
Post-Thaw Sperm Quality and Functionality in the Autochthonous Pig Breed Gochu Asturcelta

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Reagents
2.2. Genetic Resource Bank Location and Climate
2.3. Boars, Semen Collection and Cryopreservation Procedure
2.4. Assessment of Post-Thaw Sperm Parameters
2.4.1. Sperm Motility
2.4.2. Viability and Acrosomal Status
2.4.3. Mitochondrial Membrane Potential
2.4.4. Reactive Oxygen Species (ROS)
2.4.5. Apoptosis
2.4.6. Sperm Chromatin Status (Sperm Chromatin Structure Assay, SCSA)
2.5. Statistical Analysis
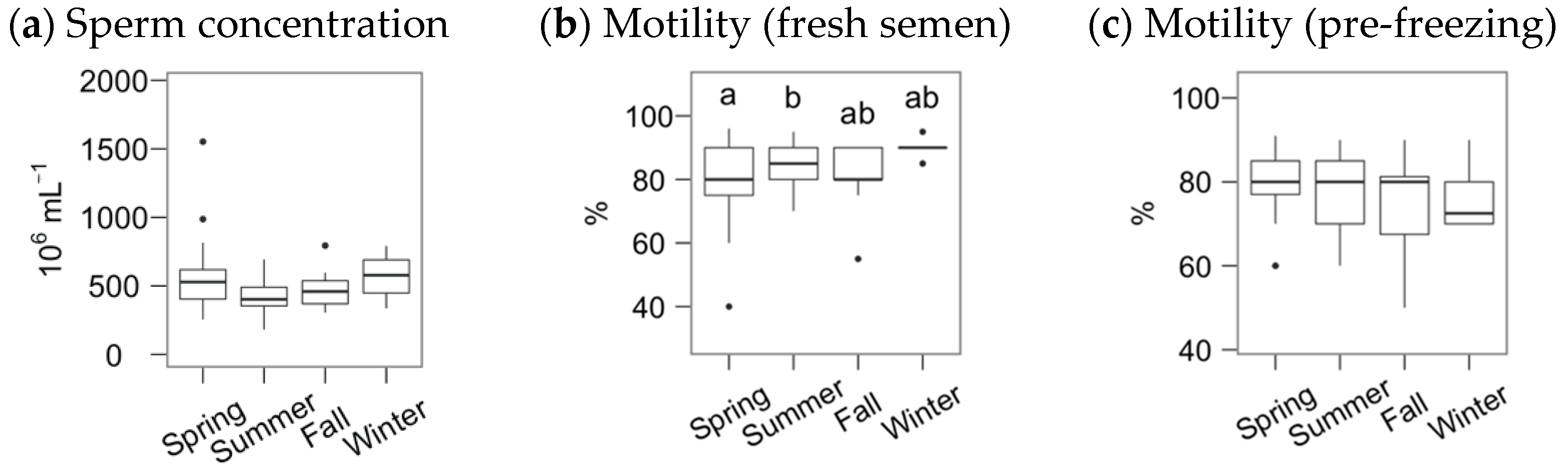
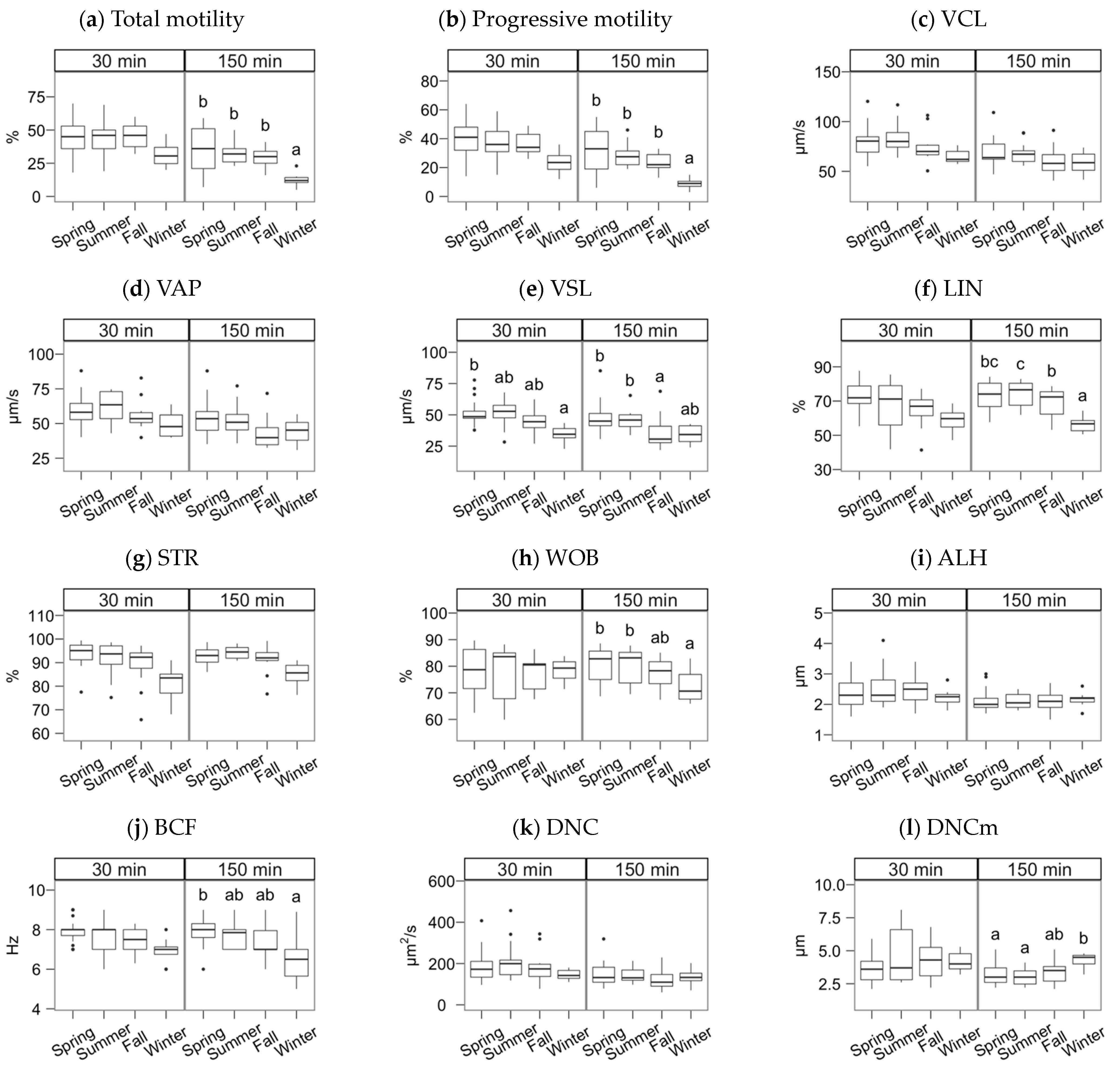
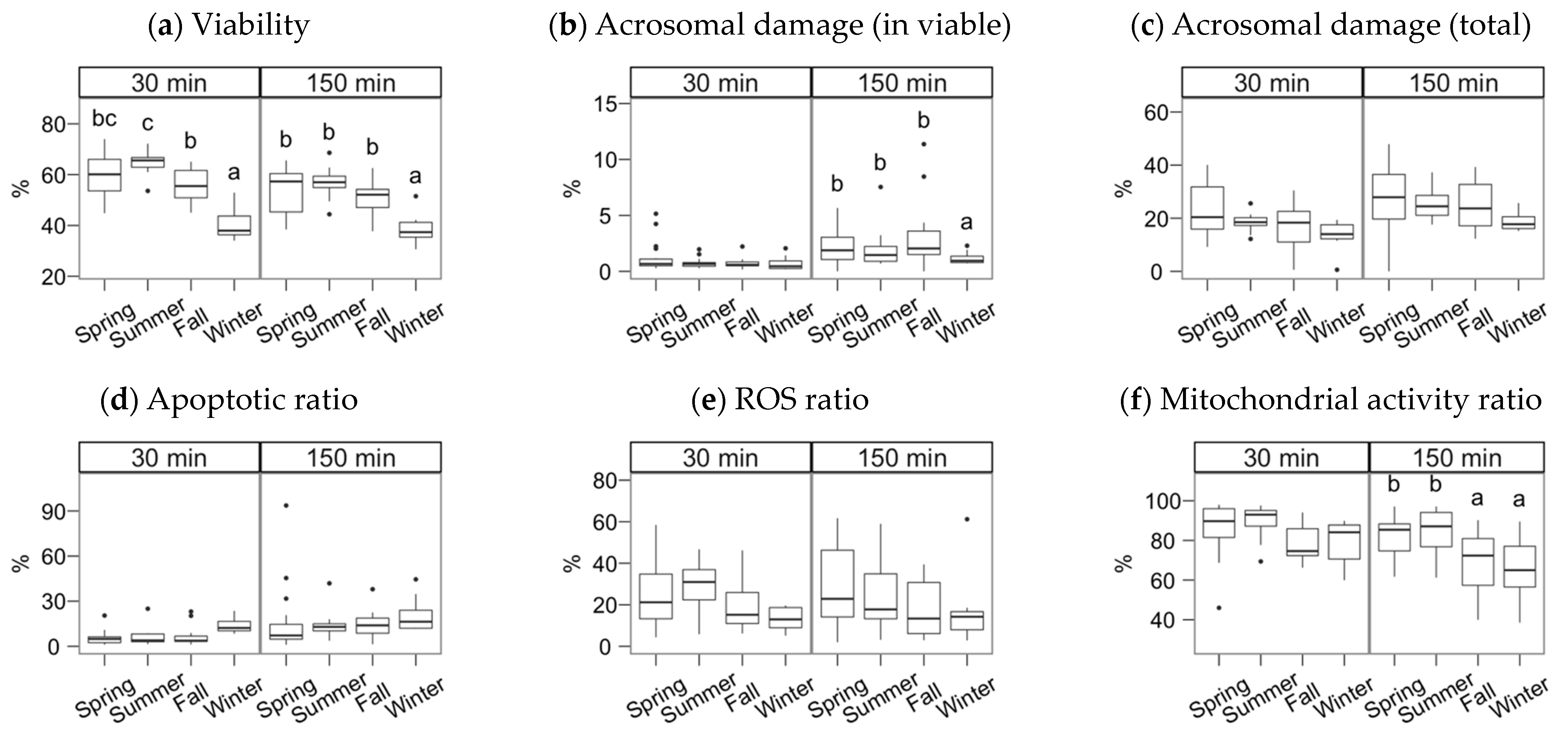
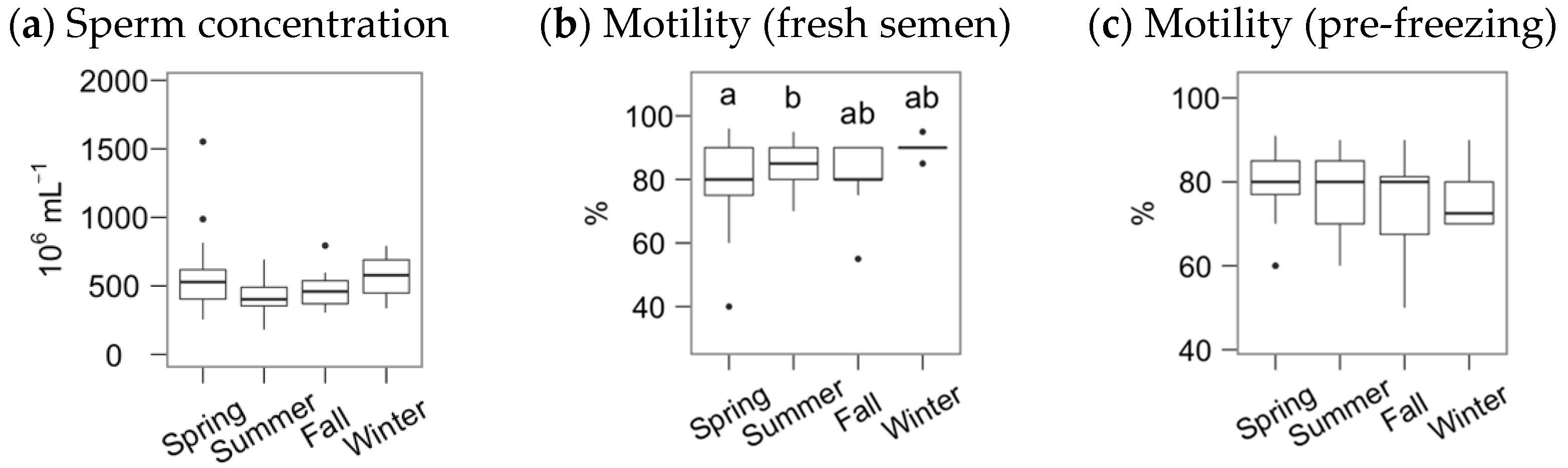
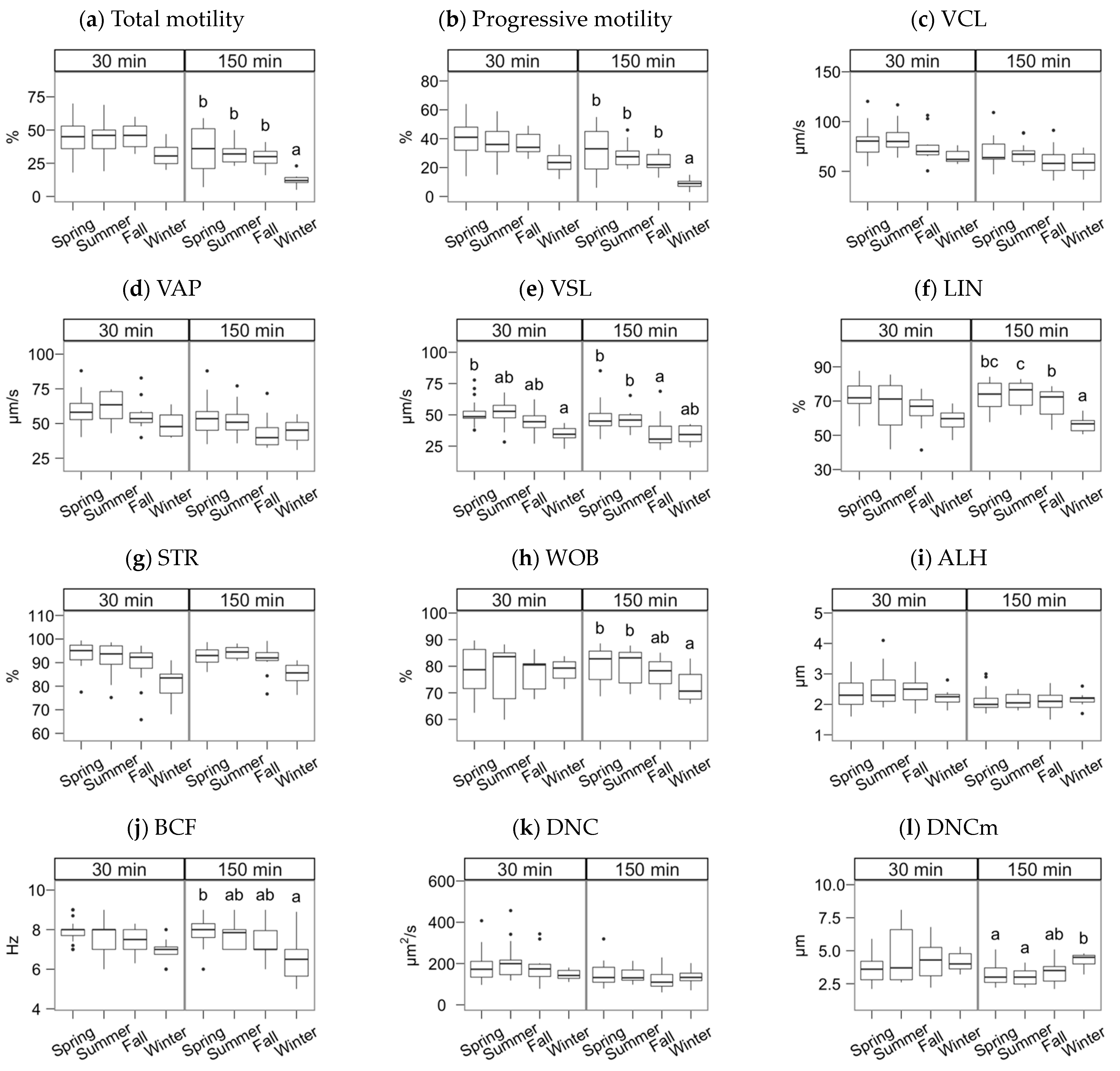
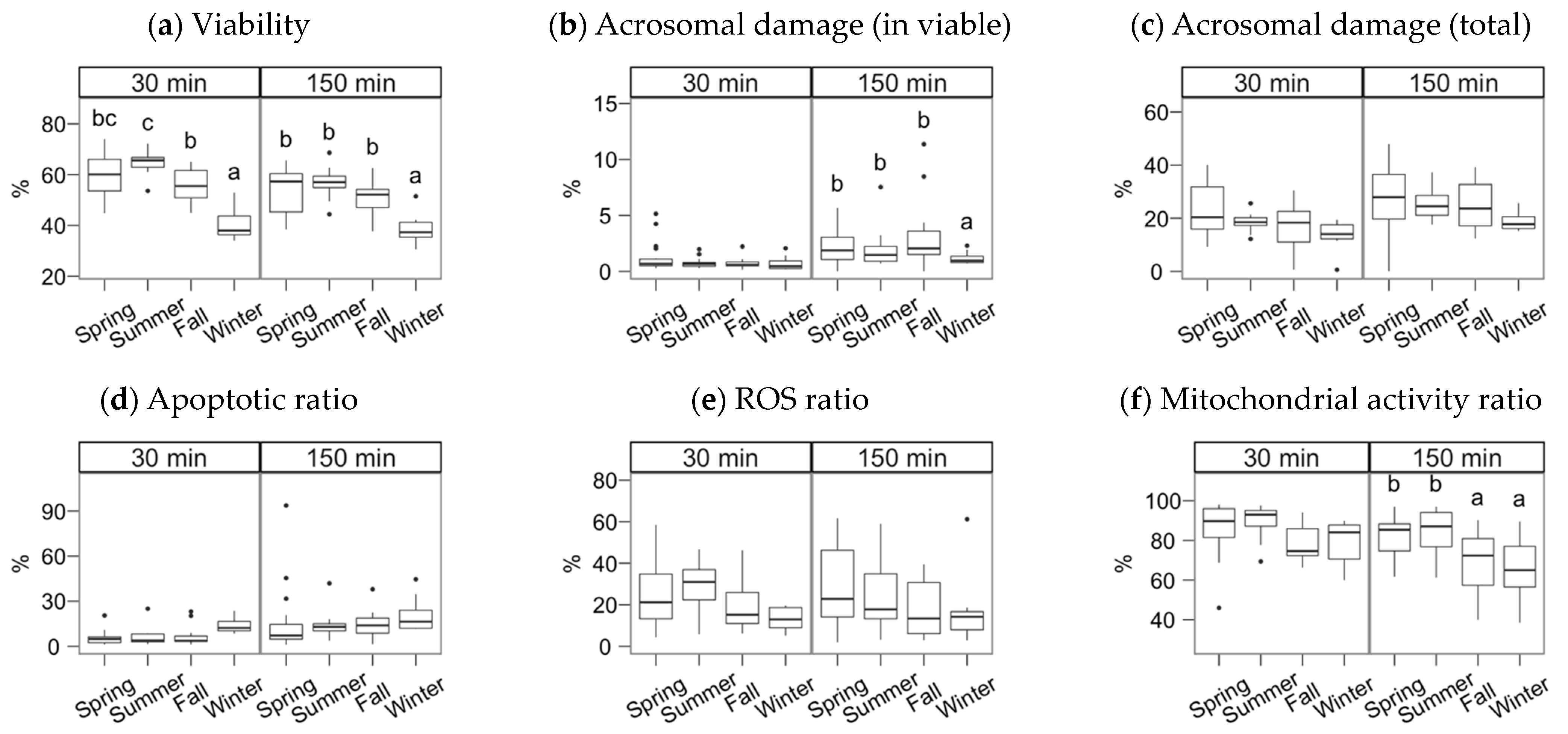
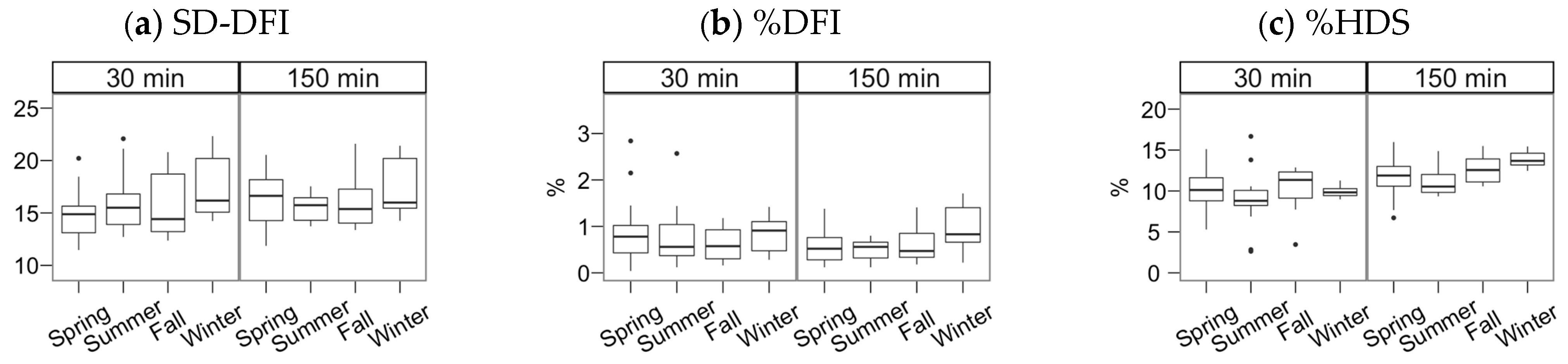
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andrabi, S.; Maxwell, W. A review on reproductive biotechnologies for conservation of endangered mammalian species. Anim. Reprod. Sci. 2007, 99, 223–243. [Google Scholar] [CrossRef]
