
Development of a Pig Mammary Epithelial Cell Culture Model as a Non-Clinical Tool for Studying Epithelial Barrier—A Contribution from the IMI-ConcePTION Project

 , , , ,
, , , ,  ,
,  , , , and
, , , and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Isolation and Culture of Porcine Mammary Epithelial Cells (pMECs)
2.3. Histological and Immunohistochemical Examination of Mammary Gland Tissues
2.4. Cell Cycle Analysis
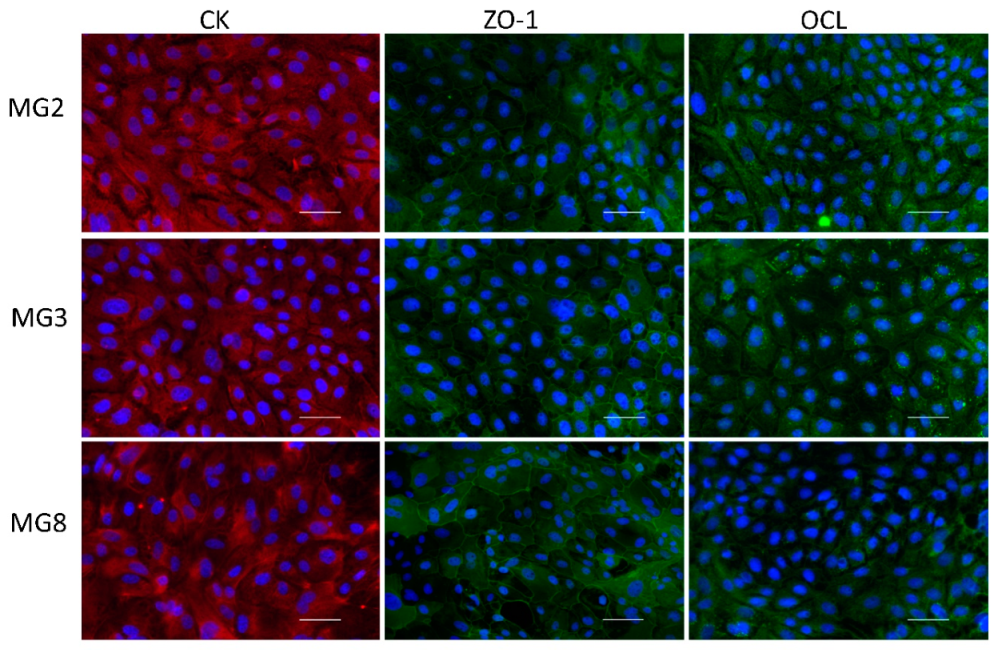
2.5. Immunofluorescence of Cytokeratins (CKs), Zonula Occludens-1 (ZO-1), Occludin (OCL)
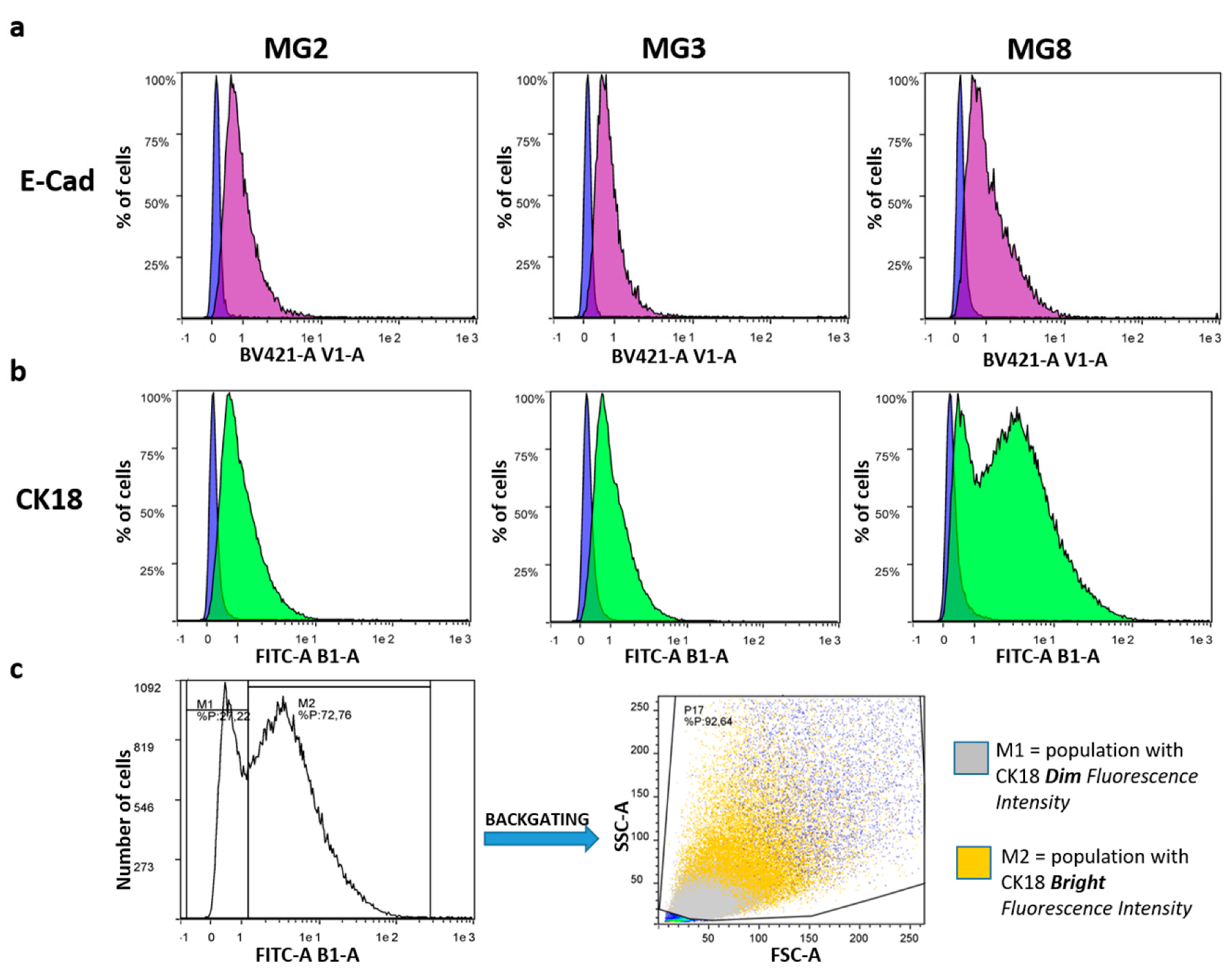
2.6. Epithelial Cadherin (E-Cad) and Cytokeratin18 (Ck18) Analysis by Flow Cytometry
2.7. Barrier Study: TEER Measure and Sodium Fluorescein Transport
2.8. pMECs RNA Extraction and Pig Drug Transporters Analysis by RT2 Profilertm PCR Array
2.9. Statistical Analysis
3. Results
3.1. Histological and Immunohistochemical Characteristics of the Mammary Glands
3.2. pMECs Morphology, Expansion and Epithelial Marker Expression
3.3. pMECs Barrier Function
3.4. pMECs Drug Transporter Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- ConcePTION Homepage. Available online: https://www.imi-conception.eu/ (accessed on 25 February 2021).
- Ventrella, D.; Forni, M.; Bacci, M.L.; Annaert, P. Non-clinical Models to Determine Drug Passage into Human Breast Milk. Curr. Pharm. Des. 2019, 25, 534–548. [Google Scholar] [CrossRef]
- ConcePTION, D3.2 Report on Lactation Characteristics of Animal Species; Selection of the Animal Species to be Used in In Vivo Studies. Available online: https://www.imi-conception.eu/wp-content/uploads/2020/04/ConcePTION-D3.2-Report-on-lactation-characteristics-of-animal-species-Selection-of-the-animal-species-to-be-used-in-in-vivo-studies.pdf (accessed on 25 February 2021).
