1. Introduction
Porcine proliferative enteropathy (PPE), commonly referred to as ileitis, is caused by Gram-negative anaerobic obligate intracellular bacterium
Lawsonia intracellularis, which is an economically important swine disease that poses significant threats to the swine industry worldwide [
1,
2]. PPE disease is commonly observed in post-weaned and growing pigs less than 4 months of age and characterized by acute diarrhea, weakness, decreased weight gain, intestinal hemorrhage, and sudden death [
3]. The mode of transmission of
L.
intracellularis is via the fecal–oral route, and lesions are characterized by thickening of the mucosa of the ileum and colon [
2,
4]. To date, prophylaxis against
L.
intracellularis infection is achieved by either administration of antibiotic growth promoters or vaccination with live attenuated
L.
intracellularis vaccine [
5]. In a recent study,
Salmonella-based
L.
intracellularis vaccines induced
L.
intracellularis-specific humoral and cell mediated immunities and conferred dual protection against PPE and salmonellosis in a murine model [
6].
In recent years, considerable research has revealed the importance of the gut microbiome in the health and growth of animals. The intestinal microbiome plays a crucial role in the development and regulation of the immune system and therefore, its composition may affect how animals respond to vaccinations [
7]. It has been recognized that the intestinal microbiome affects the immune response to natural infection or vaccination [
8]. For example, Leite et al. revealed that vaccination against
L.
intracellularis decreased levels of
S. enterica serovar Typhimurium shedding in co-infected pigs, indicating that vaccination against
L.
intracellularis may be used as a tool to prevent foodborne diseases associated with
Salmonella [
9].
L.
intracellularis seroconversion has been identified as a risk factor for the increased prevalence of pigs shedding
Salmonella enterica subsp.
enterica in farrow-to-finish herds [
10].
However, information regarding the effects of oral vaccination with live attenuated vaccine against L. intracellularis in the intestinal microbial composition of pigs is limited. Furthermore, the changes in the intestinal microbiota composition in response to vaccination against L. intracellularis are not fully understood. This study was performed to determine the effects of oral vaccination against L. intracellularis on the intestinal microbiome of weaned piglets. To do this, pigs were experimentally vaccinated with different doses of L. intracellularis vaccine; their microbial community composition was measured before and after oral administration; and finally, the microbiota of vaccinated pigs were compared to those of non-vaccinated pigs. We hypothesized that different doses of live attenuated vaccine against L. intracellularis would cause alteration in the composition of the pig gut microbiome. Understanding the impact that the pig gut microbiome plays in response to vaccination will improve our current understanding of vaccine efficacy and the underlying changes in the composition of the microbiome that may be associated with the susceptibility of pigs to PPE. The microbial biomarkers identified in this study may be used for the development of strategies to improve vaccine efficacy against PPE, especially in pigs at early life when the intestinal microbiome is more susceptible to perturbation and plays a crucial role in immune development.
2. Materials and Methods
2.1. Animals, Housing and Experimental Design
All the piglets used in this experiment were provided by the Chungnam Livestock Research Institute of South Korea (Chungcheongnam-do, South Korea), where pigs are raised free of important swine diseases, including porcine reproductive and respiratory syndrome, foot and mouth disease, and African swine fever. The piglets showed no signs of enteric or respiratory diseases when the animals were obtained for this study. A total of 64 crossbred (Duroc × [Landrace × Yorkshire]) healthy weanling pigs weaned at 4 weeks of age were housed at the Animal Research Center of Chungnam National University, South Korea. Pigs at four weeks of age were randomly divided into four groups comprising 16 animals divided into four pens with 4 pigs per pen, using a randomized complete block design for the 42-day trial. Prior to vaccination, the pigs were allowed to acclimatize for 1 week after weaning at 4 weeks of age. Then, pigs at week 0 (5 weeks of age) in the treatment groups were orally administered with three different doses (1 dose = 2 mL) of vaccine against
L.
intracellularis (Enterisol
® Ileitis, Boehringer Ingelheim Vetmedica, Inc., Ingelheim am Rhein, Germany), namely, LAW1 (0.5 dose), LAW2 (1 dose), and LAW3 (2 dose). The
L.
intracellularis vaccine was prepared by rehydrating the lyophilized live attenuated bacteria supplied as a solid cake with a solution of liquid diluent, according to the manufacturer’s instructions, and administered directly by oral drench to each pig. The pigs in pen number 1 remained unvaccinated and served as controls (CONT). All pigs in this study had similar husbandry practices to the Animal Research Center of Chungnam National University, South Korea. Room temperature was maintained at 26 °C, and the humidity was maintained constant at 60%. The pigs were housed in pens with totally slotted floors. Each pen was installed with a one-sided self-feeder and a nipple water-feeder, and pigs were given ad libitum access to feed and water throughout the experiment. A wheat–soybean meal basal diet was formulated to meet the nutrient requirements as suggested by National Research Council (NRC) [
11]. All the pigs were fed the basal diet without any antibiotics or supplementary additives. The experimental procedures used in this study were performed in accordance with the recommendations put forth in the Guide for the Use and Care of Laboratory Animals, and were approved by the Animal Care and Use Committee of Chungnam National University. Humane animal care was practiced throughout the trial and every effort was made to minimize suffering for the piglets.
2.2. Sample Collection and Genomic DNA Extraction
A total of 123 fresh fecal samples were collected directly from the rectum of each animal at week 0 (5 weeks of age, n = 63) and week 6 (11 weeks of age, n = 60) of the experiment after oral administration of the L. intracellularis vaccine. We were not able to collect a fecal sample from one piglet in LAW1 (0.5 dose) group at week 0. Before this experiment ended at week 6, two piglets from each LAW1 group (0.5 dose) and LAW3 group (2 dose) were dead, resulting in 60 samples at week 6. A total of 200 mg of feces per sample was used to extract the total DNA representing the pig gut microbial communities, using QIAamp Fast DNA Stool Mini Kit (QIAGEN, Hilden, Germany) with minor modifications. Briefly, two rounds of bead-beating the samples for 2 min at 300× g was performed with a 5 min incubation in a water bath at 70 °C between beatings. The Colibri Microvolume Spectrometer (Titertek Berthold, Pforzheim, Germany) was used to measure the concentrations of DNA, and DNAs with OD260/280 ratios of 1.80–2.15 were used in this study.
2.3. 16S rRNA Gene PCR
The V5 to V6 hypervariable regions of the 16S rRNA genes were amplified, using the universal PCR primers 799F-mod6 (5′ CMGGATTAGATACCCKGGT-3′) and 1114R (5′-GGGTTGCGCTCGTTGC-3′). The PCR amplification mixture in a reaction volume of 50 µL contained 10 pmol of each primer, 25 ng of DNA, 2.5 mM concentrations of each deoxynucleotide triphosphates (dNTPs), 2.5 U/µL of PrimeSTAR HS DNA Polymerase, and 5X PrimeSTAR Buffer (Mg2+) (Takara Bio, Inc., Shiga, Japan). The PCR cycling parameters included an initial 3 min denaturation at 98 °C, followed by 30 cycles of 98 °C for 10 s, 55 °C for 15 s, and 72 °C for 30 s, and a final 3 min extension at 72 °C. The Wizard® SV Gel and PCR Clean-Up System purification kit (Promega, Madison, WI, USA) were used to purify the PCR products.
2.4. 16S rRNA Gene Library Preparation and MiSeq Sequencing
The Illumina MiSeq platform at Macrogen Inc. (Seoul, Korea) was used to sequence 16S rRNA gene amplicons. Briefly, random fragmentation of the DNA samples followed by 5′ and 3′ adapter ligation was conducted to prepare the sequencing library. During this step, dual indices and Illumina sequencing adapters were attached to the 16S rRNA gene amplicons, using the Nextera XT Index Kit (Illumina, San Diego, CA, USA). The concentration of final products was normalized and pooled, using PicoGreen (Turner BioSystems, Inc., Sunnyvale, CA, USA), then the verification of the size of libraries was conducted using the TapeStation DNA ScreenTape D1000 (Agilent Technologies, Inc., Santa Clara, CA, USA). The PCR cycle conditions included a 3 min initial denaturation at 95 °C, 8 amplification cycles (95 °C for 30 s, 55 °C for 30 s, and 72 °C for 30 s) and a 5 min final elongation at 72 °C.
2.5. 16S rRNA Gene Analysis
The quality control of the raw sequence data generated from the Illumina MiSeq platform was performed. The sequences with a length of less than 200 bp and containing ambiguous base calls were eliminated from the demultiplexed sequence reads to minimize the effects of random sequencing errors. The UCHIME algorithm implemented in Mothur was used to identify the chimeric sequences, and those chimeric sequences were excluded for downstream analysis. Then, the operational taxonomic unit (OTU) picking using the open-reference OTU picking workflow with the SortMeRNA and SUMACLUST methods for reference OTU and de novo OTU picking were conducted, using the QIIME (Quantitative Insights into Microbial Ecology) pipeline (version 1.9.1) [
12]. The naïve Bayesian Ribosomal Database Project (RDP) classifier based on GreenGenes taxonomy reference database version 13_8 was used for the taxonomic assignment of sequence reads. For the downstream analysis, low-abundance OTUs and singletons were eliminated from the OTU table with minimum count of 4 and low-count filter based on 20% prevalence in the samples. Then, normalization of the data was conducted by rarefying the data to the minimum library size and by data scaling, using the total sum scaling (TSS) before any statistical comparison to address the sparsity of the data and the variability in the sampling depth.
2.6. Statistical Analysis
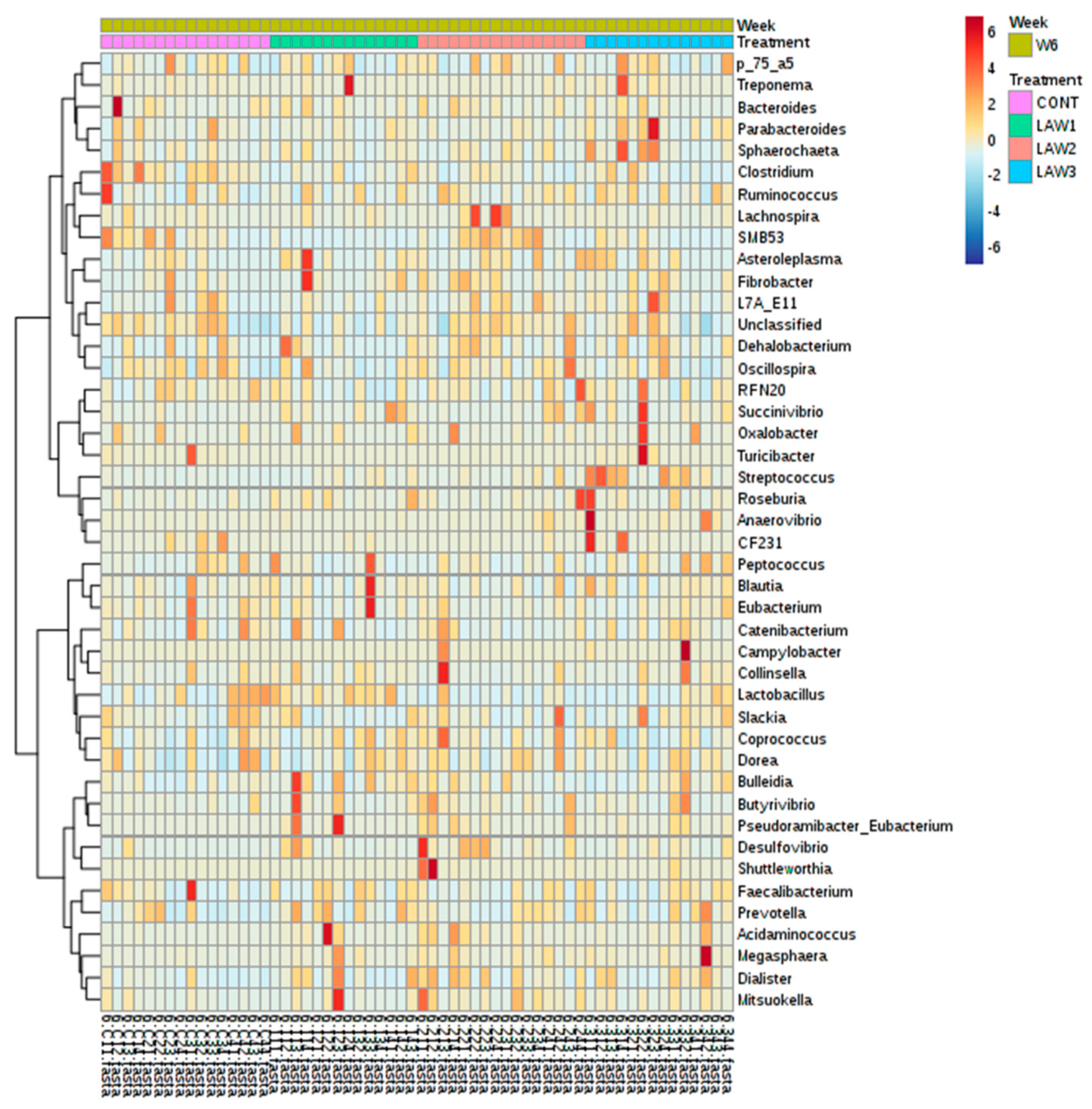
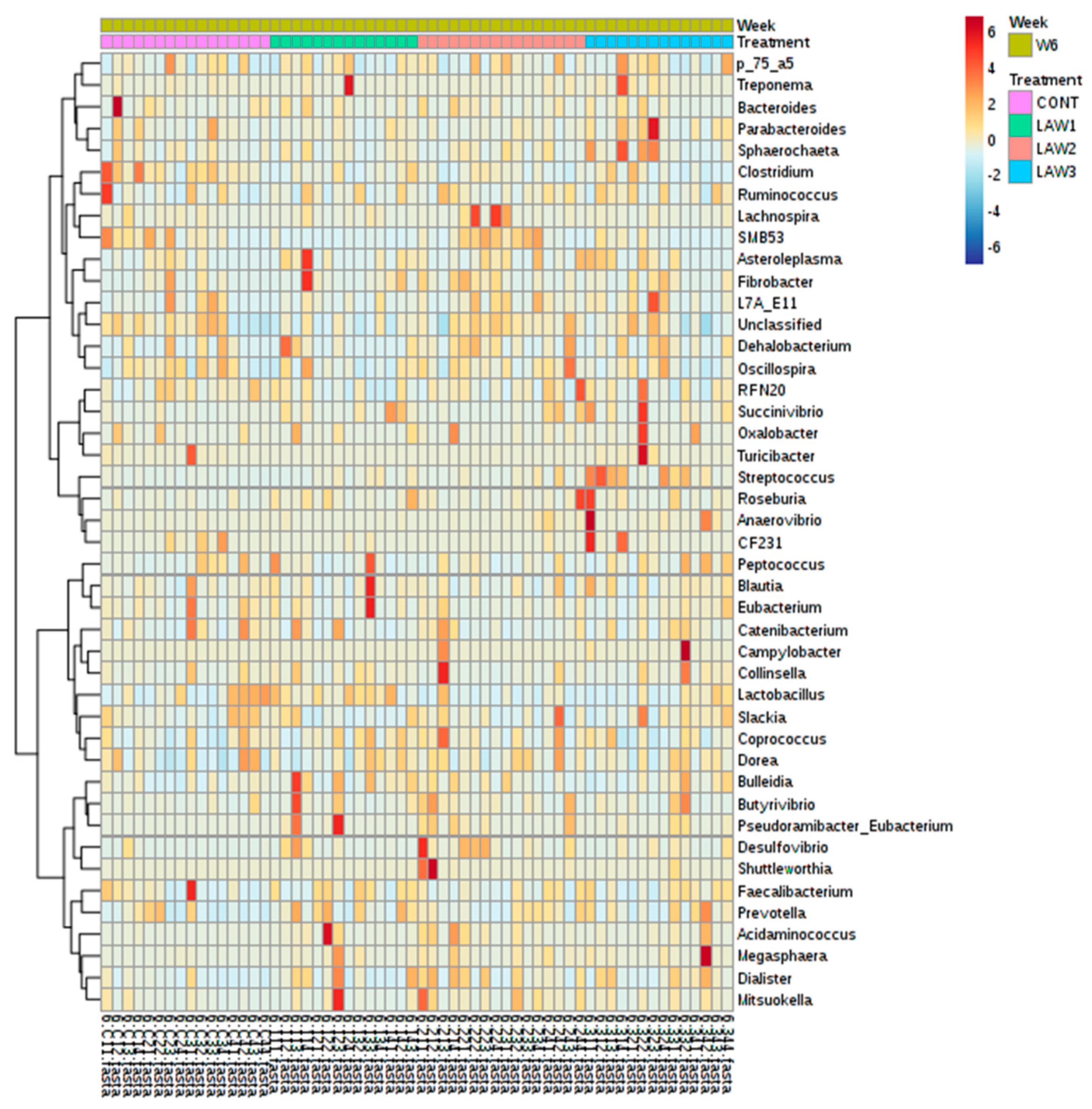
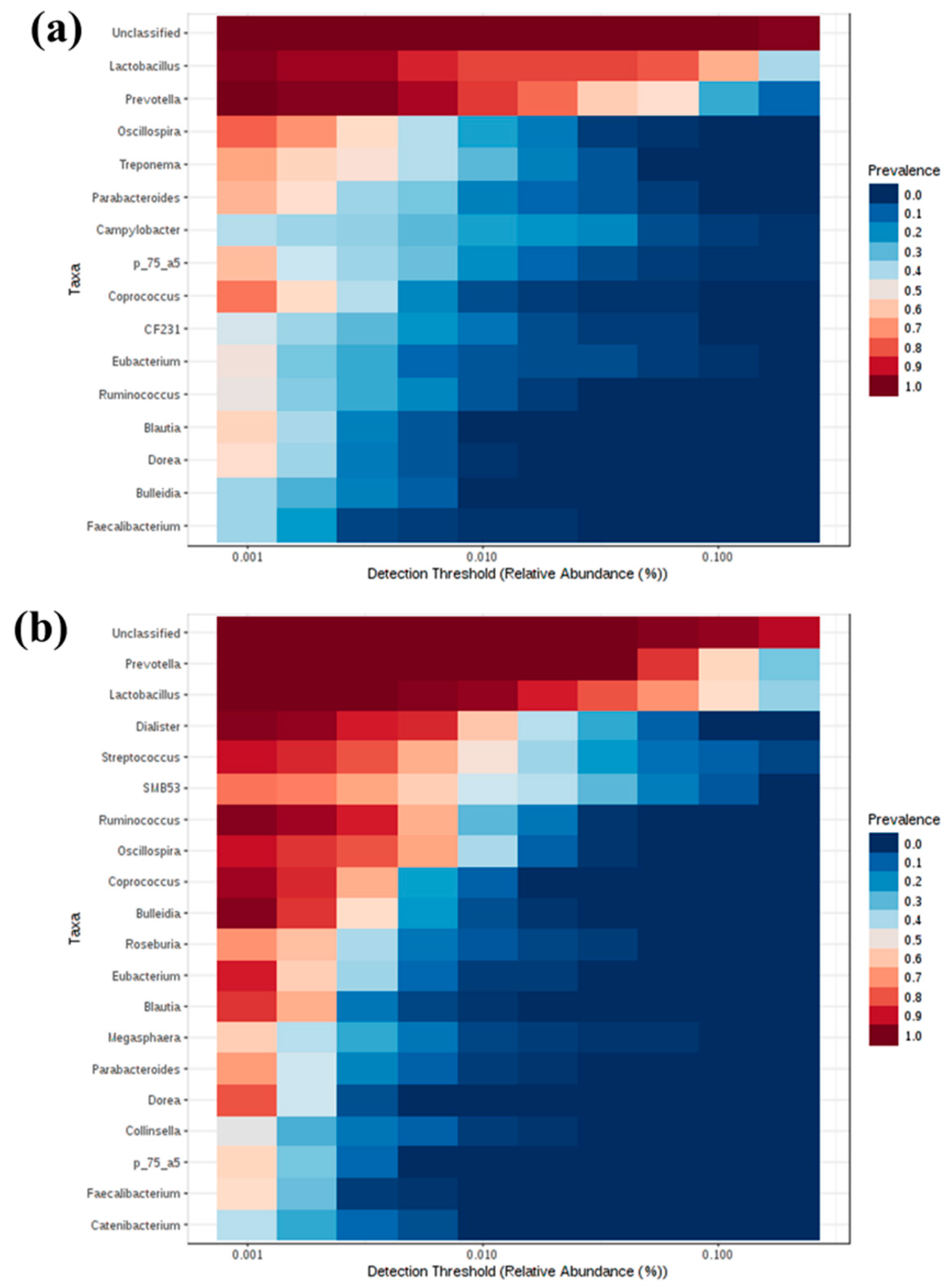
The R package MicrobiomeAnalystR and GraphPad Prism v7.00 (La Jolla, CA, USA) was used to conduct the statistical analysis. The MicrobiomeAnalystR was used to compute alpha diversity measures, including Chao1, observed OTUs, Shannon and Simpson diversity indices. Comparison of alpha diversity among the groups was conducted using the non-parametric Kruskal–Wallis test. The significant difference level was set at p < 0.05. The principal coordinate analysis (PCoA) plots at the OTU level were generated, using the weighted and unweighted UniFrac distances. Comparison of beta-diversity among the groups was conducted, using the Analysis of Similarities (ANOSIM) based on the unweighted and weighted UniFrac distances. The default parameters at 20% sample prevalence and 0.2% relative abundance were used to generate the heatmap of the core microbiota. The Euclidean distance measure and Ward clustering algorithm were used to generate the heatmap of hierarchical clustering. To elucidate the bacterial taxa that are associated with each group, the linear discriminant analysis (LDA) Effect Size (LEfSe) at the OTU level based on the non-parametric Kruskal–Wallis sum–rank test with a log LDA score threshold of 2.0 was used. A p-Value < 0.05 was considered significant.
4. Discussion
This study was performed to determine the effects of oral vaccination against L. intracellularis on the intestinal microbiome in weaned piglets. To do this, pigs were experimentally vaccinated with different doses of L. intracellularis vaccine and the differences in the microbial composition between two time points at week 0 and week 6 post vaccination were measured in order to determine the shifts in the intestinal microbiota.
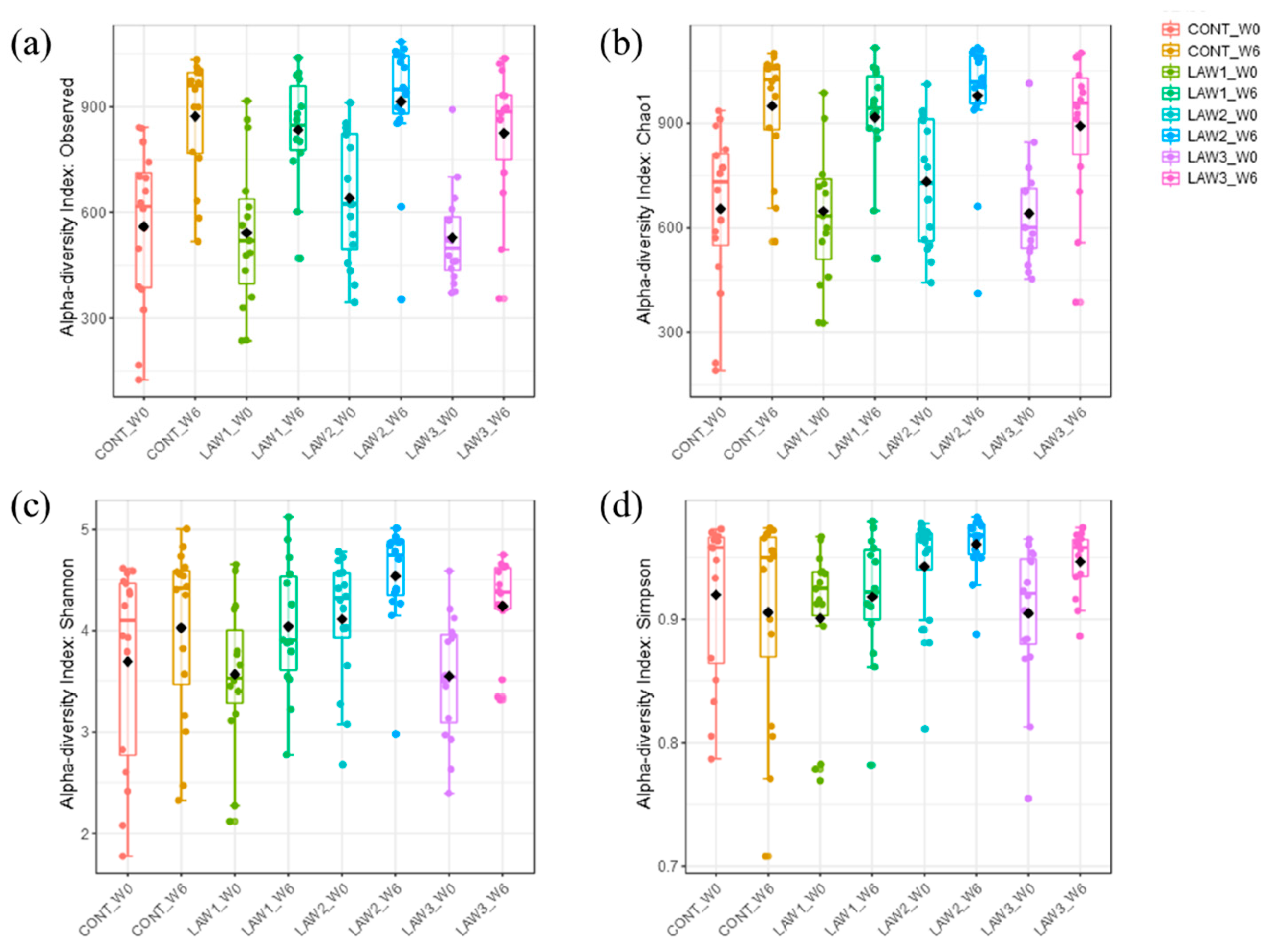
The work presented here demonstrated alterations in the composition of the gut microbiomes of pigs experimentally vaccinated with different doses of live attenuated vaccine against L. intracellularis. Alpha diversity analysis revealed that vaccination against L. intracellularis led to a significant increase in the Simpson diversity index, suggesting that significant changes in the alpha diversity were only observed in measures of species evenness and not species richness in pigs. However, no statistical differences were observed with Chao1, the number of observed OTUs, or the Shannon index at both week 0 and week 6. This also indicates that the number of species in the intestine of pigs was not altered as much as their quantities, as influenced by oral vaccination against L. intracellularis in weaned piglets.
Moreover, previous studies showed that oral challenge with
L.
intracellularis led to changes in the composition of the swine microbiome [
13]. In this study, we determined whether oral vaccination with different doses of live attenuated vaccine against
L.
intracellularis would change the microbial community structure in weaned piglets. Beta-diversity was measured using the weighted UniFrac distance, which considers the relative abundance of OTUs in the community, and the unweighted UniFrac distance, which takes into account the community membership (presence or absence of OTUs) [
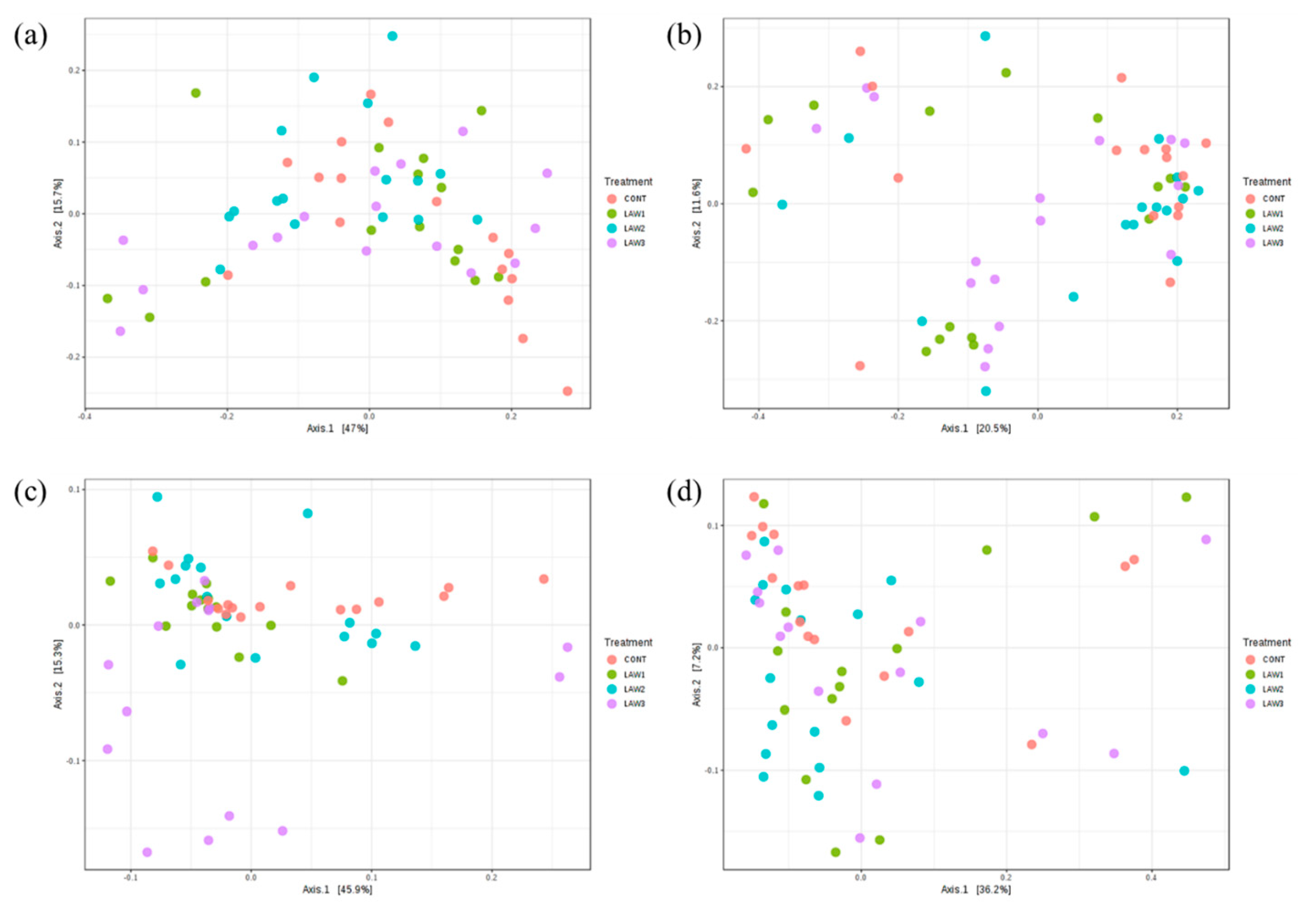
14]. The weighted and unweighted UniFrac distances were used to perform PCoA analysis to evaluate the community structure differences among the various treatment groups over the two time points. In this study, the overall compositions of the microbiomes were significantly separated by different doses of
L.
intracellularis vaccination with LAW 3 clustered distinctly, indicating that a higher dosage of
L.
intracellularis vaccine could induce alterations of the pig gut microbial community in weaned piglets. In addition, clustering of the microbiome compositions in non-vaccinated controls were segregated in different parts of the PCoA. However, the PCoA plot based on the unweighted UniFrac showed that all pigs had a similar microbiome composition regardless of the vaccination treatment (
p > 0.05). Meanwhile, microbial community structures were significantly different regardless of
L.
intracellularis vaccination between week 0 and week 6 (
p < 0.01), indicating that the intestinal microbiome of the pigs changed as the pigs aged, with an acquired microbial community composition similar to that of the adult pig gut microbiota.
We then analyzed the beta diversity separately at each time point to investigate only the effects of
L.
intracellularis vaccination on the microbial community structure among the treatment groups. At week 0, no significant differences in beta diversity were observed, suggesting that
L.
intracellularis vaccination did not have a significant impact on the microbial community structure and membership at the initial time point of the study. At week 6, it was possible to observe that
L.
intracellularis vaccination in pigs had a significant impact on the microbial community structure in pigs with a higher vaccine dose, which led to increased significant separation of the microbiota in comparison to the non-vaccinated pigs. Our findings were similar to the observations of Leite et al. [
9]; however, the advantage of this study was that significant differences observed in microbial community structure and composition were clearly due to
L.
intracellularis vaccination since we did not dually infect the pigs with other bacteria, such as
S.
enterica. Overall, our results indicate that the overall composition and structure of the pig gut microbiome was altered by oral vaccination against
L.
intracellularis.
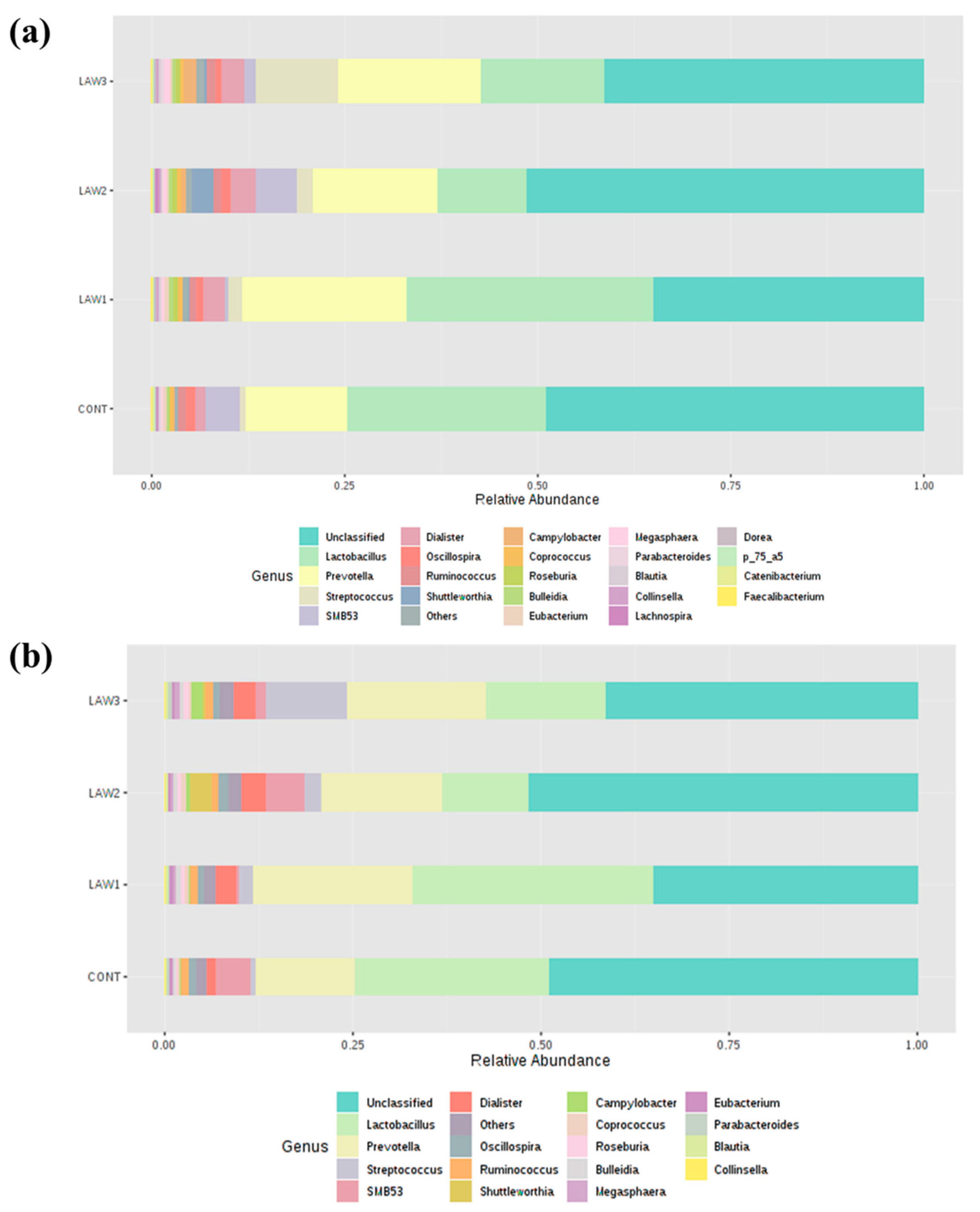
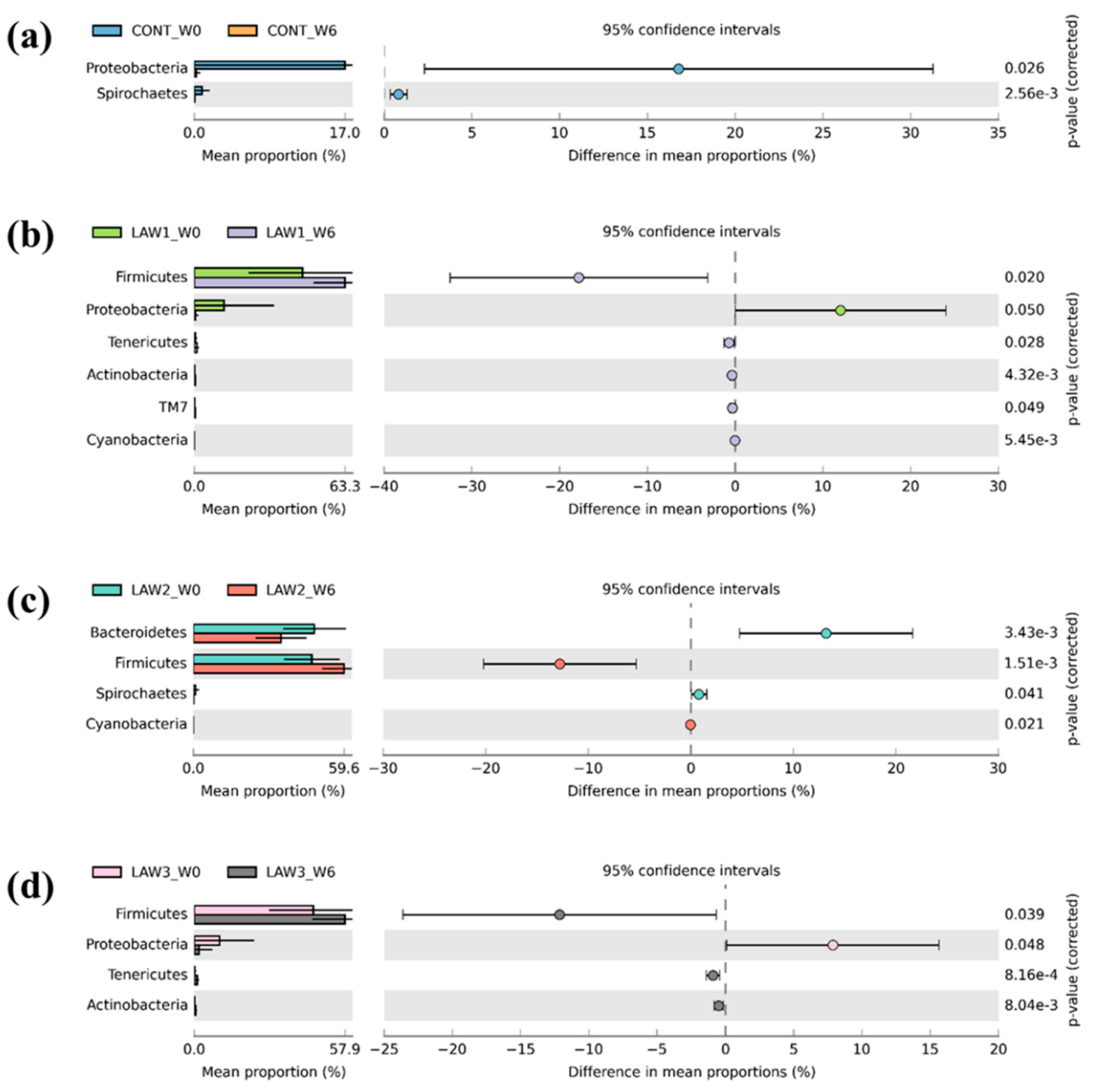
To further understand how the oral vaccination of
L.
intracellularis mediated the microbiome shifts in weaned piglets, we looked for differences in the relative abundance of differentially abundant taxa at the phylum and genus level among the treatment groups. It is known that
L.
