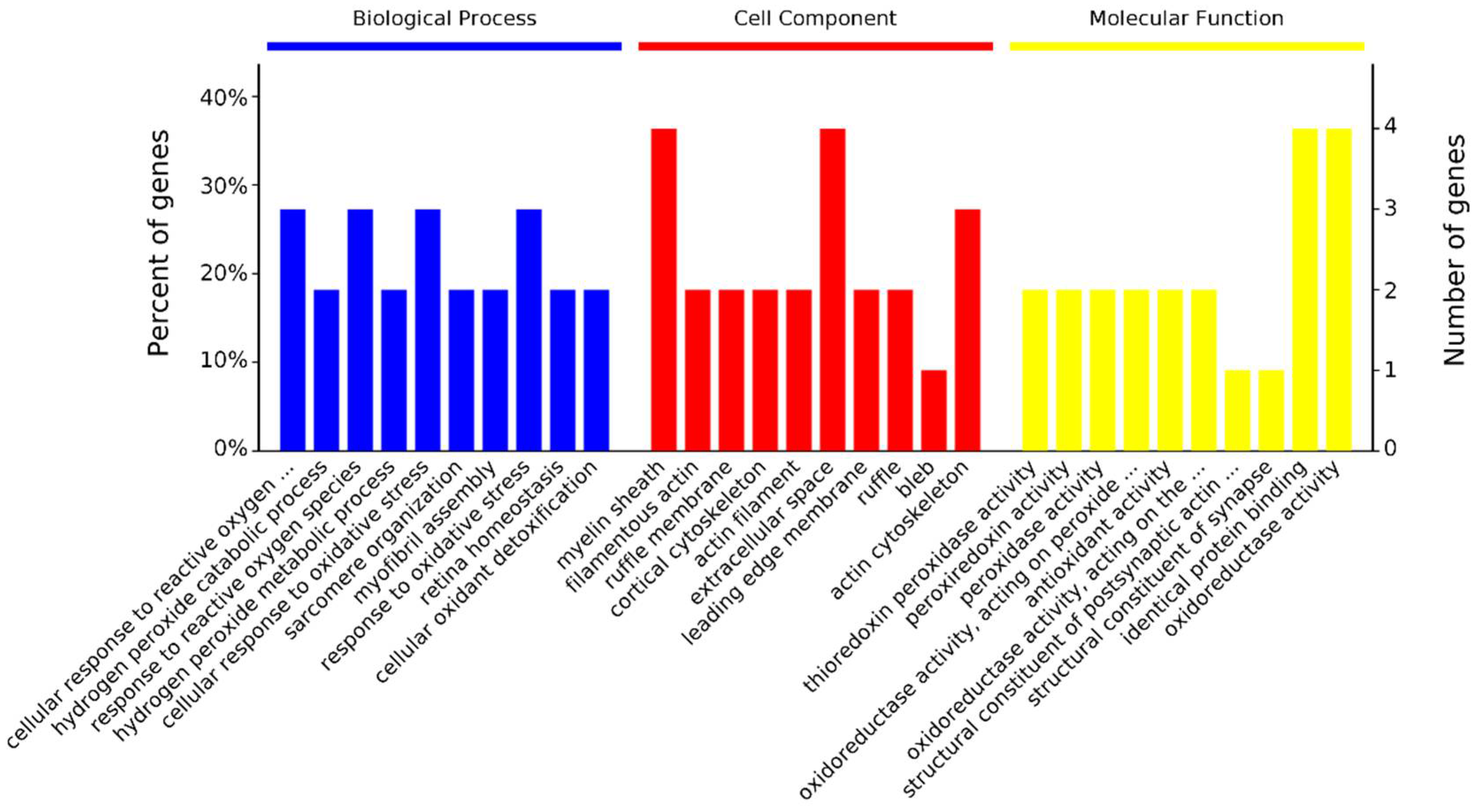
3.4. GO Enrichment for Differentially Abundant Proteins
3.5. Protein–Protein Interaction Analysis of All Identified Differentially Abundant Proteins
4. Discussion
4.1. Growth Performance of Muscovy Duck
4.2. Differentially Abundant Proteins in the Liver of Muscovy Duck
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Chartrin, P.; Bernadet, M.D.; Guy, G.; Mourot, J.; Baéza, E. The effects of genotype and overfeeding on fat level and compostion of adipose and muscle tissues in ducks. Anim. Res. 2006, 55, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Kokoszyński, D.; Saleh, M.; Bernacki, Z.; Topoliński, T.; Andryszczyk, M.; Wirwicki, M. Growth performance, carcass composition, leg bones and digestive system characteristics in Pekin duck broilers fed a diet diluted with whole wheat grain. Can. J. Anim. Sci. 2019, 99, 781–791. [Google Scholar] [CrossRef]
- Chartrin, P.; Méteau, K.; Juin, H.; Bernadet, M.D.; Guy, G.; Larzul, C.; Rémignon, H.; Mourot, J.; Duclos, M.J.; Baéza, E. Effects of Intramuscular Fat Levels on Sensory Characteristics of Duck Breast Meat. Poult. Sci. 2006, 85, 914–922. [Google Scholar] [CrossRef]
- Saez, G.; Davail, S.; Gentes, G.; Hocquette, J.F.; Jourdan, T.; Degrace, P.; Baeza, E. Gene expression and protein content in relation to intramuscular fat content in Muscovy and Pekin ducks. Poult. Sci. 2009, 88, 2382–2391. [Google Scholar] [CrossRef] [PubMed]
- Kokoszyński, D.; Wilkanowska, A.; Saleh, M.; Martin, F.; Bartosz, B. Comparison of some meat and liver quality traits in Muscovy and Pekin ducks. J. Appl. Anim. Res. 2021, 49, 118–124. [Google Scholar] [CrossRef]
- Liu, X.; Liu, Y.; Cheng, H.; Deng, Y.; Xiong, X.; Qu, X. Comparison of performance, fatty acid composition, enzymes and gene expression between overfed Xupu geese with large and small liver. Ital. J. Anim. Sci. 2021, 20, 102–111. [Google Scholar] [CrossRef]
- Gontier, K.; André, J.M.; Bernadet, M.D.; Ricaud, K.; Davail, S.J.C.B. Insulin effect on lipogenesis and fat reserve distribu tion in three genetic types of ducks during overfeeding. Comp. Biochem. Physiol. Part A 2013, 164, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Hermier, D.; Salichon, M.R.; Guy, G.; Peresson, R. Differential channelling of liver lipids in relation to susceptibility to hepatic steatosis in two species of ducks. Poult. Sci. 1999, 78, 1398–1406. [Google Scholar] [CrossRef]