- Ventrella, D.; Ashkenazi, N.; Elmi, A.; Allegaert, K.; Aniballi, C.; DeLise, A.; Devine, P.; Smits, A.; Steiner, L.; Forni, M.; et al. Animal Models for In Vivo Lactation Studies: Anatomy, Physiology and Milk Compositions in the Most Used Non-Clinical Species: A Contribution from the ConcePTION Project. Animals 2021, 11, 714. [Google Scholar] [CrossRef]
- McManaman, J.L.; Neville, M.C. Mammary physiology and milk secretion. Adv. Drug Deliv. Rev. 2003, 55, 629–641. [Google Scholar] [CrossRef]
- Hurley, W.L. Review: Mammary gland development in swine: Embryo to early lactation. Animal 2019, 13, s11–s19. [Google Scholar] [CrossRef] [PubMed]
- Toddywalla, V.S.; Kari, F.W.; Neville, M.C. Active transport of nitrofurantoin across a mouse mammary epithelial monolayer. J. Pharmacol. Exp. Ther. 1997, 280, 669–676. [Google Scholar]
- Kimura, S.; Morimoto, K.; Okamoto, H.; Ueda, H.; Kobayashi, D.; Kobayashi, J.; Morimoto, Y. Development of a human mammary epithelial cell culture model for evaluation of drug transfer into milk. Arch. Pharmacal Res. 2006, 29, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Marshall, A.M.; Pai, V.P.; Sartor, M.A.; Horseman, N.D. In vitro multipotent differentiation and barrier function of a human mammary epithelium. Cell Tissue Res. 2009, 335, 383–395. [Google Scholar] [CrossRef]
- Andersson, M.; Ersson, L.; Brandt, I.; Bergström, U. Potential transfer of neurotoxic amino acid β- N -methylamino-alanine (BMAA) from mother to infant during breast-feeding: Predictions from human cell lines. Toxicol. Appl. Pharmacol. 2017, 320, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Nauwelaerts, N.; Deferm, N.; Smits, A.; Bernardini, C.; Lammens, B.; Gandia, P.; Panchaud, A.; Nordeng, H.; Bacci, M.L.; Forni, M.; et al. A comprehensive review on non-clinical methods to study transfer of medication into breast milk – A contribution from the ConcePTION project. Biomed. Pharmacother. 2021, 136, 111038. [Google Scholar] [CrossRef]
- Kumura, H.; Tanaka, A.; Abo, Y.; Yui, S.; Shimazaki, K.-I.; Kobayashi, E.; Sayama, K. Primary Culture of Porcine Mammary Epithelial Cells as a Model System for Evaluation of Milk Protein Expression. Biosci. Biotechnol. Biochem. 2001, 65, 2098–2101. [Google Scholar] [CrossRef]
- Zheng, Y.-M.; He, X.-Y. Characteristics and EGFP expression of porcine mammary gland epithelial cells. Res. Vet. Sci. 2010, 89, 383–390. [Google Scholar] [CrossRef]
- Sun, Y.; Lin, C.; Chou, Y. Establishment and characterization of a spontaneously immortalized porcine mammary epithelial cell line. Cell Biol. Int. 2006, 30, 970–976. [Google Scholar] [CrossRef] [PubMed]
- Dahanayaka, S.; Rezaei, R.; Porter, W.W.; Johnson, G.A.; Burghardt, R.C.; Bazer, F.W.; Hou, Y.Q.; Wu, Z.L.; Wu, G. Technical note: Isolation and characterization of porcine mammary epithelial cells. J. Anim. Sci. 2015, 93, 5186–5193. [Google Scholar] [CrossRef]
- European Commission. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific PurposesText with EEA Relevance. 47. Off. J. Eur. Union 2010, 276, 33–79. [Google Scholar]
- Murtagh, J.; McArdle, E.; Gilligan, E.; Thornton, L.; Furlong, F.; Martin, F. Organization of mammary epithelial cells into 3D acinar structures requires glucocorticoid and JNK signaling. J. Cell Biol. 2004, 166, 133–143. [Google Scholar] [CrossRef]
- Servais, H.; Van Der Smissen, P.; Thirion, G.; Van Der Essen, G.; Van Bambeke, F.; Tulkens, P.M.; Mingeot-Leclercq, M.-P. Gentamicin-induced apoptosis in LLC-PK1 cells: Involvement of lysosomes and mitochondria. Toxicol. Appl. Pharmacol. 2005, 206, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, A.; Dong, Q.; Sun, L.-Z. Stem/Progenitor Cells in Murine Mammary Gland: Isolation and Functional Characterization. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2012; Volume 879, pp. 179–193. [Google Scholar] [CrossRef]