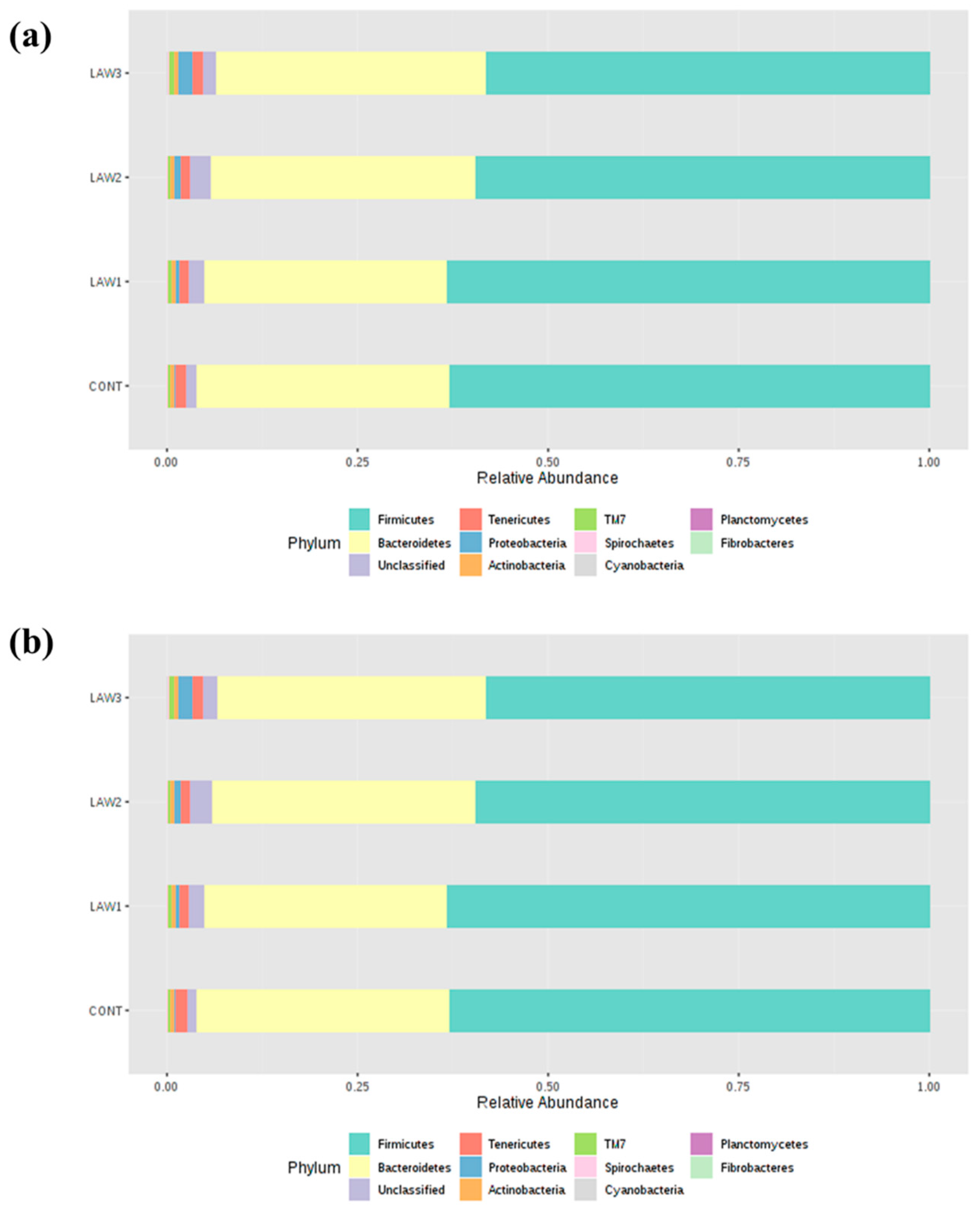
intracellularis infection can lead to changes in the intestinal microbiome of pigs, indicating complex association. In this study, taxonomic analysis at the phylum revealed that Firmicutes and Bacteroidetes were the most abundant phyla in the intestinal microbiota of piglets regardless of treatment group and time points, which accounted for more than 80% of the bacterial community, in line with other studies on pig gut microbiota [
15,
16]. Strikingly, a noticeable finding of this study was a significantly higher relative abundance of phylum Firmicutes by oral administration of
L.
intracellularis vaccine, regardless of the dose as compared to non-vaccinated controls. This finding is similar to those obtained by Leite et al. [
9], which showed higher abundance of
Firmicutes in the pig intestinal microbiome in response to infection with
L.
intracellularis. Conventionally, in piglets, the relative abundance of Firmicutes decreases and that of Bacteroidetes increases with an increase in age [
17]. Moreover, it is known that the gut microbiota of pigs early in life is more amenable to changes, hence it can be speculated that alteration in the composition of the microbiome observed in this study may be associated with oral vaccination against
L.
intracellularis.
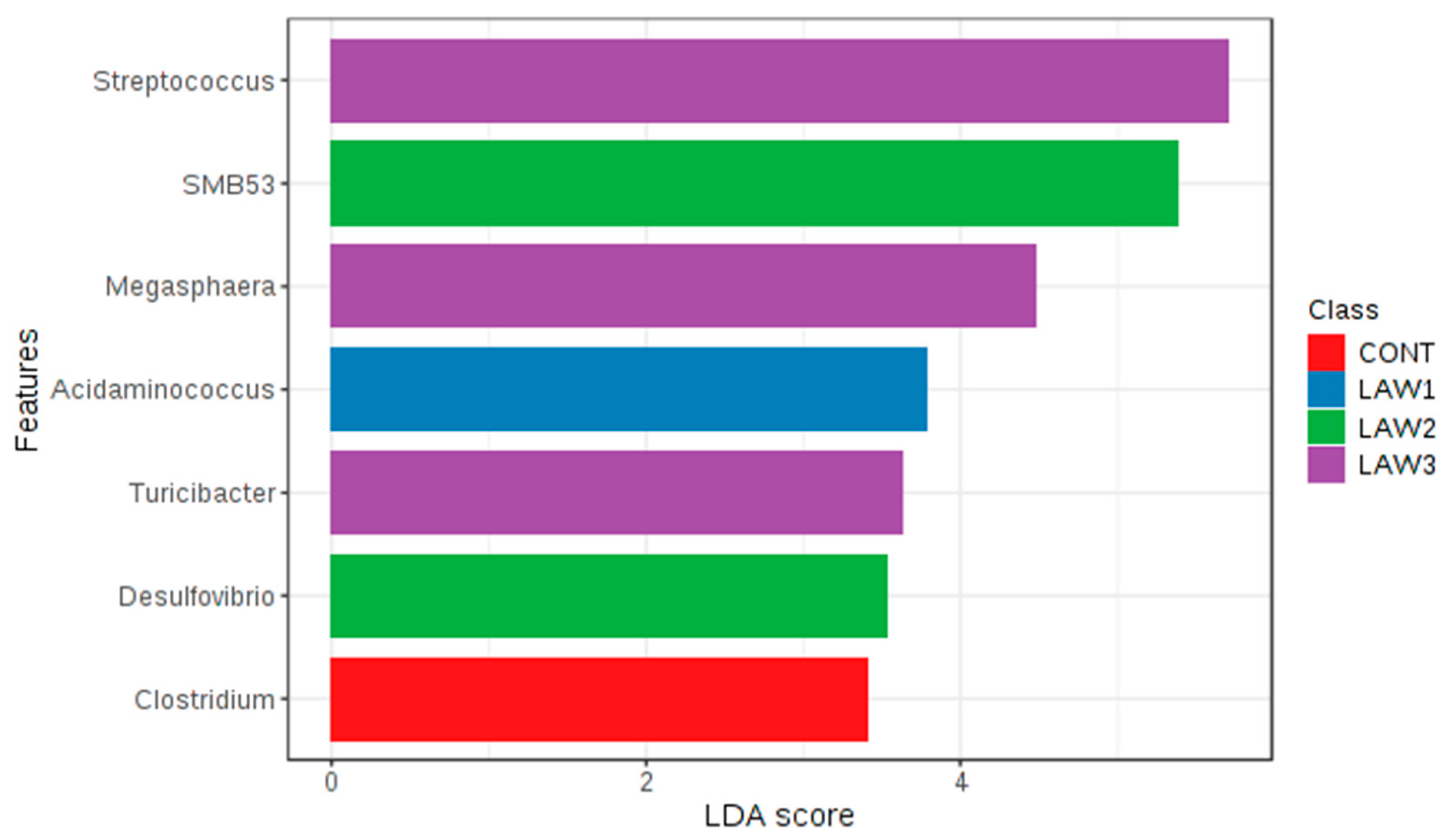
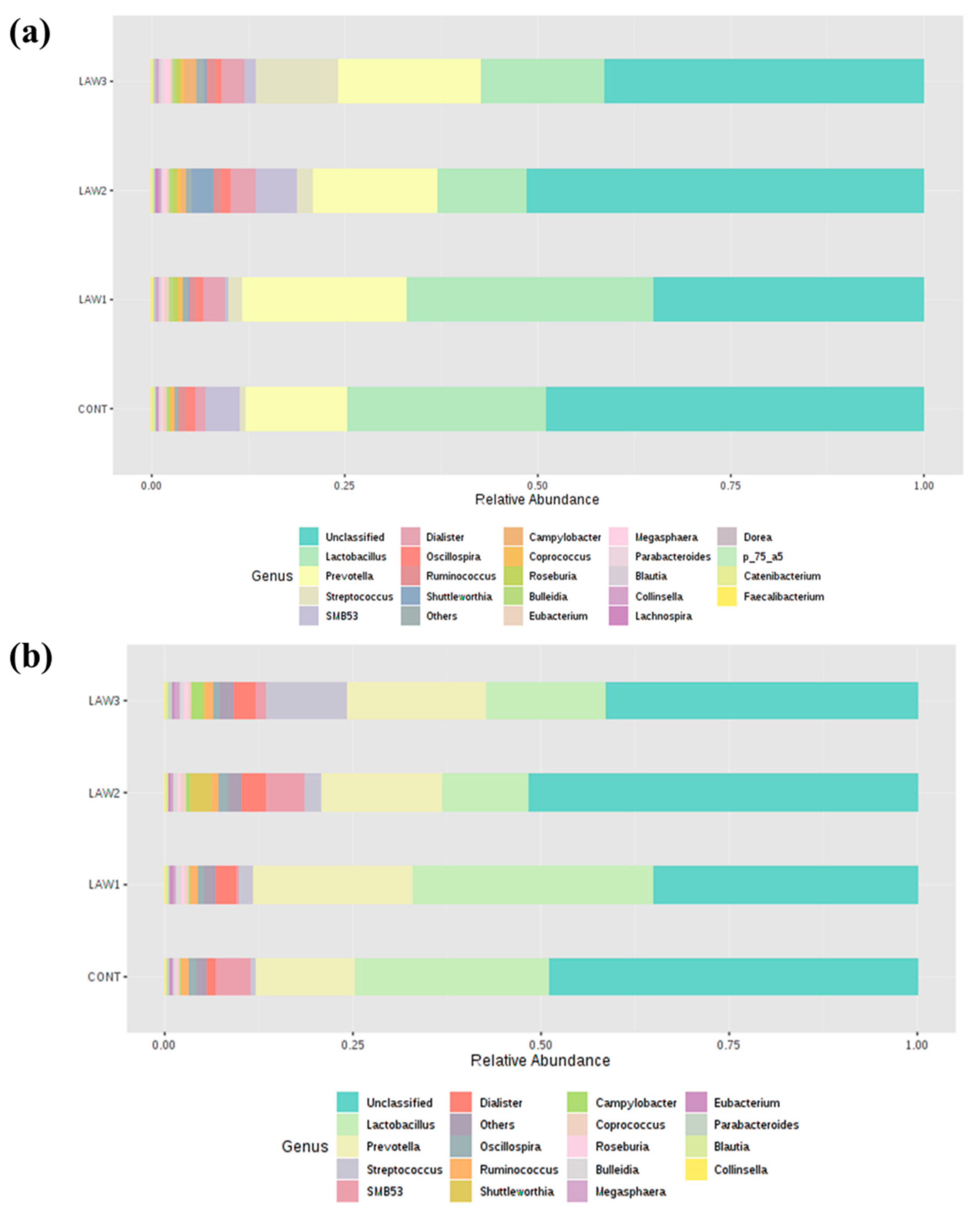
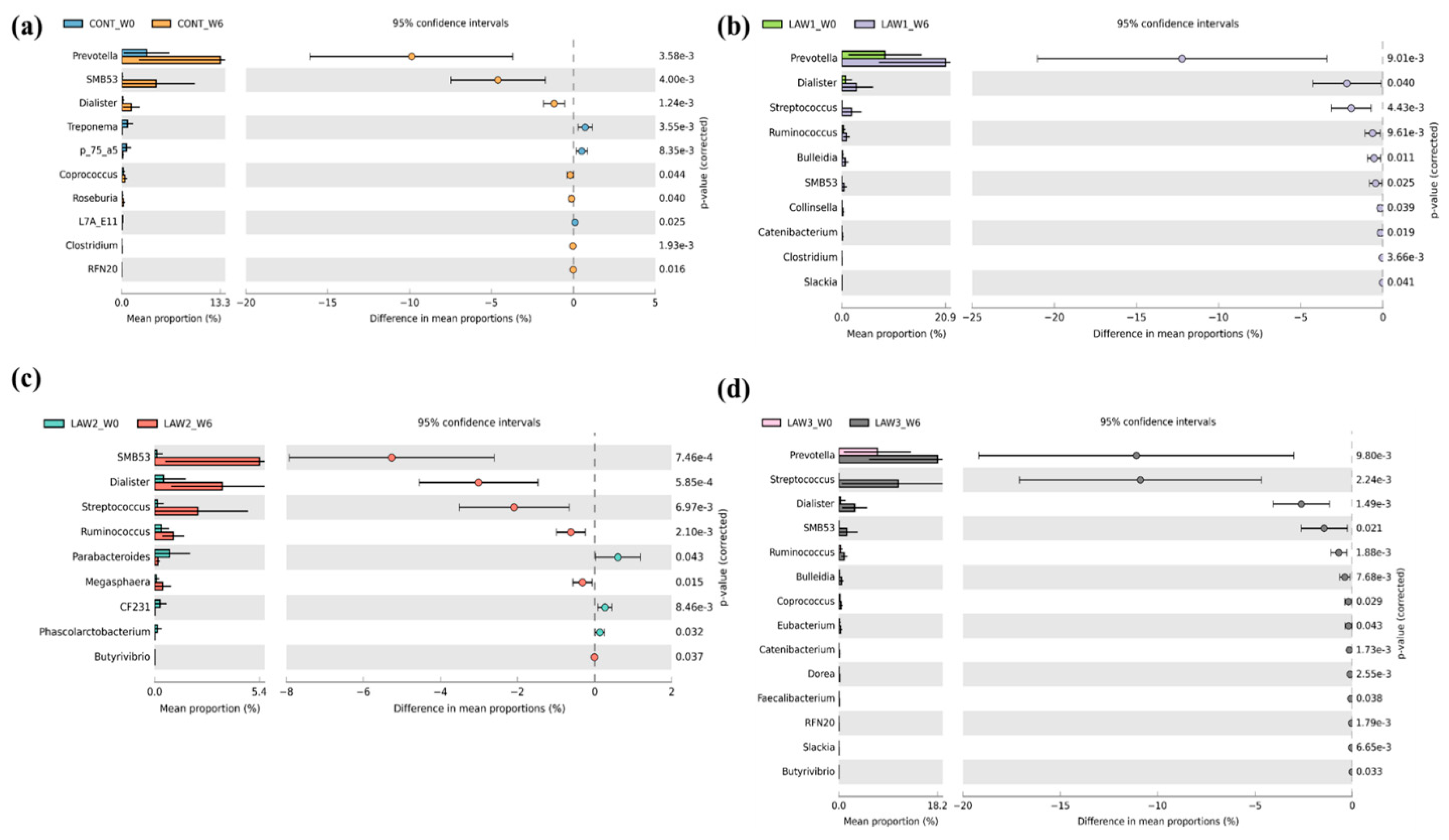
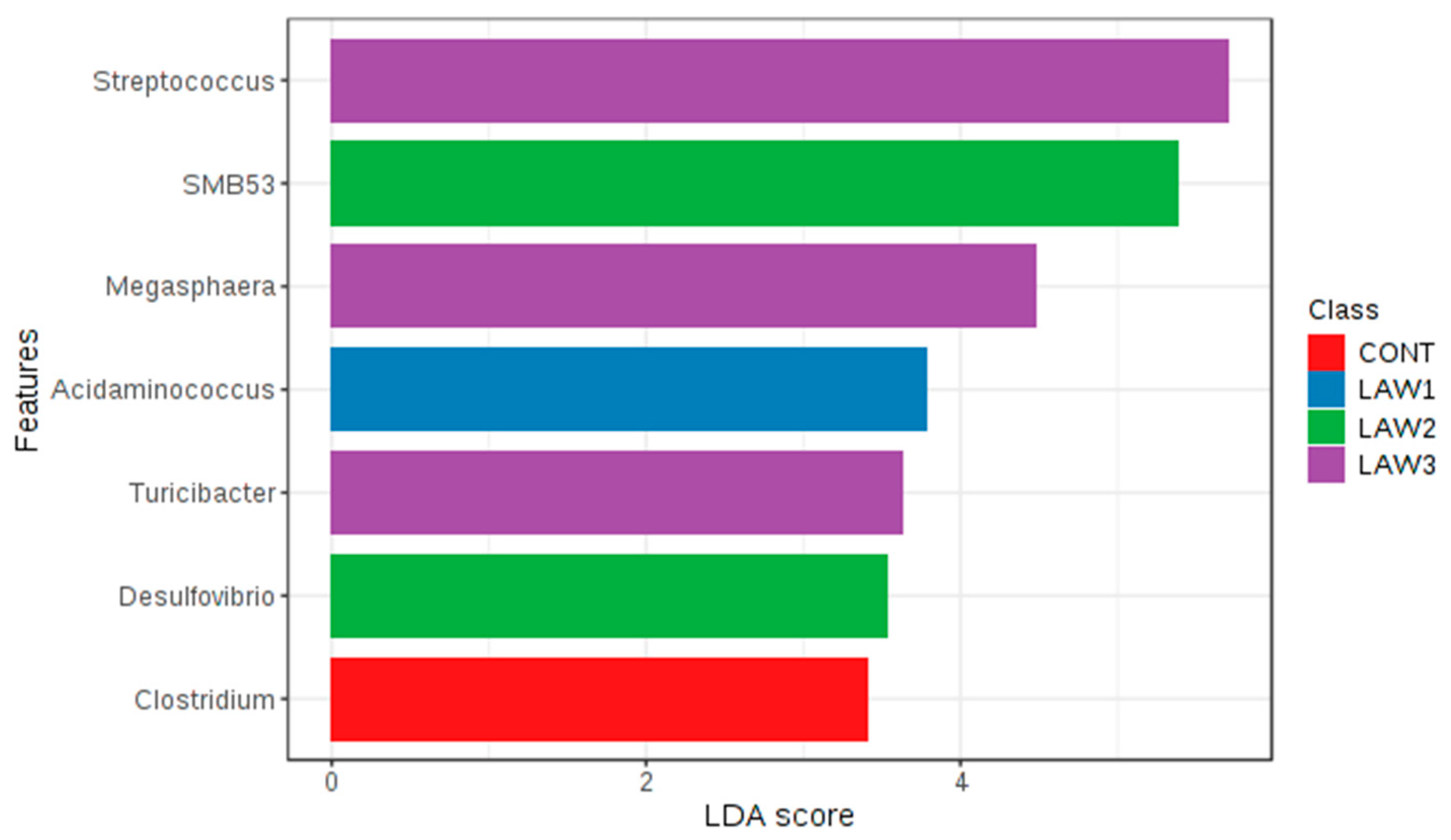
At the genus level, one of the most striking observations in this study was the significant increase in
Streptococcus and significant decrease in
Clostridium in the fecal microbiota of pigs that received oral vaccination against
L.
intracellularis regardless of dose. Interestingly, our results are similar with those of Borewicz et al., who reported that
L.
intracellularis infection could increase the level of
Streptococcus in the jejunal and ileal tissues of pigs vaccinated at 9 weeks of age [
13]. These findings indicate that similar changes in the gut microbiome were observed whether the pigs were experimentally vaccinated with a live attenuated vaccine or naturally infected with
L.
intracellularis [
13]. Previous studies revealed that some members of
Streptococcus are potentially harmful and beneficial in the intestine of pigs. For example,
Streptococcus suis is a notable pathogen in the pig industry, causing a wide variety of diseases, including meningitis, septicemia and many other infections [
18]. However, it is uncertain whether the
Streptococcus observed in this study is a harmful or beneficial bacteria. Moreover, we observed a significant decrease in the abundance of the
Clostridium species in the intestinal microbiota of pigs that received oral vaccination of
L.
intracellularis. In a similar study, the relative abundance of
Clostridium butyricum was significantly decreased in pigs dually vaccinated with
S.
enterica and
L.
intracellularis [
9]. Some
Clostridium spp. have been shown to modulate the colonic luminal metabolome by producing short-chain fatty acids, including butyrate, which can help in maintaining the gut health [
19]. Butyrate and its derivatives are reported to have positive effects on animal production, including the control of enteric pathogens, reduction in inflammation and modulation of gut microbiota [
20]. Therefore, depletion of the
Clostridium species abundance could promote and benefit the growth of pathobiont species in the pig intestinal microbiota. On the other hand, we also can say that the depletion of the
Clostridium species could also benefit pig health because some
Clostridium species are classified as pathogens. Overall, our results suggest that the significant increase in the abundance of
Streptococcus and decrease in
Clostridium due to
L.
intracellularis vaccination may play a role in maintaining pig health; however, further studies are required to determine how
L.
intracellularis vaccination affects piglet performance and gut health through pig gut microbiome modulation.
To date, very few studies have evaluated the effects of vaccination on the microbiome in pigs. Meanwhile, in human gut microbiota studies, colonization by specific bacteria, such as
Bifidobacterium, may improve vaccine response later in life, while other bacteria may lower vaccine response due to intestinal dysbiosis [
21,
22]. Previous studies provided clear evidence that the gut microbiota plays an important role in driving host health and vaccine response in pigs. For example, Munyaka et al. investigated the early-life fecal microbiota and transcriptome profiles in whole blood after vaccination for
Mycoplasma hyopneumoniae and revealed that gene expression and gut microbiota profiles could predict the vaccine response in piglets [
23]. In addition, a similar study by Leite et al. indicated that vaccination against
L.
intracellularis may be used as a promising tool to prevent
S.
enterica in swine production [
9]. In this study, some light has been shed into the potential contribution of early-life gut microbiota in the modulation of health and disease in response to vaccination against
L.
intracellularis in weaned piglets.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}