- Hérault, F.; Saez, G.; Robert, E.; Mohammad, A.A.; Davail, S.; Chartrin, P.; Baéza, E.; Diot, C. Liver gene expression in relation to hepatic steatosis and lipid secretion in two duck species. Anim. Genet. 2010, 41, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, X.; Ma, J.; Zhang, Y.; Zhang, H. Integrating genome and transcriptome profiling for elucidating the mechanism of muscle growth and lipid deposition in Pekin ducks. Sci. Rep. 2017, 7, 3837. [Google Scholar] [CrossRef]
- François, Y.; Marie-Etancelin, C.; Vignal, A.; Viala, D.; Davail, S.; Molette, C. Mule Duck “Foie Gras” Shows Different Metabolic States According to Its Quality Phenotype by Using a Proteomic Approach. J. Agric. Food Chem. 2014, 62, 7140–7150. [Google Scholar] [CrossRef]
- Zeng, T.; Jiang, X.; Li, J.; Wang, D.; Li, G.; Lu, L.; Wang, G.; Sompop, B. Comparative Proteomic Analysis of the Hepatic Response to Heat Stress in Muscovy and Pekin Ducks: Insight into Thermal Tolerance Related to Energy Metabolism. PLoS ONE 2013, 8, e76917. [Google Scholar] [CrossRef]
- Zheng, A.; Liu, G.; Zhang, Y.; Hou, S.; Chang, W.; Zhang, S.; Cai, H.; Chen, G. Proteomic analysis of liver development of lean Pekin duck (Anas platyrhynchos domestica). J. Proteom. 2012, 75, 5396–5413. [Google Scholar] [CrossRef]
- Zheng, A.; Chang, W.; Hou, S.; Zhang, S.; Cai, H.; Chen, G.; Lou, R.; Liu, G. Unraveling molecular mechanistic differences in liver metabolism between lean and fat lines of Pekin duck (Anas platyrhynchos domestica): A proteomic study. J. Proteom. 2014, 98, 271–288. [Google Scholar] [CrossRef] [PubMed]
- Dale, N. National research council nutrient requirements of poultry-ninth revised edition (1994). J. Appl. Poult. Res. 1994, 3, 101. [Google Scholar] [CrossRef]
- Chen, X.; Zhu, W.; Liu, X.; Li, T.; Geng, Z.; Wan, X. The growth performance, meat quality, and gut bacteria of broilers raised with or without antibiotics and green tea powder. J. Appl. Poult. Res. 2019, 28, 712–721. [Google Scholar] [CrossRef]
- Xu, T.S.; Gu, L.H.; Sun, Y.; Zhang, H.X.; Hou, S.S. Characterization of MUSTN1 gene and its relationship with skeletal muscle development at postnatal stages in Pekin ducks. Genet. Mol. Res. 2015, 14, 4448–4460. [Google Scholar] [CrossRef]