- FAO. The Second Report on the State of the World’s Animal Genetic Resources for Food and Agriculture; FAO commission on genetic resources for food and agriculture, Assessments 2015; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018. [Google Scholar]
- Henson, E.L. In Situ Conservation of Livestock and Poultry; FAO Animal Production & Health Paper 99; Food and Agriculture Organization of the United Nations: Rome, Italy, 1992; 112p. [Google Scholar]
- Evans, N.; Yarwood, R. The Politicization of Livestock: Rare Breeds and Countryside Conservation. Sociol. Rural. 2000, 40, 228–248. [Google Scholar] [CrossRef]
- Solti, L.; Crichton, E.; Loskutoff, N.; Cseh, S. Economical and ecological importance of indigenous livestock and the application of assisted reroduction to their preservation. Theriogenology 2000, 53, 149–162. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Influence of Climate on Livestock Breeding. Agrobiodiversity: The Case for Conserving Domestic and Related Animals; FAO Fact sheet on the conservation of domestic animal genetic resources; Food and Agriculture Organization of the United Nations: Rome, Italy, 1993. [Google Scholar]
- Marsoner, T.; Vigl, L.E.; Manck, F.; Jaritz, G.; Tappeiner, U.; Tasser, E. Indigenous livestock breeds as indicators for cultural ecosystem services: A spatial analysis within the Alpine Space. Ecol. Indic. 2018, 94, 55–63. [Google Scholar] [CrossRef]
- Mendelsohn, R. The challenge of conserving indigenous domesticated animals. Ecol. Econ. 2003, 45, 501–510. [Google Scholar] [CrossRef]
- Woelders, H.; Windig, J.; Hiemstra, S. How Developments in Cryobiology, Reproductive Technologies and Conservation Genomics Could Shape Gene Banking Strategies for (Farm) Animals. Reprod. Domest. Anim. 2012, 47, 264–273. [Google Scholar] [CrossRef]
- Argamentería-Gutiérrez, A.; de la Roza-Delgado, B.; Cueto-Ardavín, M.; Hidalgo-Ordóñez, C.O.; Tamargo-Miguel, C.; Menéndez-Fernández, J. Guía del Gochu Asturcelta; Servicio Regional de investigación y Desarrollo Agroalimentario: Villaviciosa, Spain, 2012; ISBN 978-84-695-3048-1. [Google Scholar]
- Tamargo, C.; Rodríguez, A.; Fernández, A.; Merino, M.; Reyes, M.; Benito, J.; Hidalgo, C. Programa de conservación del Gochu Astur-Celta: Creación de un banco de germoplasma. Arch. Zootec. 2011, 60, 365–368. [Google Scholar] [CrossRef] [Green Version]
- Holt, W.V. Role of reproductive technologies and genetic resource banks in animal conservation. Rev. Reprod. 1999, 4, 143–150. [Google Scholar] [CrossRef]
- Leroy, G.; Boettcher, P.; Besbes, B.; Danchin-Burge, C.; Baumung, R.; Hiemstra, S.J. Cryoconservation of Animal Genetic Resources in Europe and Two African Countries: A Gap Analysis. Diversity 2019, 11, 240. [Google Scholar] [CrossRef] [Green Version]
- Guthrie, H.; Welch, G. Impact of storage prior to cryopreservation on plasma membrane function and fertility of boar sperm. Theriogenology 2005, 63, 396–410. [Google Scholar] [CrossRef]
- Roca, J.; Hernández, M.; Carvajal, G.; Vazquez, J.M.; Martínez, E.A. Factors influencing boar sperm cryosurvival1. J. Anim. Sci. 2006, 84, 2692–2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrilla, I.; Del Olmo, D.; Sijses, L.; Martinez-Alborcia, M.J.; Cuello, C.; Vazquez, J.M.; Martinez, E.A.; Roca, J. Differences in the ability of spermatozoa from individual boar ejaculates to withstand different semen-processing techniques. Anim. Reprod. Sci. 2012, 132, 66–73. [Google Scholar] [CrossRef]
- Yeste, M. Recent Advances in Boar Sperm Cryopreservation: State of the Art and Current Perspectives. Reprod. Domest. Anim. 2015, 50, 71–79. [Google Scholar] [CrossRef]
- Yeste, M. Sperm cryopreservation update: Cryodamage, markers, and factors affecting the sperm freezability in pigs. Theriogenology 2016, 85, 47–64. [Google Scholar] [CrossRef] [PubMed]
- Yeste, M.; Bonet, S.; Rodríguez-Gil, J.E. Artificial insemination with frozen-thawed boar sperm. Mol. Reprod. Dev. 2017, 84, 802–813. [Google Scholar] [CrossRef] [Green Version]
- Holt, W.V.; Medrano, A. Assessment of boar sperm function in relation to freezing and storage. Biosci. Proc. 2020, 52, 213–222. [Google Scholar] [CrossRef]
- Fraser, L.; Strzeżek, J. Effect of different procedures of ejaculate collection, extenders and packages on DNA integrity of boar spermatozoa following freezing–thawing. Anim. Reprod. Sci. 2007, 99, 317–329. [Google Scholar] [CrossRef]
- Knox, R. The Fertility of Frozen Boar Sperm When used for Artificial Insemination. Reprod. Domest. Anim. 2015, 50, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Tamargo, C.; Hidalgo, C.O.; Caamaño, J.N.; Salman, A.; Fueyo, C.; Arija, C.; Fernández, Á.; Merino, M.J.; Martínez-Pastor, F. Assessment of a germplasm bank for the autochthonous cattle breed Asturiana de la Montaña: Extender (Biociphos vs. BIOXCell) affected sperm quality but not field fertility. Reprod. Domest. Anim. 2019, 54, 90–93. [Google Scholar] [CrossRef]
- Pearodwong, P.; Tretipskul, C.; Panyathong, R.; Sang-Gassanee, K.; Collell, M.; Muns, R.; Tummaruk, P. Reproductive performance of weaned sows after single fixed-time artificial insemination under a tropical climate: Influences of season and insemination technique. Theriogenology 2020, 142, 54–61. [Google Scholar] [CrossRef]
- Barranco, I.; Ortega, M.D.; Martinez-Alborcia, M.J.; Vazquez, J.M.; Martinez, E.A.; Roca, J. Season of ejaculate collection influences the freezability of boar spermatozoa. Cryobiology 2013, 67, 299–304. [Google Scholar] [CrossRef] [PubMed]
- De Cáceres, M.; Martin-StPaul, N.; Turco, M.; Cabon, A.; Granda, V. Estimating daily meteorological data and downscaling climate models over landscapes. Environ. Model. Softw. 2018, 108, 186–196. [Google Scholar] [CrossRef]
- Hijmans, R.J. Geosphere: Spherical Trigonometry. R Package Version 1.5-10. 2019. Available online: https://CRAN.R-project.org/package=geosphere (accessed on 5 June 2021).
- Carvajal, G.; Cuello, C.; Ruiz, M.; Vazquez, J.M.; Martínez, E.A.; Roca, J. Effects of Centrifugation Before Freezing on Boar Sperm Cryosurvival. J. Androl. 2004, 25, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Pursel, V.G.; Johnson, L.A. Freezing of Boar Spermatozoa: Fertilizing Capacity with Concentrated Semen and a New Thawing Procedure. J. Anim. Sci. 1975, 40, 99–102. [Google Scholar] [CrossRef]
- Cremades, T.; Vazquez, J.M.; Martínez, E.A.; Roca, J.; Rodriguez-Martinez, H.; Abáigar, T. Kinematic Changes During the Cryopreservation of Boar Spermatozoa. J. Androl. 2005, 26, 610–618. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Parrilla, I.; Ortega, M.D.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J. Post-thaw boar sperm motility is affected by prolonged storage of sperm in liquid nitrogen. A retrospective study. Cryobiology 2018, 80, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Barranco, I.; Tvarijonaviciute, A.; Molina, M.F.; Martinez, E.A.; Rodriguez-Martinez, H.; Parrilla, I.; Roca, J. Seminal plasma antioxidants are directly involved in boar sperm cryotolerance. Theriogenology 2018, 107, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Perez-Patiño, C.; Barranco, I.; Li, J.; Padilla, L.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J.; Parrilla, I. Cryopreservation Differentially Alters the Proteome of Epididymal and Ejaculated Pig Spermatozoa. Int. J. Mol. Sci. 2019, 20, 1791. [Google Scholar] [CrossRef] [Green Version]
- Guthrie, H.D.; Welch, G.R. Determination of intracellular reactive oxygen species and high mitochondrial membrane potential in Percoll-treated viable boar sperm using fluorescence-activated flow cytometry1. J. Anim. Sci. 2006, 84, 2089–2100. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Roca, J.; Pérez-Patiño, C.; Barranco, I.; Martinez, E.A.; Rodriguez-Martinez, H.; Parrilla, I. Is boar sperm freezability more intrinsically linked to spermatozoa than to the surrounding seminal plasma? Anim. Reprod. Sci. 2018, 195, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Gago, R.; Álvarez-Rodríguez, M.; Alonso, M.E.; González, J.R.; Alegre, B.; Domínguez, J.C.; Martínez-Pastor, F. Thawing boar semen in the presence of seminal plasma improves motility, modifies subpopulation patterns and reduces chromatin alterations. Reprod. Fertil. Dev. 2017, 29, 1576–1584. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.r-project.org (accessed on 5 June 2021).
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyal, S.M.; Kennedy, B.W.; Wilkins, J.N. Boar, breed and environmental effects on motility of frozen-thawed spermatozoa. Can. J. Anim. Sci. 1986, 66, 663–668. [Google Scholar] [CrossRef]
- Martín-Hidalgo, D.; Barón, F.; Robina, A.; Bragado, M.; de Llera, A.H.; García-Marín, L.; Gil, M. Inter- and intra-breed comparative study of sperm motility and viability in Iberian and Duroc boar semen during long-term storage in MR-A and XCell extenders. Anim. Reprod. Sci. 2013, 139, 109–114. [Google Scholar] [CrossRef]
- Žaja, I.Ž.; Samardžija, M.; Vince, S.; Majić-Balić, I.; Vilić, M.; Đuričić, D.; Milinković-Tur, S. Influence of boar breeds or hybrid genetic composition on semen quality and seminal plasma biochemical variables. Anim. Reprod. Sci. 2016, 164, 169–176. [Google Scholar] [CrossRef]
- Žaja, I.Ž.; Samardžija, M.; Vince, S.; Vilić, M.; Majić-Balić, I.; Đuričić, D.; Milinković-Tur, S. Differences in seminal plasma and spermatozoa antioxidative systems and seminal plasma lipid and protein levels among boar breeds and hybrid genetic traits. Anim. Reprod. Sci. 2016, 170, 75–82. [Google Scholar] [CrossRef]
- Chanapiwat, P.; Kaeoket, K. Breed of Boar Influences the Optimal Concentration of Gamma-oryzanol Needed for Semen Cryopreservation. Reprod. Domest. Anim. 2015, 50, 221–226. [Google Scholar] [CrossRef]
- Bailey, J.L.; Lessard, C.; Jacques, J.; Brèque, C.; Dobrinski, I.; Zeng, W.; Galantino-Homer, H.L. Cryopreservation of boar semen and its future importance to the industry. Theriogenology 2008, 70, 1251–1259. [Google Scholar] [CrossRef]
- Waterhouse, K.E.; Hofmo, P.O.; Tverdal, A.; Miller, R.R. Within and between breed differences in freezing tolerance and plasma membrane fatty acid composition of boar sperm. Reproduction 2006, 131, 887–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pintado, B.; de la Fuente, J.; Roldan, E.R. Permeability of boar and bull spermatozoa to the nucleic acid stains propidium iodide or Hoechst 33258, or to eosin: Accuracy in the assessment of cell viability. J. Reprod. Fertil. 2000, 118, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Martinez, H.; Wallgren, M. Advances in Boar Semen Cryopreservation. Vet. Med. Int. 2011, 2011, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, M.; Roca, J.; Gil, M.A.; Vazquez, J.M.; Martínez, E.A. Adjustments on the cryopreservation conditions reduce the incidence of boar ejaculates with poor sperm freezability. Theriogenology 2007, 67, 1436–1445. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.J.; Lee, S.H.; Park, C.S. Comparison of Semen Characteristics, Frozen-Thawed Sperm Viability, Testosterone Concentration and Embryo Development between Yorkshire Boar A and B. Asian-Australas. J. Anim. Sci. 2004, 17, 612–616. [Google Scholar] [CrossRef]
- Trzcinska, M.; Bryła, M.; Smorag, Z. Apoptotic-like changes in the spermatozoa of fresh and stored boar semen and the quality of embryos produced in vivo. Anim. Reprod. Sci. 2011, 124, 90–97. [Google Scholar] [CrossRef]
- Pezo, F.; Zambrano, F.; Uribe, P.; Moya, C.; de Andrade, A.F.C.; Risopatron, J.; Yeste, M.; Burgos, R.A.; Sánchez, R. Oxidative and nitrosative stress in frozen-thawed pig spermatozoa. I: Protective effect of melatonin and butylhydroxytoluene on sperm function. Res. Vet. Sci. 2021, 136, 143–150. [Google Scholar] [CrossRef]
- Moraes, C.R.; Meyers, S. The sperm mitochondrion: Organelle of many functions. Anim. Reprod. Sci. 2018, 194, 71–80. [Google Scholar] [CrossRef]
- Peña, F.J. Assessment of fresh and frozen–thawed boar semen using an Annexin-V assay: A new method of evaluating sperm membrane integrity. Theriogenology 2003, 60, 677–689. [Google Scholar] [CrossRef]
- Evenson, D.P.; Larson, K.L.; Jost, L.K. Sperm Chromatin Structure Assay: Its Clinical Use for Detecting Sperm DNA Fragmentation in Male Infertility and Comparisons With Other Techniques. J. Androl. 2002, 23, 25–43. [Google Scholar] [CrossRef]
- Majd, N.E.; Sadeghi, N.; Tavalaee, M.; Tabandeh, M.R.; Nasr-Esfahani, M.-H. Evaluation of Oxidative Stress in Testis and Sperm of Rat Following Induced Varicocele. Urol. J. 2019, 16, 300–306. [Google Scholar]
- Hernández, M.; Roca, J.; Ballester, J.; Vazquez, J.M.; Martínez, E.A.; Johannisson, A.; Saravia, F.; Rodriguez-Martinez, H. Differences in SCSA outcome among boars with different sperm freezability. Int. J. Androl. 2006, 29, 583–591. [Google Scholar] [CrossRef]
- Didion, B.A.; Kasperson, K.M.; Wixon, R.L.; Evenson, D.P. Boar Fertility and Sperm Chromatin Structure Status: A Retrospective Report. J. Androl. 2009, 30, 655–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evenson, D.P. Loss of livestock breeding efficiency due to uncompensable sperm nuclear defects. Reprod. Fertil. Dev. 1999, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Evenson, D.P. The Sperm Chromatin Structure Assay (SCSA®) and other sperm DNA fragmentation tests for evaluation of sperm nuclear DNA integrity as related to fertility. Anim. Reprod. Sci. 2016, 169, 56–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evenson, D.; Thompson, L.; Jost, L. Flow cytometric evaluation of boar semen by the sperm chromatin structure assay as related to cryopreservation and fertility. Theriogenology 1994, 41, 637–651. [Google Scholar] [CrossRef]
- Blackburn, H.D. Genetic Selection and Conservation of Genetic Diversity*. Reprod. Domest. Anim. 2012, 47, 249–254. [Google Scholar] [CrossRef]
- Knecht, D.; Jankowska-Mąkosa, A.; Duziński, K. The effect of age, interval collection and season on selected semen parameters and prediction of AI boars productivity. Livest. Sci. 2017, 201, 13–21. [Google Scholar] [CrossRef]
- Czubaszek, M.; