
Supplementing Garlic Nanohydrogel Optimized Growth, Gastrointestinal Integrity and Economics and Ameliorated Necrotic Enteritis in Broiler Chickens Using a Clostridium perfringens Challenge Model

 , ,
, ,  ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
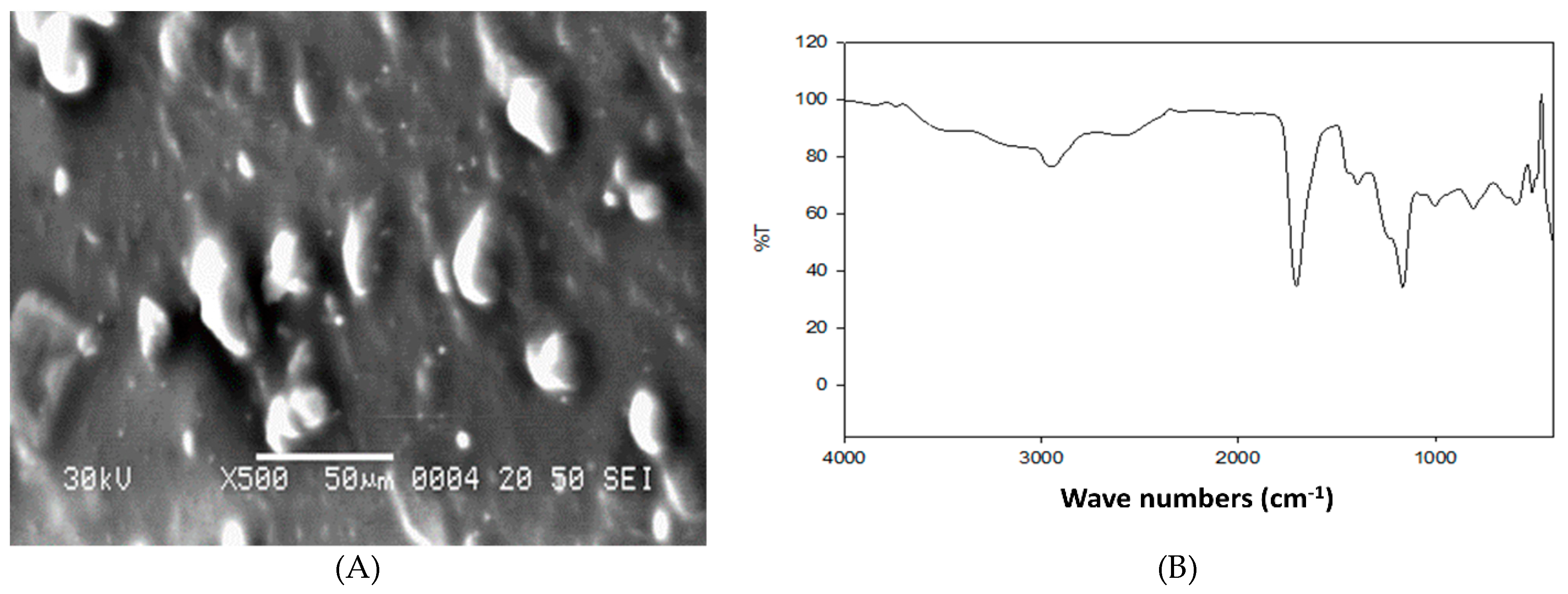
2.1. Garlic Nano-Hydrogel Preparation
2.2. Birds, Diets and Experimental Design
2.3. Monitoring Chicken’s Growth Performance
2.4. Clostridium Perfringens Challenge Model
2.5. Expression Analysis by Reverse Transcription Quantitative Real-Time PCR (RT-qPCR)
2.6. Biochemical Indices
2.7. Intestinal Lesion Score
2.8. Quantitative Detection of Ileal C. perfringens by Real-Time PCR
2.9. Economic Analysis
2.10. Statistical Analysis
3. Results
3.1. Growth Performance Parameters
3.2. Gene Expression Analysis
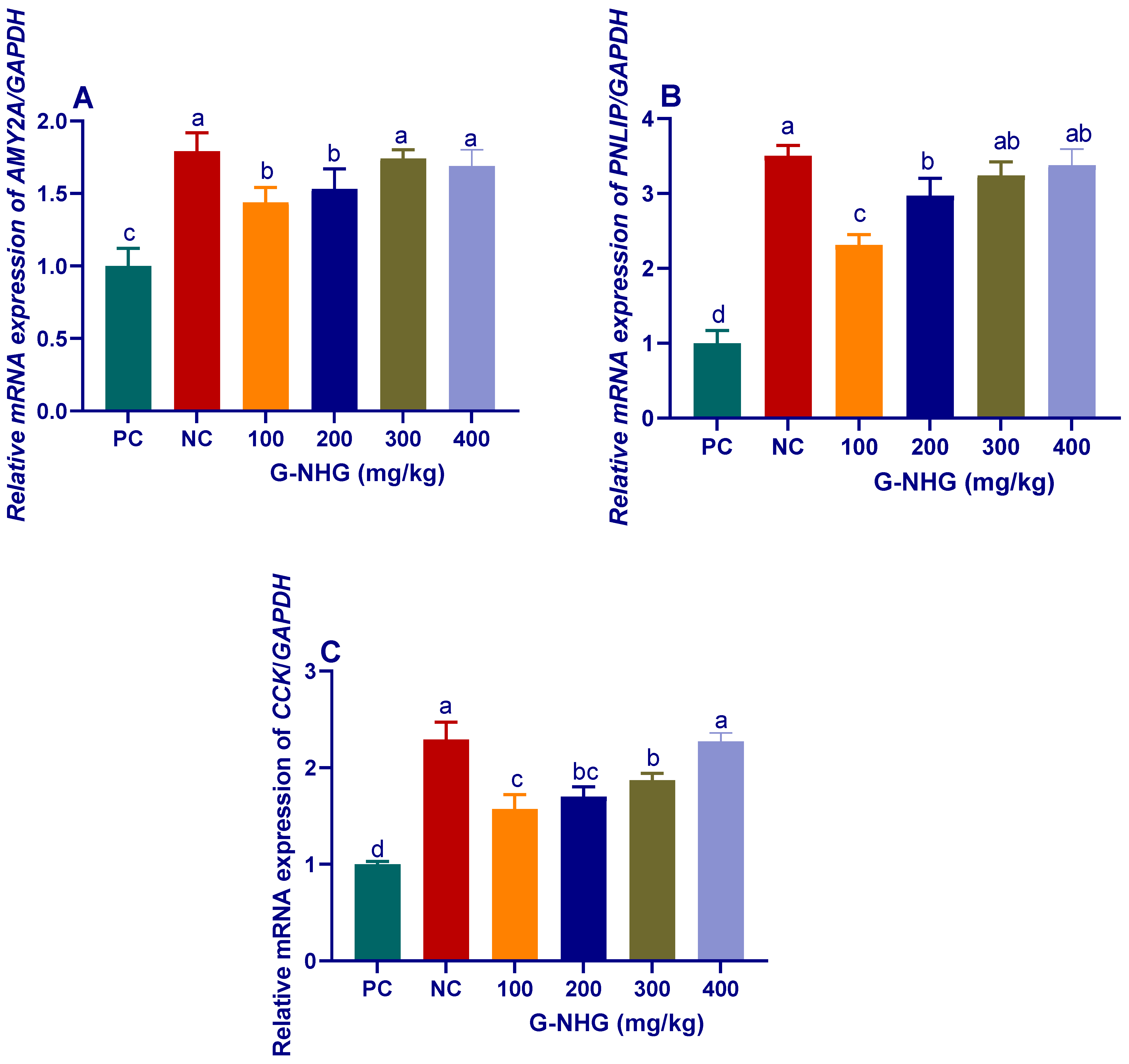
3.2.1. Digestive Enzymes
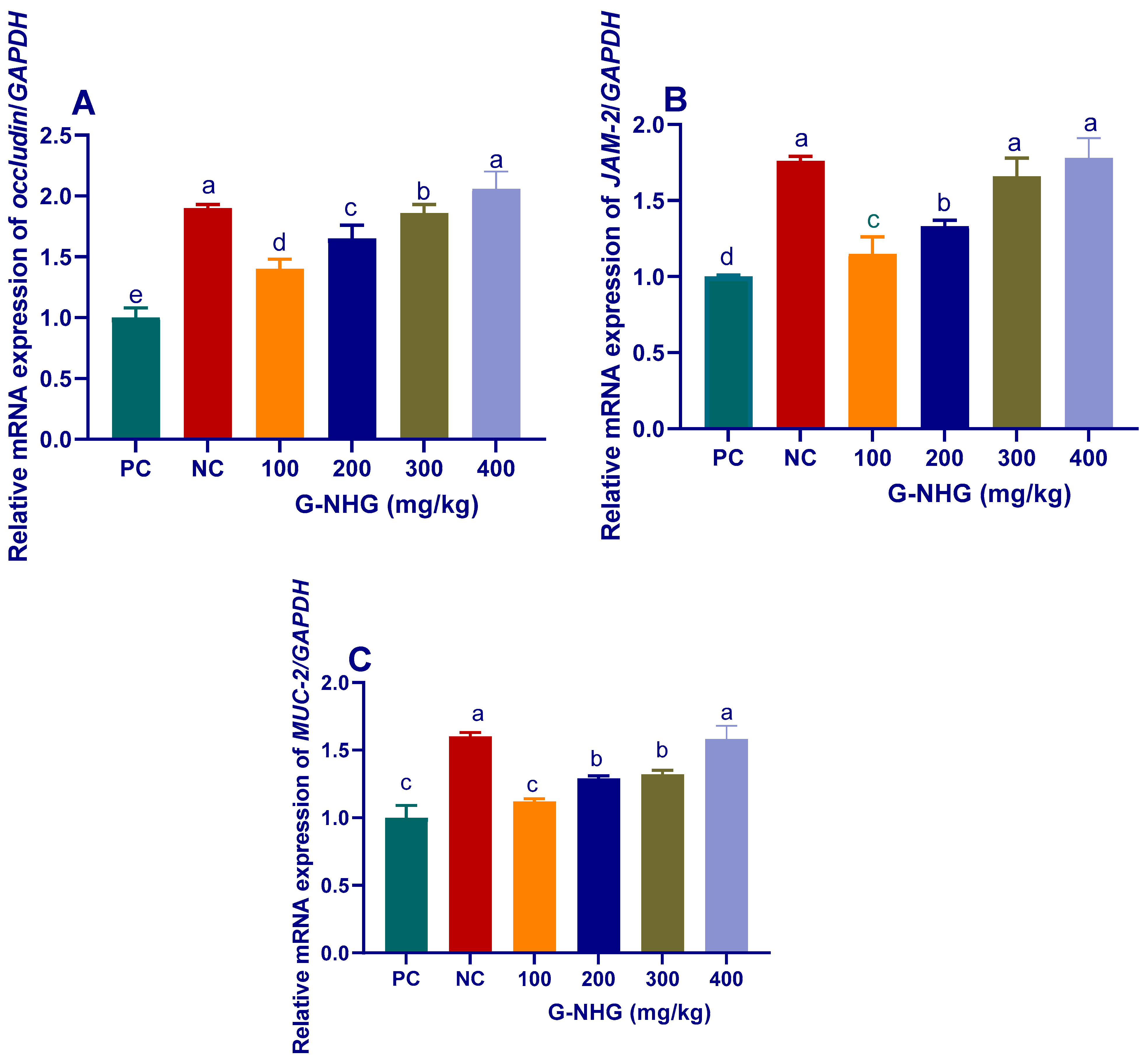
3.2.2. Tight Junction Proteins and Mucin-2
3.3. Serum Activities of Lysozyme, Nitric Oxide and Myeloperoxidase
3.4. Serum Biochemical Parameters
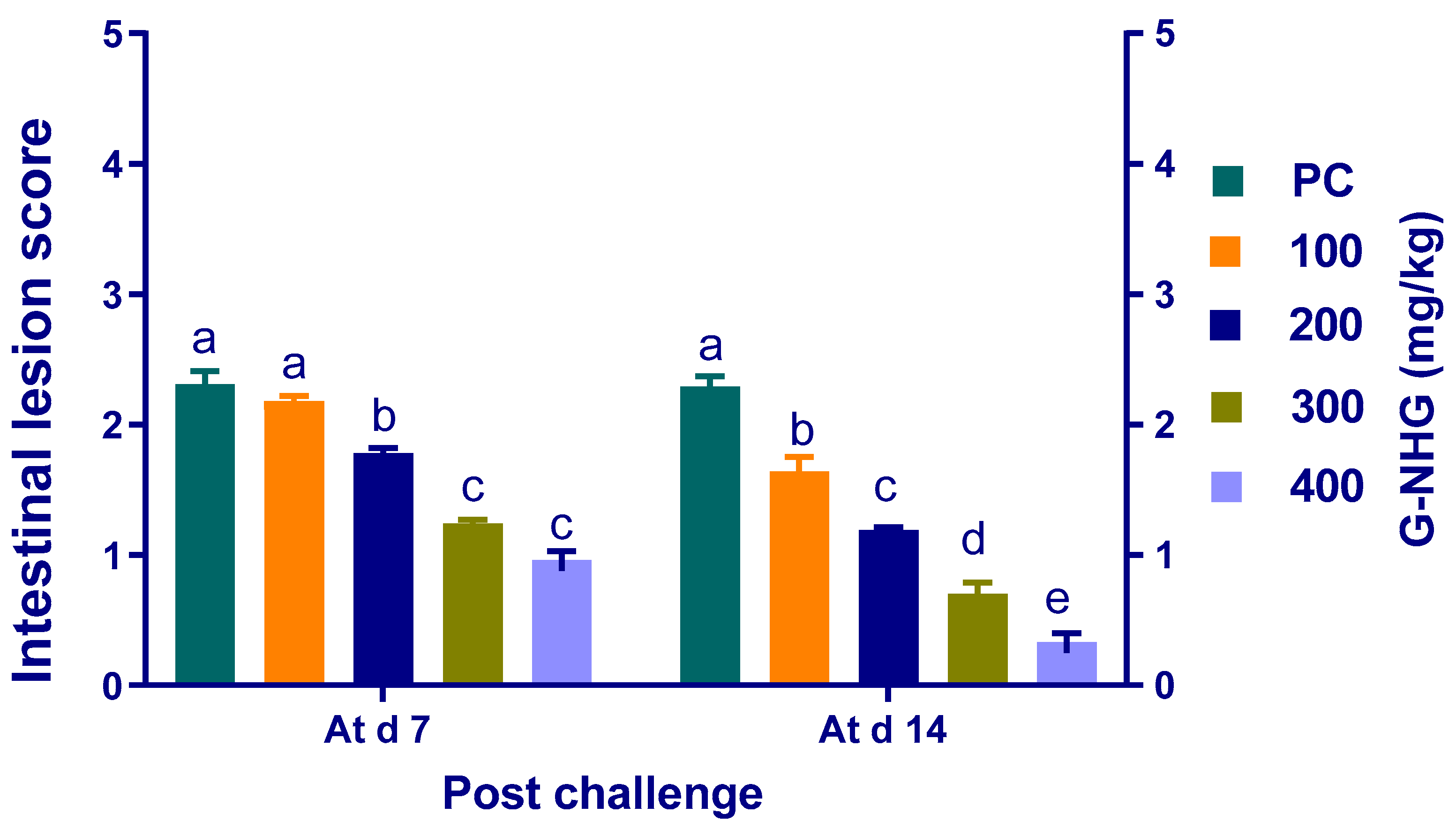
3.5. Intestinal Lesion Scoring
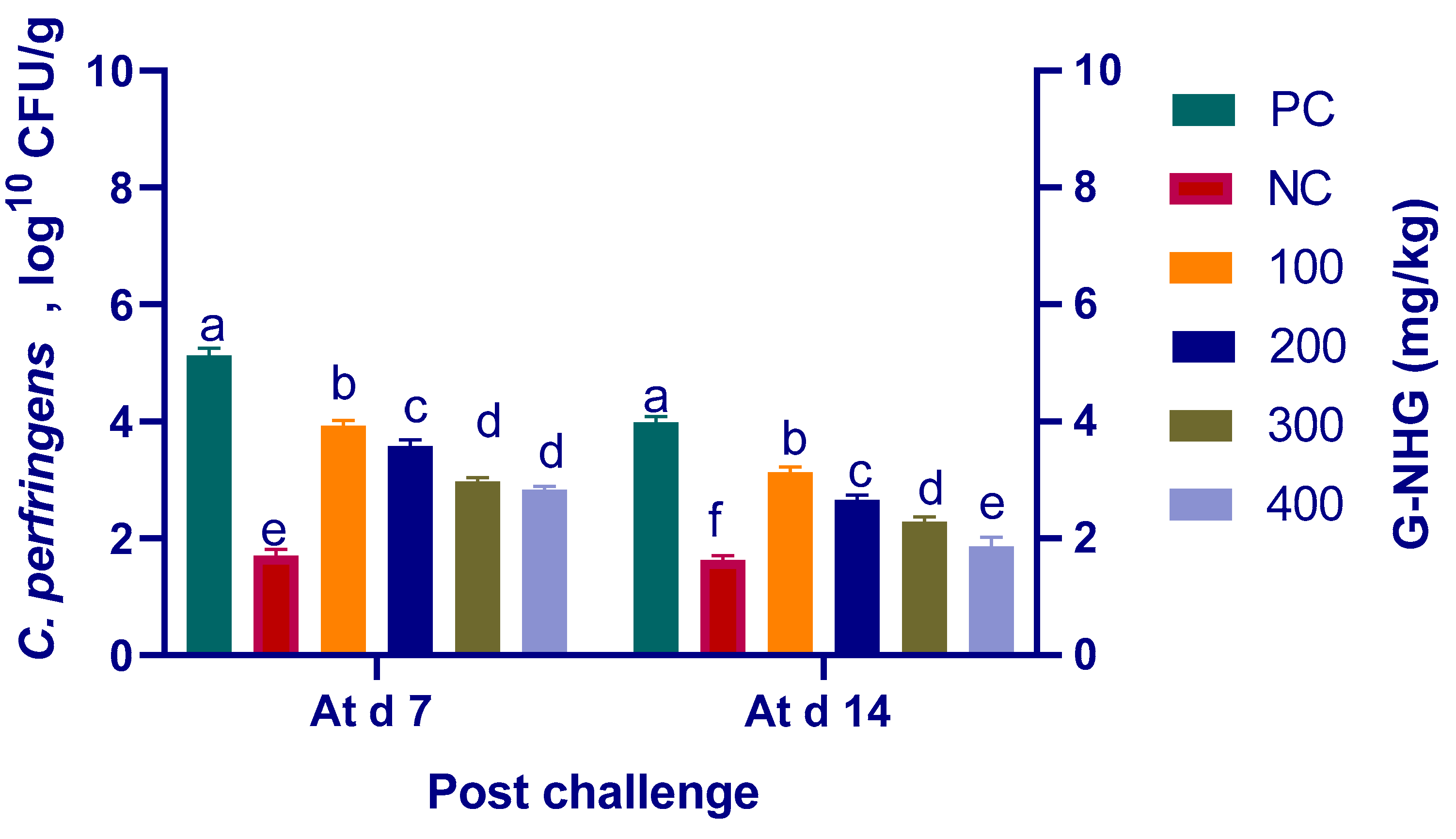
3.6. Quantitation of Ileal C. perfringens
3.7. Economic Evaluation Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Hamid, A.; Marwa, I.; El-Aziz, A.; Norhan, K.; Samir, M.; El-Naenaeey, E.-S.Y.; Abo Remela, E.M.; Mosbah, R.A.; Bendary, M.M. Genetic diversity of Campylobacter jejuni isolated from avian and human sources in Egypt. Front. Microbiol. 2019, 10, 2353. [Google Scholar] [CrossRef]
- Abdelhamied, M.I.I. Insights into antimicrobial resistance and virulence genes of emergent multidrug resistant avian pathogenic Escherichia coli in Egypt: How closely related are they? France 2015, 166, 304–314. [Google Scholar]
- Young, R.; Craig, A. Too Hard to Swallow: The Truth about Drugs and Poultry. In The Use and Misuse of Antibiotics in UK Agriculture-Part 3: Residues of Dangerous Drugs in Intensively Produced Chicken Meat and Eggs; Soil Association: Bristol, UK, 2001. [Google Scholar]