- Zhu, W.; Zhang, J.; He, K.; Geng, Z.; Chen, X. Proteomic analysis of fertilized egg yolk proteins during embryonic development. Poult. Sci. 2020, 99, 2775–2784. [Google Scholar] [CrossRef]
- Baeza, E.; Dessay, C.; Wacrenier, N.; Marche, G.; Listrat, A. Effect of selection for improved body weight and composition on muscle and meat characteristics in Muscovy duck. Br. Poult. Sci. 2002, 43, 560–568. [Google Scholar] [CrossRef]
- Murawska, D. The effect of age on the growth rate of tissues and organs and the percentage content of edible and nonedible carcass components in Pekin ducks. Poult. Sci. 2012, 91, 2030–2038. [Google Scholar] [CrossRef]
- Sonaiya, E.B.; Benyi, K. Abdominal Fat in 12- to 16-Week-Old Broiler Birds as Influenced by Age, Sex, and Strain. Poult. Sci. 1983, 62, 1793–1799. [Google Scholar] [CrossRef]
- Damon, M.; Louveau, I.; Lefaucheur, L.; Lebret, B.; Vincent, A.; Leroy, P.; Sanchez, M.P.; Herpin, P.; Gondret, F. Number of intramuscular adipocytes and fatty acid binding protein-4 content are significant indicators of intramuscular fat level in crossbred Large White Duroc pigs. J. Anim. Sci. 2006, 84, 1083–1092. [Google Scholar] [CrossRef]
- Li, D.; Pan, Z.; Zhang, K.; Yu, M.; Yu, D.; Lu, Y.; Wang, J.; Zhang, J.; Zhang, K.; Du, W. Identification of the Differentially Expressed Genes of Muscle Growth and Intramuscular Fat Metabolism in the Development Stage of Yellow Broilers. Genes 2020, 11, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, R.N.S.J.C.B. Isocitrate dehydrogenase activity and its regulation by estradiol in tissues of rats of various ages. Cell Biochem. Funct. 1988, 6, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, L.; Li, Y.; Fu, J.; Zhen, L.; Yang, Q.; Li, S. Tyrosine phosphorylation of dihydrolipoamide dehydrogenase as a potential cadmium target and its inhibitory role in regulating mouse sperm motility. Toxicology 2016, 357, 52–64. [Google Scholar] [CrossRef]
- Breźnicka, E.A.; Chmielnicka, J.; Wójcikiewicz-Herma, L. Tissue origin of MDH isozymes in blood serum of rats exposed to alkylmercurials. J. Appl. Toxicol. 1983, 3, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xu, C.; Shi, J.; Ding, J.; Wan, X.; Chen, D.; Gao, J.; Li, C.; Zhang, J.; Lin, Y.; et al. Fatty acids promote fatty liver disease via the dysregulation of 3-mercaptopyruvate sulfurtransferase/hydrogen sulfide pathway. Gut 2017, 67, 2169–2180. [Google Scholar] [CrossRef]