- Dong, Q.; Wang, D.; Bandyopadhyay, A.; Gao, H.; Gorena, K.M.; Hildreth, K.; Rebel, V.I.; Walter, C.A.; Huang, C.; Sun, L.-Z. Mammospheres from murine mammary stem cell-enriched basal cells: Clonal characteristics and repopulating potential. Stem Cell Res. 2013, 10, 396–404. [Google Scholar] [CrossRef]
- Ogorevc, J.; Zorc, M.; Dovč, P. Development of an In Vitro Goat Mammary Gland Model: Establishment, Characterization, and Applications of Primary Goat Mammary Cell Cultures. In Goat Science; IntechOpen: London, UK, 2018. [Google Scholar]
- Zaniboni, A.; Bernardini, C.; Alessandri, M.; Mangano, C.; Zannoni, A.; Bianchi, F.; Sarli, G.; Calzà, L.; Bacci, M.L.; Forni, M. Cells derived from porcine aorta tunica media show mesenchymal stromal-like cell properties in in vitro culture. Am. J. Physiol. Cell Physiol. 2014, 306, C322–C333. [Google Scholar] [CrossRef][Green Version]
- Levi, M.; Salaroli, R.; Parenti, F.; De Maria, R.; Zannoni, A.; Bernardini, C.; Gola, C.; Brocco, A.; Marangio, A.; Benazzi, C.; et al. Doxorubicin treatment modulates chemoresistance and affects the cell cycle in two canine mammary tumour cell lines. BMC Vet. Res. 2021, 17, 1–15. [Google Scholar] [CrossRef]
- Bernardini, C.; Algieri, C.; Mantia, L.D.; Zannoni, A.; Salaroli, R.; Trombetti, F.; Forni, M.; Pagliarani, A.; Nesci, S. Relationship between serum concentration, functional parameters and cell bioenergetics in IPEC-J2 cell line. Histochem. Cell Biol. 2021, 1–9. [Google Scholar] [CrossRef]
- Tubon, I.; Zannoni, A.; Bernardini, C.; Salaroli, R.; Bertocchi, M.; Mandrioli, R.; Vinueza, D.; Antognoni, F.; Forni, M. In Vitro Anti-Inflammatory Effect of Salvia sagittata Ethanolic Extract on Primary Cultures of Porcine Aortic Endothelial Cells. Oxidative Med. Cell. Longev. 2019, 2019, 6829173. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER Measurement Techniques for In Vitro Barrier Model Systems. J. Lab. Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef] [PubMed]
- Ingels, F.; Deferme, S.; Delbar, N.; Oth, M.; Augustijns, P. Implementation of the caco-2 cell culture model as a predictive tool for the oral absorption of drugs. In-house evaluation procedures. J. Pharm. Belg. 2003, 57, 153–158. [Google Scholar]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique; Methuen & Co Ltd.: London, UK, 1959. [Google Scholar]
- Sternlicht, M.D. Key stages in mammary gland development: The cues that regulate ductal branching morphogenesis. Breast Cancer Res. 2005, 8, 201. [Google Scholar] [CrossRef] [PubMed]
- Oakes, S.; Hilton, H.N.; Ormandy, C.J. Key stages in mammary gland development - The alveolar switch: Coordinating the proliferative cues and cell fate decisions that drive the formation of lobuloalveoli from ductal epithelium. Breast Cancer Res. 2006, 8, 207. [Google Scholar] [CrossRef]
- Moll, R.; Franke, W.W.; Schiller, D.L.; Geiger, B.; Krepler, R. The catalog of human cytokeratins: Patterns of expression in normal epithelia, tumors and cultured cells. Cell 1982, 31, 11–24. [Google Scholar] [CrossRef]
- Diego, C.; Eugenio, M.; Silvia, M.; Elisabetta, M.; Paolo, A.; Mario, B. Bovine mammary epithelial cells retain stem-like phenotype in long-term cultures. Res. Vet. Sci. 2014, 97, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M. Molecular Structure of Tight Junctions and Their Role in Epithelial Transport. Physiology 2001, 16, 126–130. [Google Scholar] [CrossRef]
- Stelwagen, K.; Singh, K. The Role of Tight Junctions in Mammary Gland Function. J. Mammary Gland. Biol. Neoplasia 2013, 19, 131–138. [Google Scholar] [CrossRef]
- Kari, F.W.; Weaver, R.; Neville, M.C. Active transport of nitrofurantoin across the mammary epithelium in vivo. J. Pharmacol. Exp. Ther. 1997, 280, 664–668. [Google Scholar] [PubMed]
- Ito, S.; Alcorn, J. Xenobiotic transporter expression and function in the human mammary gland. Adv. Drug Deliv. Rev. 2003, 55, 653–665. [Google Scholar] [CrossRef]
- Dean, M. ABC Transporters, Drug Resistance, and Cancer Stem Cells. J. Mammary Gland. Biol. Neoplasia 2009, 14, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Vlaming, M.L.; Lagas, J.S.; Schinkel, A.H. Physiological and pharmacological roles of ABCG2 (BCRP): Recent findings in Abcg2 knockout mice. Adv. Drug Deliv. Rev. 2009, 61, 14–25. [Google Scholar] [CrossRef] [PubMed]
- García-Lino, A.M.; Fernández, I. Álvarez; Blanco-Paniagua, E.; Merino, G.; Álvarez, A.I. Transporters in the Mammary Gland—Contribution to Presence of Nutrients and Drugs into Milk. Nutrients 2019, 11, 2372. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | P. Number | Species | Supplier | Application |
|---|---|---|---|---|
| Anti CK | GA053 | Mouse monoclonal | Agilent Dako | IF: 1:150 IHC: 1:300 |
| Anti E-Cadherin | 610181 | Mouse monoclonal | BD Transduction Laboratories | IHC: 1:2000 |
| Anti ZO-1 | 61-7300 | Rabbit | Thermo Fisher | IF: 1:100 |
| Anti OCL | H-279 | Rabbit | Santa Cruz Biotechnology | IF: 1:50 |
| Anti-Rabbit IgG-Alexa Fluor 488 | A11034 | Goat | Thermo Fisher | IF: 5 μg/mL |
| Anti-Mouse IgG-Alexa Fluor 594 | A11032 | Goat | Thermo Fisher | IF: 5 μg/mL |
| Brilliant Violet 421TM anti-E-cadherin | 147319 | Rat monoclonal | BioLegend | FC: 17 µL/106 |
| FITC anti-cytokeratin 18 | ab52459 | Mouse monoclonal | Abcam | FC: 10 µL/106 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernardini, C.; La Mantia, D.; Salaroli, R.; Zannoni, A.; Nauwelaerts, N.; Deferm, N.; Ventrella, D.; Bacci, M.L.; Sarli, G.; Bouisset-Leonard, M.; et al. Development of a Pig Mammary Epithelial Cell Culture Model as a Non-Clinical Tool for Studying Epithelial Barrier—A Contribution from the IMI-ConcePTION Project. Animals 2021, 11, 2012. https://doi.org/10.3390/ani11072012
Bernardini C, La Mantia D, Salaroli R, Zannoni A, Nauwelaerts N, Deferm N, Ventrella D, Bacci ML, Sarli G, Bouisset-Leonard M, et al. Development of a Pig Mammary Epithelial Cell Culture Model as a Non-Clinical Tool for Studying Epithelial Barrier—A Contribution from the IMI-ConcePTION Project. Animals. 2021; 11(7):2012. https://doi.org/10.3390/ani11072012
Chicago/Turabian StyleBernardini, Chiara, Debora La Mantia, Roberta Salaroli, Augusta Zannoni, Nina Nauwelaerts, Neel Deferm, Domenico Ventrella, Maria Laura Bacci, Giuseppe Sarli, Michele Bouisset-Leonard, and et al. 2021. "Development of a Pig Mammary Epithelial Cell Culture Model as a Non-Clinical Tool for Studying Epithelial Barrier—A Contribution from the IMI-ConcePTION Project" Animals 11, no. 7: 2012. https://doi.org/10.3390/ani11072012
APA StyleBernardini, C., La Mantia, D., Salaroli, R., Zannoni, A., Nauwelaerts, N., Deferm, N., Ventrella, D., Bacci, M. L., Sarli, G., Bouisset-Leonard, M., Annaert, P., & Forni, M. (2021). Development of a Pig Mammary Epithelial Cell Culture Model as a Non-Clinical Tool for Studying Epithelial Barrier—A Contribution from the IMI-ConcePTION Project. Animals, 11(7), 2012. https://doi.org/10.3390/ani11072012