- Wade, B.; Keyburn, A. The true cost of necrotic enteritis. World Poult. 2015, 31, 16–17. [Google Scholar]
- McDevitt, R.; Brooker, J.; Acamovic, T.; Sparks, N. Necrotic enteritis; a continuing challenge for the poultry industry. Worlds Poult. Sci. J. 2006, 62, 221–247. [Google Scholar] [CrossRef]
- Rood, J.I.; Adams, V.; Lacey, J.; Lyras, D.; McClane, B.A.; Melville, S.B.; Moore, R.J.; Popoff, M.R.; Sarker, M.R.; Songer, J.G. Expansion of the Clostridium perfringens toxin-based typing scheme. Anaerobe 2018, 53, 5–10. [Google Scholar] [CrossRef]
- Bryant, A.E.; Stevens, D.L. The pathogenesis of gas gangrene. In The Clostridia; Elsevier: Amsterdam, The Netherlands, 1997; pp. 185–196. [Google Scholar]
- Prescott, J.F.; Parreira, V.R.; Mehdizadeh Gohari, I.; Lepp, D.; Gong, J. The pathogenesis of necrotic enteritis in chickens: What we know and what we need to know: A review. Avian Pathol. 2016, 45, 288–294. [Google Scholar] [CrossRef]
- Takehara, M.; Takagishi, T.; Seike, S.; Ohtani, K.; Kobayashi, K.; Miyamoto, K.; Shimizu, T.; Nagahama, M. Clostridium perfringens α-toxin impairs innate immunity via inhibition of neutrophil differentiation. Sci. Rep. 2016, 6, 28192. [Google Scholar] [CrossRef]
- Liu, D.; Guo, Y.; Wang, Z.; Yuan, J. Exogenous lysozyme influences Clostridium perfringens colonization and intestinal barrier function in broiler chickens. Avian Pathol. 2010, 39, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Du, E.; Gan, L.; Li, Z.; Wang, W.; Liu, D.; Guo, Y. In vitro antibacterial activity of thymol and carvacrol and their effects on broiler chickens challenged with Clostridium perfringens. J. Anim. Sci. Biotechnol. 2015, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Li, C.; Liu, D.; Guo, Y. Inflammatory responses to a Clostridium perfringens type A strain and α-toxin in primary intestinal epithelial cells of chicken embryos. Avian Pathol. 2015, 44, 81–91. [Google Scholar] [CrossRef]
- Du, E.; Wang, W.; Gan, L.; Li, Z.; Guo, S.; Guo, Y. Effects of thymol and carvacrol supplementation on intestinal integrity and immune responses of broiler chickens challenged with Clostridium perfringens. J. Anim. Sci. Biotechnol. 2016, 7, 19. [Google Scholar] [CrossRef]
- Wei, H.-K.; Xue, H.-X.; Zhou, Z.; Peng, J. A carvacrol–thymol blend decreased intestinal oxidative stress and influenced selected microbes without changing the messenger RNA levels of tight junction proteins in jejunal mucosa of weaning piglets. Animal 2017, 11, 193–201. [Google Scholar] [CrossRef]
- Abd El-Hamid, M.I.; El-Sayed, M.; Ali, A.R.; Abdallah, H.; Arnaout, M.I.; El-Mowalid, G.A. Marjoram extract down-regulates the expression of Pasteurella multocida adhesion, colonization and toxin genes: A potential mechanism for its antimicrobial activity. Comp. Immunol. Microbiol. Infect. Dis. 2019, 62, 101–108. [Google Scholar] [CrossRef]
- Ibrahim, D.; Abdelfattah-Hassan, A.; Badawi, M.; Ismail, T.A.; Bendary, M.M.; Abdelaziz, A.M.; Mosbah, R.A.; Mohamed, D.I.; Arisha, A.H.; El-Hamid, M.I.A. Thymol nanoemulsion promoted broiler chicken’s growth, gastrointestinal barrier and bacterial community and conferred protection against Salmonella typhimurium. Sci. Rep. 2021, 11, 7742. [Google Scholar] [CrossRef]
- Brenes, A.; Roura, E. Essential oils in poultry nutrition: Main effects and modes of action. Anim. Feed Sci. Technol. 2010, 158, 1–14. [Google Scholar] [CrossRef]
- Placha, I.; Takacova, J.; Ryzner, M.; Cobanova, K.; Laukova, A.; Strompfova, V.; Venglovska, K.; Faix, S. Effect of thyme essential oil and selenium on intestine integrity and antioxidant status of broilers. Br. Poult. Sci. 2014, 55, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Wlodarska, M.; Willing, B.P.; Bravo, D.M.; Finlay, B.B. Phytonutrient diet supplementation promotes beneficial Clostridia species and intestinal mucus secretion resulting in protection against enteric infection. Sci. Rep. 2015, 5, 9253. [Google Scholar] [CrossRef]
- Kim, D.; Lillehoj, H.; Lee, S.; Jang, S.; Bravo, D. High-throughput gene expression analysis of intestinal intraepithelial lymphocytes after oral feeding of carvacrol, cinnamaldehyde, or Capsicum oleoresin. Poult. Sci. 2010, 89, 68–81. [Google Scholar] [CrossRef]
- Corzo-Martínez, M.; Corzo, N.; Villamiel, M. Biological properties of onions and garlic. Trends Food Sci. Technol. 2007, 18, 609–625. [Google Scholar] [CrossRef]
- Sarica, S.; Ciftci, A.; Demir, E.; Kilinc, K.; Yildirim, Y. Use of an antibiotic growth promoter and two herbal natural feed additives with and without exogenous enzymes in wheat based broiler diets. S. Afr. J. Anim. Sci. 2005, 35, 61–72. [Google Scholar] [CrossRef]
- Brzóska, F.; Sliwinski, B.; Michalik-Rutkowska, O.; Sliwa, J. The effect of garlic (Allium sativum L.) on growth performance, mortality rate, meat and blood parameters in broilers. Ann. Anim. Sci. 2015, 15, 961. [Google Scholar] [CrossRef][Green Version]
- Kırkpınar, F.; Ünlü, H.B.; Özdemir, G. Effects of oregano and garlic essential oils on performance, carcase, organ and blood characteristics and intestinal microflora of broilers. Livest. Sci. 2011, 137, 219–225. [Google Scholar] [CrossRef]
- Milea, Ș.-A.; Aprodu, I.; Vasile, A.M.; Barbu, V.; Râpeanu, G.; Bahrim, G.E.; Stănciuc, N. Widen the functionality of flavonoids from yellow onion skins through extraction and microencapsulation in whey proteins hydrolysates and different polymers. J. Food Eng. 2019, 251, 29–35. [Google Scholar] [CrossRef]
- Gangadoo, S.; Stanley, D.; Hughes, R.J.; Moore, R.J.; Chapman, J. Nanoparticles in feed: Progress and prospects in poultry research. Trends Food Sci. Technol. 2016, 58, 115–126. [Google Scholar] [CrossRef]
- Amiri, N.; Afsharmanesh, M.; Salarmoini, M.; Meimandipour, A.; Hosseini, S.; Ebrahimnejad, H. Nanoencapsulation (in vitro and in vivo) as an efficient technology to boost the potential of garlic essential oil as alternatives for antibiotics in broiler nutrition. Animal 2021, 15, 100022. [Google Scholar] [CrossRef]
- Aviagen, W. Ross 308: Broiler’s Management and Nutrition Specification. 2018. Available online: https://en.aviagen.com/assets/Tech_Center/Ross_Broiler/Ross-BroilerHandbook2018-EN.pdf (accessed on 20 May 2021).
- AOAC. Official Methods of Analysis of AOAC International; Association of Official Analytical Chemists: Rockville, MD, USA, 2012. [Google Scholar]
- Al-Khalaifah, H.S.; Shahin, S.E.; Omar, A.E.; Mohammed, H.A.; Mahmoud, H.I.; Ibrahim, D. Effects of graded levels of microbial fermented or enzymatically treated dried brewer’s grains on growth, digestive and nutrient transporter genes expression and cost effectiveness in broiler chickens. BMC Vet. Res. 2020, 16, 424. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, D.; Abdelfattah-Hassan, A.; Arisha, A.H.; Abd El-Aziz, R.M.; Sherief, W.R.; Adil, S.H.; El Sayed, R.; Metwally, A.E. Impact of feeding anaerobically fermented feed supplemented with acidifiers on its quality and growth performance, intestinal villi and enteric pathogens of mulard ducks. Livest. Sci. 2020, 242, 104299. [Google Scholar] [CrossRef]
- Ibrahim, D.; Moustafa, A.; Shahin, S.; Sherief, W.; Farag, M.; Nassan, M. Impact of fermented or enzymatically fermented dried olive pomace on growth, expression of digestive enzymes and glucose transporters genes, oxidative stability of frozen meat and economic efficiency of broiler chickens. Front. Vet. Sci. 2021, 8, 442. [Google Scholar] [CrossRef]
- Yin, D.; Du, E.; Yuan, J.; Gao, J.; Wang, Y.; Aggrey, S.E.; Guo, Y. Supplemental thymol and carvacrol increases ileum Lactobacillus population and reduces effect of necrotic enteritis caused by Clostridium perfringes in chickens. Sci. Rep. 2017, 7, 7334. [Google Scholar] [CrossRef]
- Prescott, J.; Sivendra, R.; Barnum, D. The use of bacitracin in the prevention and treatment of experimentally-induced necrotic enteritis in the chicken. Can. Vet. J. 1978, 19, 181. [Google Scholar]
- Underwood, W.; Anthony, R. AVMA guidelines for the euthanasia of animals: 2020 edition. Retrieved March 2013, 30, 2001–2020. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wise, M.G.; Siragusa, G.R. Quantitative detection of Clostridium perfringens in the broiler fowl gastrointestinal tract by real-time PCR. Appl. Environ. Microbiol. 2005, 71, 3911–3916. [Google Scholar] [CrossRef]
- Williams, R. A compartmentalised model for the estimation of the cost of coccidiosis to the world’s chicken production industry. Int. J. Parasitol. 1999, 29, 1209–1229. [Google Scholar] [CrossRef]
- Pant, S.; Bhatt, P.; Shekhar, S.; Krishna, G. Evaluation of economic losses due to coccidiosis in layer birds in tarai region of Uttarakhand. Int. J. Curr. Microbiol. App. Sci. 2018, 7, 381–386. [Google Scholar] [CrossRef]
- Kirubakaran, A.; Moorthy, M.; Chitra, R.; Prabakar, G. Influence of combinations of fenugreek, garlic, and black pepper powder on production traits of the broilers. Vet. World 2016, 9, 470. [Google Scholar] [CrossRef]
- Amagase, H.; Milner, J.A. Impact of various sources of garlic and their constituents on 7, 12-dimethylbenz [α] anthracene binding to mammary cell DNA. Carcinogenesis 1993, 14, 1627–1631. [Google Scholar] [CrossRef]
- Sheoran, N.; Kumar, R.; Kumar, A.; Batra, K.; Sihag, S.; Maan, S.; Maan, N. Nutrigenomic evaluation of garlic (Allium sativum) and holy basil (Ocimum sanctum) leaf powder supplementation on growth performance and immune characteristics in broilers. Vet. World 2017, 10, 121. [Google Scholar] [CrossRef] [PubMed]
- Massad, M.A.; Ramamneh, D.; Sharafat, A.A.; Hussain, N. Effect of using garlic on the economical and physiological characteristics of broiler chickens. Int. J. Environ. Sci. Nat. Resour. 2018, 10, 54–58. [Google Scholar]
- Peinado, M.; Ruiz, R.; Echávarri, A.; Aranda-Olmedo, I.; Rubio, L. Garlic derivative PTS-O modulates intestinal microbiota composition and improves digestibility in growing broiler chickens. Anim. Feed Sci. Technol. 2013, 181, 87–92. [Google Scholar] [CrossRef]
- Van Immerseel, F.; Rood, J.I.; Moore, R.J.; Titball, R.W. Rethinking our understanding of the pathogenesis of necrotic enteritis in chickens. Trends Microbiol. 2009, 17, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, J.; Wilkie, D.; Van Kessel, A.; Drew, M. Potential strategies for controlling necrotic enteritis in broiler chickens in post-antibiotic era. Anim. Feed Sci. Technol. 2006, 129, 60–88. [Google Scholar] [CrossRef]
- Kim, D.K.; Lillehoj, H.S.; Lee, S.H.; Lillehoj, E.P.; Bravo, D. Improved resistance to Eimeria acervulina infection in chickens due to dietary supplementation with garlic metabolites. Br. J. Nutr. 2013, 109, 76–88. [Google Scholar] [CrossRef]
- Lanzotti, V.; Bonanomi, G.; Scala, F. What makes Allium species effective against pathogenic microbes? Phytochem. Rev. 2013, 12, 751–772. [Google Scholar] [CrossRef]
- Lee, Y.; Lee, S.-H.; Gadde, U.; Oh, S.; Lee, S.; Lillehoj, H. Allium hookeri supplementation improves intestinal immune response against necrotic enteritis in young broiler chickens. Poult. Sci. 2018, 97, 1899–1908. [Google Scholar] [CrossRef]
- Jimoh, A.; Ibitoye, E.; Dabai, Y.; Garba, S. In vivo antimicrobial potentials of garlic against Clostridium perfringens and its promotant effects on performance of broiler chickens. Pak. J. Biol. Sci. PJBS 2013, 16, 1978–1984. [Google Scholar] [CrossRef]
- Kothari, D.; Lee, W.-D.; Niu, K.-M.; Kim, S.-K. The genus Allium as poultry feed additive: A review. Animals 2019, 9, 1032. [Google Scholar] [CrossRef]
- Herawati, M. The effect of feeding red ginger (Zingiber officinale rosc) as phytobiotic on broiler slaughter weight and meat quality. Int. J. Poult. Sci. 2011, 10, 983–985. [Google Scholar]
- Ramakrishna Rao, R.; Platel, K.; Srinivasan, K. In vitro influence of spices and spice-active principles on digestive enzymes of rat pancreas and small intestine. Food/Nahrung 2003, 47, 408–412. [Google Scholar] [CrossRef]
- Pham, V.H.; Kan, L.; Huang, J.; Geng, Y.; Zhen, W.; Guo, Y.; Abbas, W.; Wang, Z. Dietary encapsulated essential oils and organic acids mixture improves gut health in broiler chickens challenged with necrotic enteritis. J. Anim. Sci. Biotechnol. 2020, 11, 18. [Google Scholar] [CrossRef]
- Rajput, I.; Li, L.; Xin, X.; Wu, B.; Juan, Z.; Cui, Z.; Yu, D.; Li, W. Effect of Saccharomyces boulardii and Bacillus subtilis B10 on intestinal ultrastructure modulation and mucosal immunity development mechanism in broiler chickens. Poult. Sci. 2013, 92, 956–965. [Google Scholar] [CrossRef]
- König, J.; Wells, J.; Cani, P.D.; García-Ródenas, C.L.; MacDonald, T.; Mercenier, A.; Whyte, J.; Troost, F.; Brummer, R.-J. Human intestinal barrier function in health and disease. Clin. Transl. Gastroenterol. 2016, 7, e196. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Gadde, U.D.; Oh, S.; Lee, Y.; Davis, E.; Zimmerman, N.; Rehberger, T.; Lillehoj, H.S. Dietary Bacillus subtilis-based direct-fed microbials alleviate LPS-induced intestinal immunological stress and improve intestinal barrier gene expression in commercial broiler chickens. Res. Vet. Sci. 2017, 114, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Farahat, M.; Ibrahim, D.; Kishawy, A.; Abdallah, H.; Hernandez-Santana, A.; Attia, G. Effect of cereal type and plant extract addition on the growth performance, intestinal morphology, caecal microflora, and gut barriers gene expression of broiler chickens. Animal 2021, 15, 100056. [Google Scholar] [CrossRef] [PubMed]
- Forder, R.; Nattrass, G.; Geier, M.; Hughes, R.; Hynd, P. Quantitative analyses of genes associated with mucin synthesis of broiler chickens with induced necrotic enteritis. Poult. Sci. 2012, 91, 1335–1341. [Google Scholar] [CrossRef]
- Awad, W.A.; Hess, C.; Hess, M. Enteric pathogens and their toxin-induced disruption of the intestinal barrier through alteration of tight junctions in chickens. Toxins 2017, 9, 60. [Google Scholar] [CrossRef]
- Wang, H.; Liang, S.; Li, X.; Yang, X.; Long, F.; Yang, X. Effects of encapsulated essential oils and organic acids on laying performance, egg quality, intestinal morphology, barrier function, and microflora count of hens during the early laying period. Poult. Sci. 2019, 98, 6751–6760. [Google Scholar] [CrossRef]
- Amiri, N.; Afsharmanesh, M.; Salarmoini, M.; Meimandipour, A.; Hosseini, S.; Ebrahimnejad, H. Effects of nanoencapsulated cumin essential oil as an alternative to the antibiotic growth promoter in broiler diets. J. Appl. Poult. Res. 2020, 29, 875–885. [Google Scholar] [CrossRef]
- Prasad, R.; Rose, M.; Virmani, M.; Garg, S.; Puri, J. Lipid profile of chicken (Gallus domesticus) in response to dietary supplementation of garlic (Allium sativum). Int. J. Poult. Sci. 2009, 8, 270–276. [Google Scholar] [CrossRef]
- Qureshi, A.A.; Din, Z.; Abuirmeileh, N.; Burger, W.; Ahmad, Y.; Elson, C. Suppression of avian hepatic lipid metabolism by solvent extracts of garlic: Impact on serum lipids. J. Nutr. 1983, 113, 1746–1755. [Google Scholar] [CrossRef] [PubMed]
- Eidi, A.; Eidi, M.; Esmaeili, E. Antidiabetic effect of garlic (Allium sativum L.) in normal and streptozotocin-induced diabetic rats. Phytomedicine 2006, 13, 624–629. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Jin, S.; Yang, H. Effect of dietary garlic bulb and husk on the physicochemical properties of chicken meat. Poult. Sci. 2009, 88, 398–405. [Google Scholar] [CrossRef]
- Omar, A.E.; Al-Khalaifah, H.S.; Ismail, T.A.; El-Aziz, A.; Reda, M.; El-Mandrawy, S.A.; Shalaby, S.I.; Ibrahim, D. Performance, serum biochemical and immunological parameters, and digestive enzyme and intestinal barrier-related gene expression of broiler chickens fed fermented fava bean by-products as a substitute for conventional feed. Front. Vet. Sci. 2021, 8, 740. [Google Scholar]
- Awad, N.F.; Abd El-Hamid, M.I.; Hashem, Y.; Erfan, A.; Abdelrahman, B.; Mahmoud, H.I. Impact of single and mixed infections with Escherichia coli and Mycoplasma gallisepticum on Newcastle disease virus vaccine performance in broiler chickens: An in vivo perspective. J. Appl. Microbiol. 2019, 127, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Aziz, N.K.; Abd El-Hamid, M.I.; Bendary, M.M.; El-Azazy, A.A.; Ammar, A.M. Existence of vancomycin resistance among methicillin resistant S. aurues recovered from animal and human sources in Egypt. Slov. Vet. Res. 2018, 55, 221–230. [Google Scholar]
- Abd El-Hamid, M.I.; Bendary, M. Comparative phenotypic and genotypic discrimination of methicillin resistant and susceptible Staphylococcus aureus in Egypt. Cell. Mol. Biol. 2015, 61, 101–112. [Google Scholar]
- Elsom, G.K. An antibacterial assay of aqueous extract of garlic against anaerobic/microaerophilic and aerobic bacteria. Microb. Ecol. Health Dis. 2000, 12, 81–84. [Google Scholar]
- Ross, Z.; O’Gara, E.A.; Hill, D.J.; Sleightholme, H.; Maslin, D.J. Antimicrobial properties of garlic oil against human enteric bacteria: Evaluation of methodologies and comparisons with garlic oil sulfides and garlic powder. Appl. Environ. Microbiol. 2001, 67, 475–480. [Google Scholar] [CrossRef]
- Katata-Seru, L.; Lebepe, T.C.; Aremu, O.S.; Bahadur, I. Application of Taguchi method to optimize garlic essential oil nanoemulsions. J. Mol. Liq. 2017, 244, 279–284. [Google Scholar] [CrossRef]
- Osman, K.M.; El-Enbaawy, M.I.; Ezzeldin, N.A.; Hussein, H.M. Nitric oxide and lysozyme production as an impact to Clostridium perfringens mastitis. Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, 505–511. [Google Scholar] [CrossRef]
- El-Gogary, M.R.; El-Khateeb, A.Y.; Megahed, A.M. Effect of physiological and chemical nano garlic supplementation on broiler chickens. Plant Arch. 2019, 19, 695–705. [Google Scholar]
- Hamza, R.Z.; Abd El-Aziz, S.A.; Said, A.A.; Khairy, M.H.; Mahmoud, S.H.; Habib, W.A.; El-Shenawy, N.S. Improving the efficacy of garlic extract in African catfish against copper sulfate-induced immunological and histological effects. Reg. Stud. Mar. Sci. 2021, 41, 101579. [Google Scholar] [CrossRef]
- Windisch, W.; Schedle, K.; Plitzner, C.; Kroismayr, A. Use of phytogenic products as feed additives for swine and poultry. J. Anim. Sci. 2008, 86, E140–E148. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, D.; Moustafa, A.; Metwally, A.S.; Nassan, M.A.; Abdallah, K.; Eldemery, F.; Tufarelli, V.; Laudadio, V.; Kishawy, A.T. Potential application of Cornelian cherry extract on broiler chickens: Growth, expression of antioxidant biomarker and glucose transport genes, and oxidative stability of frozen meat. Animals 2021, 11, 1038. [Google Scholar] [CrossRef]
- Ibrahim, D.; Sewid, A.H.; Arisha, A.H.; Abd El-Fattah, A.H.; Abdelaziz, A.M.; Al-Jabr, O.A.; Kishawy, A.T. Influence of glycyrrhiza glabra extract on growth, gene expression of gut integrity, and Campylobacter jejuni colonization in broiler chickens. Front. Vet. Sci. 2020, 7, 612063. [Google Scholar] [CrossRef] [PubMed]
- Puvača, N.; Brkić, I.; Jahić, M.; Nikolić, S.R.; Radović, G.; Ivanišević, D.; Đokić, M.; Bošković, D.; Ilić, D.; Brkanlić, S. The effect of using natural or biotic dietary supplements in poultry nutrition on the effectiveness of meat production. Sustainability 2020, 12, 4373. [Google Scholar] [CrossRef]
- Oleforuh-Okoleh, V.; Chukwu, G.; Adeolu, A. Effect of ground ginger and garlic on the growth performance, carcass quality and economics of production of broiler chickens. Glob. J. Biosci. Biotechnol. 2014, 3, 225–229. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Starter (1–10 d) | Grower (11–22 d) | Finisher (23–38 d) | |
|---|---|---|---|
| Ingredient, % | |||
| Yellow corn | 57.9 | 57.9 | 63.2 |
| Soybean meal, 48% | 35.4 | 35.4 | 28.1 |
| Corn gluten, 60% | 1.5 | 1.5 | 1.5 |
| Soybean oil | 1.40 | 1.4 | 3.7 |
| Calcium carbonate | 0.50 | 0.5 | 0.5 |
| Monocalcium phosphate | 1.40 | 1.4 | 1.2 |
| Common salt | 0.30 | 0.3 | 0.3 |
| Premix * | 0.80 | 0.8 | 0.8 |
| DL-Methionine, 98% | 0.20 | 0.20 | 0.10 |
| Lysine, Hcl, 78% | 0.30 | 0.30 | 0.20 |
| Antitoxin | 0.10 | 0.10 | 0.10 |
| Choline chloride | 0.20 | ||
| Chemical composition | |||
| ME, Kcal/Kg | 3003 | 3106 | 3200 |
| CP % | 23.02 | 21.50 | 20.00 |
| EE % | 3.88 | 5.11 | 6.25 |
| CF % | 2.66 | 2.59 | 2.52 |
| Ca % | 1.18 | 1.00 | 0.81 |
| Available P % | 0.51 | 0.47 | 0.39 |
| Lysine % | 1.43 | 1.26 | 1.15 |
| Methionine % | 0.54 | 0.50 | 0.42 |
| Gene | Primer Sequence (5′–3′) | Accession No |
|---|---|---|
| AMY2A | F-CGGAGTG↓GATGTTAACGACTGG R-ATGTTCGCAGACCCAGTCATTG | NM_001001473.2 |
| PNLIP | F-GCATCTGGGAAG↓GAACTAGGG R- TGAACCACAAGCATAGCCCA | NM_001277382.1 |
| CCK | F-AGGTTCCACTGGGAGGTTCT R-CGCCTGCTGTTCTTTAGGAG | XM_015281332.1 |
| occludin | F-ACGGCAAAGCCAACATCTAC R-ATCCGCCACGTTCTTCAC | XM_031604121.1 |
| JAM-2 | F-AGACAG GAACAGGCAGTGCT R-TCCAATCCCATTTGA GGCTA | XM_031556661.1 |
| Muc-2 | F-AAACAACGGCCATGTTTCAT R-GTGTGACACTGGTGTGCTGA | NM_001318434 |
| GAPDH | F-GGTGGTGCTAAGCGTGTTA R-CCCTCCACAATGCCAA | NM205518 |
| Group | PC | NC | G-NHG, mg/kg | p Value | SEM | ||||
|---|---|---|---|---|---|---|---|---|---|
| Parameter | 100 | 200 | 300 | 400 | |||||
| Initial body weight (g/bird) | 44.00 | 44.00 | 44.20 | 44.40 | 44.40 | 44.60 | 0.99 | 0.71 | |
| Starter (1–10 d) | |||||||||
| BW, g/bird | 320.22 d | 321.26 c,d | 321.76 c,d | 327.08 b,c | 331.22 b | 339.62 a | <0.001 | 6.03 | |
| BWG, g/bird | 275.62 d | 277.26 c,d | 277.56 c,d | 282.68 b,c | 287.22 b | 295.22 a | <0.001 | 6.29 | |
| FI, g/bird | 330.40 a | 330.82 a | 335.28 a | 330.86 a | 332.86 a | 315.08 b | <0.001 | 10.33 | |
| FCR | 1.20 b | 1.19 b | 1.21 b | 1.17 b | 1.16 b | 1.07 a | <0.001 | 0.08 | |
| Grower (11–22 d) | |||||||||
| BW, g/bird | 1208 c | 1206 c | 1230 b | 1245 b | 1287 a | 1284 a | <0.001 | 50.20 | |
| BWG, g/bird | 888 c | 885 c | 908 b | 918 b | 955 a | 944 a | <0.001 | 60.26 | |
| FI, g/bird | 1218 a | 1226 a | 1226 a | 1229 a | 1228 a | 1167 b | <0.001 | 18.54 | |
| FCR | 1.37 a | 1.38 a | 1.35 a | 1.34 a | 1.29 b | 1.23 c | <0.001 | 0.07 | |
| Finisher (23–38) | |||||||||
| BW, g/bird | 1895 d | 2402 a | 1912 d | 2075 c | 2261 b | 2482 a | <0.001 | 21.54 | |
| BWG, g/bird | 686 d | 1195 a | 682 d | 830 c | 974 b | 1198 a | <0.001 | 18.63 | |
| FI, g/bird | 2011 c | 2302 c | 1968 c | 2052 b | 2328 b | 2556 a | <0.001 | 29.78 | |
| FCR | 2.9 a | 1.93 d | 2.89 a | 2.47 b | 2.39 b | 2.13 c | <0.001 | 0.05 | |
| Overall performance (1–38 d) | |||||||||
| BWG, g/bird | 1850 e | 2358 b | 1868 e | 2031 d | 2217 c | 2438 a | <0.001 | 18.62 | |
| FI, g/bird | 3560 c | 3859 b | 3530 c | 3612 c | 3889 b | 4039 a | <0.001 | 16.25 | |
| FCR | 1.92 a | 1.63 c | 1.89 a | 1.78 b | 1.75 b | 1.66 c | <0.001 | 0.03 | |
| Group | At 7 d Post Challenge | At 14 d Post Challenge | |||||
|---|---|---|---|---|---|---|---|
| Lysozyme (U/mL) | MPO (U/L) | NO (μmol/L) | Lysozyme (U/mL) | MPO (U/L) | NO (μmol/L) | ||
| PC NC |
212.127 a 186.733 c |
32.000 a 25.300 d |
6.383 a 4.833 b |
127.033 a 72.233 f |
29.440 a 21.033 d |
6.383 a 4.833 b | |
| G-NHG, mg/kg | 100 | 199.417 b | 29.067 b | 6.033 a | 121.430 b | 27.450 b | 5.240 b |
| 200 | 191.137 b,c | 29.133 b | 5.967 a | 101.410 c | 26.453 b,c | 5.317 a,b | |
| 300 | 176.823 d | 27.513 c | 5.240 b | 87.373 d | 25.407 c | 3.940 c | |
| 400 | 171.000 d | 25.427 d | 4.243 c | 77.817 e | 22.127 d | 3.210 d | |
| P value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| SEM | 2.148 | 0.288 | 0.109 | 1.077 | 0.391 | 0.086 | |
| Group | ALT, U/L | AST, U/L | Uric acid, μmol/L | Creatinine, mg/dL | Cholesterol, mg/dL | Triglycerides, mg/dL | HDL, mg/dL | LDL, mg/dL | VLDL, mg/dL | |
|---|---|---|---|---|---|---|---|---|---|---|
| PC | 63.967 a | 51.767 a | 13.633 a | 0.580 a | 140.400 a | 108.367 b | 33.200 b | 85.567 a | 21.633 a | |
| NC | 38.800 b | 36.967 b | 11.900 a,b | 0.403 b | 139.760 a | 108.167 a | 34.333 b | 83.753 a | 21.673 a | |
| G-NHG, mg/kg | 100 | 39.867 b | 34.267 b | 10.067 b | 0.504 a | 136.543 a | 101.263 bc | 36.680 a,b | 79.611 a | 20.253 a,b |
| 200 | 39.200 b | 33.220 b | 9.933 b | 0.383 b | 130.617 b | 104.930 bc | 37.533 a,b | 72.097 b | 20.986 a,b | |
| 300 | 42.900 b | 33.233 b | 10.367 b | 0.411 b | 123.300 c | 98.263 b | 38.067 a,b | 65.581 b | 19.653 b | |
| 400 | 43.273 b | 33.167 b | 11.600 a,b | 0.397 b | 109.450 d | 88.667 c | 40.133 a | 51.583 c | 17.733 c | |
| p value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| SEM | 1.347 | 1.076 | 0.664 | 0.580 | 1.047 | 1.767 | 1.074 | 1.841 | 0.353 | |
| Group | PC | NC | G-NHG, mg/kg | SEM | p Value | ||||
|---|---|---|---|---|---|---|---|---|---|
| Parameter | 100 | 200 | 300 | 400 | |||||
| Economic parameters ($) | |||||||||
| Feed cost (AVC) | 1.5503 d | 1.6807 c | 1.5941 d | 1.6893 c | 1.8820 b | 1.9774 a | 0.011 | <0.001 | |
| Feed cost/kg body weight | 0.8383 a | 0.7127 b | 0.8535 a | 0.8318 a | 0.8490 a | 0.8419 a | 0.007 | <0.001 | |
| Total expenses | 2.7116 e | 2.8420 c | 2.7554 d | 2.8506 c | 3.0433 b | 3.1387 a | 0.011 | <0.001 | |
| Total revenue | 3.4227d | 4.3393 a | 3.4538 d | 3.7491 c | 4.0838 b | 4.3234 a | 0.016 | <0.001 | |
| Net profit | 0.7112 e | 1.4973 a | 0.6984 e | 0.8985 d | 1.0406 c | 1.1847 b | 0.022 | <0.001 | |
| Economic efficiency | 0.4594 d | 0.8909 a | 0.4382 d | 0.5321 c | 0.5538 bc | 0.5991 b | 0.016 | <0.001 | |
| Cost/benefit ratio | 1.2625 d | 1.5269 a | 1.2535 d | 1.3153 c | 1.3422 c | 1.3774 b | 0.009 | <0.001 | |
| Profitability index | 0.2075 d | 0.3450 a | 0.2022 d | 0.2396 c | 0.2548 c | 0.2740 b | 0.005 | <0.001 | |
| Losses due to mortality | |||||||||
| Profit losses | 2.2995 a | 0.6243 b | 0.9980 b | 1.1009 b | 1.0458 b | 0.4973 b | 0.266 | <0.001 | |
| Costs of mortality | 8.6910 a | 1.1343 b | 3.8585 b | 3.4253 b | 3.0660 b | 1.2588 b | 0.807 | <0.001 | |
| Total losses | 10.990 a | 1.7586 b | 4.8564 b | 4.5263 b | 4.1118 b | 1.7561 b | 1.05 | <0.001 | |
| Mortality % | 32.00 a | 4.00 b | 14.00 b | 12.00 b | 10.00 b | 4.00 b | 2.76 | <0.001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, D.; Ismail, T.A.; Khalifa, E.; Abd El-Kader, S.A.; Mohamed, D.I.; Mohamed, D.T.; Shahin, S.E.; Abd El-Hamid, M.I. Supplementing Garlic Nanohydrogel Optimized Growth, Gastrointestinal Integrity and Economics and Ameliorated Necrotic Enteritis in Broiler Chickens Using a Clostridium perfringens Challenge Model. Animals 2021, 11, 2027. https://doi.org/10.3390/ani11072027
Ibrahim D, Ismail TA, Khalifa E, Abd El-Kader SA, Mohamed DI, Mohamed DT, Shahin SE, Abd El-Hamid MI. Supplementing Garlic Nanohydrogel Optimized Growth, Gastrointestinal Integrity and Economics and Ameliorated Necrotic Enteritis in Broiler Chickens Using a Clostridium perfringens Challenge Model. Animals. 2021; 11(7):2027. https://doi.org/10.3390/ani11072027
Chicago/Turabian StyleIbrahim, Doaa, Tamer Ahmed Ismail, Eman Khalifa, Shaimaa A. Abd El-Kader, Dalia Ibrahim Mohamed, Dalia T. Mohamed, Sara E. Shahin, and Marwa I. Abd El-Hamid. 2021. "Supplementing Garlic Nanohydrogel Optimized Growth, Gastrointestinal Integrity and Economics and Ameliorated Necrotic Enteritis in Broiler Chickens Using a Clostridium perfringens Challenge Model" Animals 11, no. 7: 2027. https://doi.org/10.3390/ani11072027
APA StyleIbrahim, D., Ismail, T. A., Khalifa, E., Abd El-Kader, S. A., Mohamed, D. I., Mohamed, D. T., Shahin, S. E., & Abd El-Hamid, M. I. (2021). Supplementing Garlic Nanohydrogel Optimized Growth, Gastrointestinal Integrity and Economics and Ameliorated Necrotic Enteritis in Broiler Chickens Using a Clostridium perfringens Challenge Model. Animals, 11(7), 2027. https://doi.org/10.3390/ani11072027