- Tommaso, L.D.; Franchi, G.; Park, Y.N.; Fiamengo, B.; Destro, A.; Morenghi, E.; Montorsi, M.; Torzilli, G.; Tommasini, M.; Terracciano, L. Diagnostic value of HSP70, glypican 3, and glutamine synthetase in hepatocellular nodules in cirrhosis. Hepatology 2010, 45, 725–734. [Google Scholar] [CrossRef]
- Awano, S.; Koshimune, S.; Kurihara, E.; Gohara, K.; Sakai, A.; Soh, I.; Hamasaki, T.; Ansai, T.; Takehara, T. The assessment of methyl mercaptan, an important clinical marker for the diagnosis of oral malodor. J. Dent. 2004, 32, 555–559. [Google Scholar] [CrossRef]
- Nakano, M.; Shin, K.; Wakabayashi, H.; Yamauchi, K.; Hironaka, S. Inactivating effects of the lactoperoxidase system on bacterial lyases involved in oral malodour production. J. Med. Microbiol. 2015, 64, 1244–1252. [Google Scholar] [CrossRef]
- Yu, G.; Wang, Y.; Wang, S.; Duan, C.; Wei, L.; Gao, J.; Chai, T.; Cai, Y. Effects of Microbial Aerosol in Poultry House on Meat Ducks’ Immune Function. Front. Microbiol. 2016, 7, 1245. [Google Scholar] [CrossRef] [Green Version]
- Yabe-Nishimura, C. Aldose reductase in glucose toxicity: A potential target for the prevention of diabetic complications. Pharmacol. Rev. 1998, 50, 21–33. [Google Scholar] [PubMed]
- Mcnulty, H.; Mcpartlin, J.M.; Weir, D.G.; Scott, J.M. Folate catabolism is related to growth rate in weanling rats. J. Nutr. 1995, 125, 99–103. [Google Scholar] [PubMed]
- Istvan, E.S.; Palnitkar, M.; Buchanan, S.K.; Deisenhofer, J. Crystal structure of the catalytic portion of human HMG-CoA reductase: Insights into regulation of activity and catalysis. EMBO J. 2014, 19, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Guémené, D.; Guy, G. The past, present and future of force-feeding and “foie gras” production. World’s Poult. Sci, J. 2004, 60, 210–222. [Google Scholar] [CrossRef]
- Parim, B.N.; Harishankar, N.; Balaji, M.; Santosh, K.M.; Sailaja, P.; Sajjalaguddam, R. Mitigating efficacy of piperine in the physiological derangements of high fat diet induced obesity in Sprague Dawley rats. Chem. Biol. Interact. 2014, 221, 42–51. [Google Scholar]
- Spurlock, M.E.; Savage, J.E. Effect of Dietary Protein and Selected Antioxidants on Fatty Liver Hemorrhagic Syndrome Induced in Japanese Quail1. Poult. Sci. 1993, 72, 2095–2105. [Google Scholar] [CrossRef]
- Wang, C.; Chen, Z.; Li, S.; Zhang, Y.; Jia, S.; Li, J.; Chi, Y.; Miao, Y.; Guan, Y.; Yang, J. Hepatic overexpression of ATP synthase β subunit activates PI3K/Akt pathway to ameliorate hyperglycemia of diabetic mice. Diabetes 2014, 63, 947–959. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Bara, L.; Nathalie, M.-G.; Hélène, M.; Carole, B.; Céline, B.; Renaud, D.; Hervé, R. Cellular markers of mule duck livers after force-feeding. Poult. Sci. 2020, 99, 3567–3573. [Google Scholar]
- Rhee, S.G.; Woo, H.A.; Kil, I.S.; Bae, S.H. Peroxiredoxin functions as a peroxidase and a regulator and sensor of local peroxides. J. Biol. Chem. 2012, 287, 4403–4410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, A.G.; Winterbourn, C.C.; Hampton, M.B. Mitochondrial peroxiredoxin involvement in antioxidant defence and redox signalling. Biochem. J. 2010, 425, 313–325. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.Y.; Lee, J.S.; Alves, T.; Ladiges, W.; Rabinovitch, P.S. Mitochondrial Targeted Catalase Protects Against High-Fat Diet-Induced Muscle Insulin Resistance by Decreasing Intramuscular Lipid Accumulation. Diabetes 2017, 66, 2072. [Google Scholar] [CrossRef] [Green Version]
- Simitzis, P.E.; Kalogeraki, E.; Goliomytis, M.; Charismiadou, M.A.; Triantaphyllopoulos, K.; Ayoutanti, A.; Niforou, K.; Hager-Theodorides, A.L.; Deligeorgis, S.G. Impact of stocking density on broiler growth performance, meat characteristics, behavioural components and indicators of physiological and oxidative stress. Bri. Poult. Sci. 2012, 53, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.J.; Won, J.K.; Ryu, H.S.; Choi, W.M.; Cho, H.; Cho, E.J.; Lee, J.H.; Kim, Y.J.; Suh, K.S.; Jang, J.J. A novel prognostic factor for hepatocellular carcinoma: Protein disulfide isomerase. Korean J. Intern. Med. 2014, 29, 580–587. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
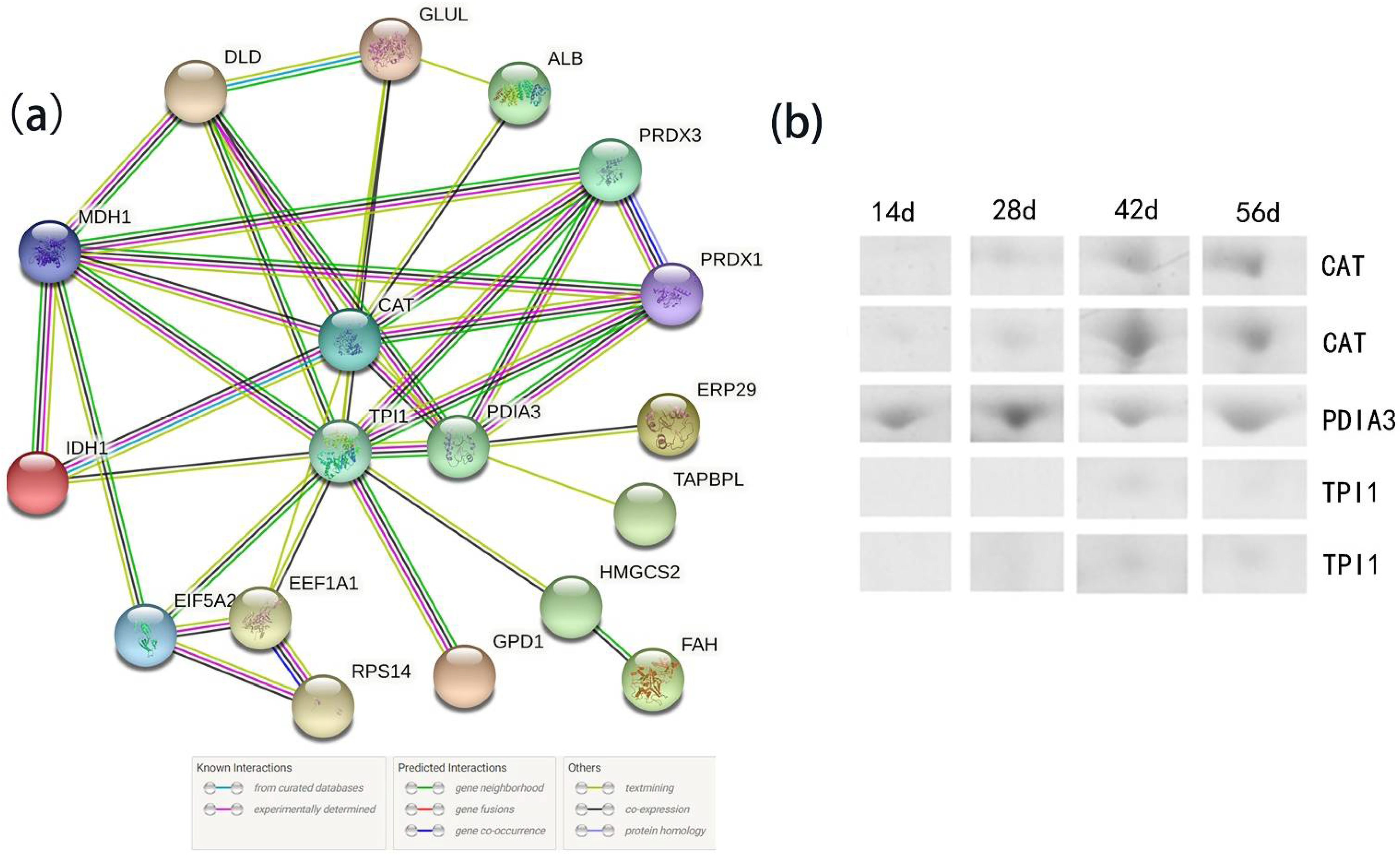{kind=link}
| Age (Day) | IMF 1 % | SFT 2 (mm) | AFW 3 (g) |
|---|---|---|---|
| 14 | / | 1.37 b ± 0.252 | 0.45 c ± 0.370 |
| 28 | 2.89 a ± 0.65 | 1.62 b ± 0.274 | 3.31 c ± 1.73 |
| 42 | 1.15 b ± 0.27 | 2.50 a ± 0.556 | 28.23 b ± 6.95 |
| 56 | 1.25 b ± 0.36 | 2.38 a ± 0.557 | 44.23 a ± 12.189 |
| Sort | Spot 1 | Accession 2 | Proteins Description | S 3 | M 4 | SC 5 (%) | MW (kDa)/pI 6 | |
|---|---|---|---|---|---|---|---|---|
| Thero. | Obs. | |||||||
| Enzymes and catalytic function | 1 | A0A493TQM7 | Catalase (CAT) | 127 | 12 | 14.8 | 60.2/7.7 | 95.0/7.9 |
| 11 | A0A493TQM7 | Catalase (CAT) | 92 | 8 | 9.3 | 60.2/7.7 | 95.0/8.2 | |
| 6 | U3J597 | Isocitrate dehydrogenase [NADP] (IDH) | 439 | 24 | 27.5 | 47.0/7.6 | 70.0/9.0 | |
| 7 | U3J597 | Isocitrate dehydrogenase [NADP] (IDH) | 467 | 24 | 27.5 | 47.0/7.6 | 70.0/8.7 | |
| 9 | U3I1N5 | Sulfurtransferase (TST) | 187 | 15 | 25.2 | 33.1/7.1 | 39.0/8.2 | |
| 42 | U3I1N5 | Sulfurtransferase (TST) | 70 | 3 | 8.8 | 33.1/7.1 | 41.0/8.7 | |
| 10 | U3IR48 | Dihydrolipoyl dehydrogenase (DLD) | 932 | 35 | 22.5 | 50.1/8.4 | 82.0/8.2 | |
| 13 | U3IR48 | Dihydrolipoyl dehydrogenase (DLD) | 220 | 8 | 12.8 | 50.1/8.4 | 82.0/8.0 | |
| 51 | U3IR48 | Dihydrolipoyl dehydrogenase (DLD) | 88 | 3 | 7.4 | 50.1/8.4 | 72.0/8.1 | |
| 12 | A0A493SW39 | Fumarylacetoacetase (FAH) | 221 | 13 | 16.2 | 46.7/7.2 | 68.0/8.2 | |
| 14 | A0A493T656 | Glycerol-3-phosphate dehydrogenase [NAD(+)] (GPD1) | 57 | 7 | 10.3 | 39.5/8.2 | 43.0/7.9 | |
| 56 | A0A493TUY2 | Glyceraldehyde-3-phosphate dehydrogenase (TAPBL) | 63 | 7 | 10.6 | 42.2/5.3 | 34.0/7.4 | |
| 15 | U3IR57 | Malate dehydrogenase (MDH1) | 115 | 9 | 19.8 | 36.7/6.9 | 45.0/7.8 | |
| 53 | U3IR57 | Malate dehydrogenase (MDH1) | 245 | 11 | 17.7 | 36.7/6.9 | 44.0/7.1 | |
| 16 | A0A493T5Z6 | Glutamine synthetase (GS) | 211 | 12 | 11.4 | 52.0/7.6 | 55.0/7.7 | |
| 17 | A0A493T5Z6 | Glutamine synthetase (GS) | 123 | 4 | 3.5 | 52.0/7.6 | 55.0/7.6 | |
| 48 | A0A493T5Z6 | Glutamine synthetase (GS) | 55 | 2 | 4.5 | 52.0/7.6 | 49.0/8.0 | |
| 19 | A0A493TH73 | Quinoid dihydropteridine reductase (QDPR) | 187 | 2 | 15.5 | 28.2/6.1 | 39.0/7.3 | |
| 20 | A0A493TX20 | Methyl mercaptan (MM) oxidase | 57 | 6 | 9.3 | 57.2/7.2 | 75.0/7.2 | |
| 25 | U3IS54 | Protein disulfide-isomerase (PDIA3) | 615 | 47 | 38.1 | 56.6/5.8 | 80.0/6.3 | |
| 33 | A0A493U078 | Peroxiredoxin 3 (PRDX3) | 146 | 8 | 12.9 | 29.1/9.3 | 39.0/7.8 | |
| 36 | U3II01 | Peroxiredoxin 1 (PRDX1) | 193 | 11 | 19.1 | 22.6/8.2 | 27.0/7.0 | |
| 34 | A0A493STH1 | Pyruvate dehydrogenase E1 component subunit alpha (PDH) | 226 | 11 | 16.9 | 45.4/8.2 | 58.0/7.5 | |
| 46 | A0A493STH1 | Pyruvate dehydrogenase E1 component subunit alpha (PDH) | 332 | 15 | 20.8 | 45.4/8.2 | 58.0/8.7 | |
| 45 | U3I8D8 | Triosephosphate isomerase (TPI1) | 233 | 11 | 27 | 24.2/7.7 | 40.0/8.5 | |
| 52 | U3I8D8 | Triosephosphate isomerase (TPI1) | 387 | 16 | 26.1 | 24.2/7.7 | 49.0/7.3 | |
| 47 | U3II47 | Aldo-keto reductase family 1 member A1 (AKR1A1) | 285 | 16 | 16.5 | 37.3/7.2 | 54.0/8.5 | |
| 50 | U3II47 | Aldo-keto reductase family 1 member A1 (AKR1A1) | 74 | 4 | 7.6 | 37.3/7.2 | 54.0/8.1 | |
| 49 | U3J9Y2 | L-lactate dehydrogenase (LD) | 360 | 16 | 24.3 | 36.5/7.1 | 52.0/8.0 | |
| 54 | A0A493T8G2 | 3-hydroxy-3-methylglutaryl coenzyme A synthase (HMGCS2) | 231 | 9 | 7.5 | 60.6/8.7 | 72.0/7.5 | |
| 55 | U3IT75 | 5-formyltetrahydrofolate cyclo-ligase | 74 | 9 | 21.6 | 24.1/6.4 | 34.0/7.0 | |
| Structural and functional proteins | 2 | A0A493TSC9 | Zinc finger and BTB domain containing22 (ZBTB22) | 70 | 2 | 1.7 | 57.5/6.6 | 34.0/7.5 |
| 27 | A0A493TSC9 | Zinc finger and BTB domain containing 22 (ZBTB22) | 43 | 3 | 1.7 | 23.2/5.2 | 98.0/5.0 | |
| 4 | U3IP65 | ATP synthase subunit beta (ATP5B) | 1087 | 54 | 38.7 | 53.5/5.2 | 75.0/5.7 | |
| 18 | U3J2L0 | Ribosomal protein S14 (RPS14) | 66 | 1 | 7.1 | 16.6/8.9 | 93.0/7.4 | |
| 22 | U3IL97 | Keratin 12 (KRT12) | 123 | 6 | 8.4 | 49.9/5.0 | 75.0/6.7 | |
| 23 | U3IM27 | Aldedh domain-containing protein | 141 | 12 | 17.1 | 46.7/5.7 | 75.0/6.5 | |
| 38 | U3IM27 | Aldedh domain-containing protein | 314 | 22 | 19.9 | 46.7/5.7 | 75.0/6.9 | |
| 24 | A0A493TAD1 | Eukaryotic translation initiation factor 5A (EIF5A2) | 130 | 5 | 5.6 | 15.9/5.5 | 42.0/6.4 | |
| 57 | A0A493SUB8 | Elongation factor 1-alpha (EEF1A1) | 100 | 3 | 4.1 | 50.5/9.1 | 96.0/9.1 | |
| 26 | A0A493TIR6 | Albumin (ALB) | 2282 | 144 | 62.5 | 69.3/5.9 | 98.0/6.2 | |
| 29 | A0A493T6B1 | IF rod domain-containing protein | 93 | 3 | 3.4 | 52.7/4.8 | 26.0/7.9 | |
| 43 | A0A493T6B1 | IF rod domain-containing protein | 102 | 4 | 3.4 | 52.7/4.8 | 53.0/8.7 | |
| 30 | A0A493TP63 | IF rod domain-containing protein | 71 | 4 | 5.8 | 55.3/4.9 | 33.0/8.1 | |
| 32 | A0A493U1E6 | Histone H2B | 93 | 3 | 15.1 | 14.0/10.3 | 70.0/7.8 | |
| 37 | A0A493TBB4 | Ovotransferrin | 328 | 25 | 18.8 | 82.6/6.9 | 130.0/7.0 | |
| 39 | A0A493SXF3 | Tropomyosin 4 (TPM4) | 33 | 1 | 2.4 | 28.8/4.6 | 45.0/6.0 | |
| 41 | U3ILS4 | Endoplasmic reticulum protein 29 (ERP29) | 207 | 19 | 26.8 | 29.2/8.6 | 41.0/9.2 | |
| 44 | U3J7T0 | Myosin motor domain-containing protein | 1588 | 102 | 30.3 | 187.8/5.4 | 95.0/8.8 | |
| Uncharacterized proteins | 5 | A0A493SU68 | Uncharacterized protein | 66 | 19 | 10.5 | 23.3/5.2 | 27.0/9.0 |
| 21 | A0A493SU68 | Uncharacterized protein | 70 | 3 | 10.5 | 23.2/5.2 | 34.0/7.2 | |
| 31 | A0A493SU68 | Uncharacterized protein | 70 | 2 | 10.5 | 23.2/5.2 | 70.0/8.1 | |
| 35 | A0A493SU68 | Uncharacterized protein | 76 | 2 | 10.5 | 23.2/5.2 | 69.0/8.2 | |
| 8 | A0A493TMK3 | Uncharacterized protein | 756 | 43 | 29.4 | 37.1/9.3 | 26.0/8.5 | |
| 3 | A0A493T8E7 | Uncharacterized protein (ACTG1) | 641 | 44 | 34.6 | 42.2/5.3 | 70.0/6.1 | |
| 40 | A0A493T8E7 | Uncharacterized protein (ACTG1) | 87 | 5 | 4.5 | 42.2/5.3 | 40.0/6.1 | |
| 28 | A0A493TX61 | Uncharacterized protein | 261 | 26 | 30.8 | 20.9/5.8 | 39.0/8.4 | |
| 14 vs. 28 Day | 28 vs. 42 Day | 42 vs. 56 Day | |
|---|---|---|---|
| Significant spots | 17 | 41 | 4 |
| Significant proteins | 13 | 39 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, W.; Chen, X.; Wei, C.; Zhao, Y.; Liu, Z.; Geng, Z. Hepatic Proteomic Analysis Reveals That Enhanced Carboxylic Acid Metabolism and Oxidoreduction Promote Muscle and Fat Deposition in Muscovy Duck. Animals 2021, 11, 2180. https://doi.org/10.3390/ani11082180
Yang W, Chen X, Wei C, Zhao Y, Liu Z, Geng Z. Hepatic Proteomic Analysis Reveals That Enhanced Carboxylic Acid Metabolism and Oxidoreduction Promote Muscle and Fat Deposition in Muscovy Duck. Animals. 2021; 11(8):2180. https://doi.org/10.3390/ani11082180
Chicago/Turabian StyleYang, Wanli, Xingyong Chen, Congcong Wei, Yutong Zhao, Zhengquan Liu, and Zhaoyu Geng. 2021. "Hepatic Proteomic Analysis Reveals That Enhanced Carboxylic Acid Metabolism and Oxidoreduction Promote Muscle and Fat Deposition in Muscovy Duck" Animals 11, no. 8: 2180. https://doi.org/10.3390/ani11082180
APA StyleYang, W., Chen, X., Wei, C., Zhao, Y., Liu, Z., & Geng, Z. (2021). Hepatic Proteomic Analysis Reveals That Enhanced Carboxylic Acid Metabolism and Oxidoreduction Promote Muscle and Fat Deposition in Muscovy Duck. Animals, 11(8), 2180. https://doi.org/10.3390/ani11082180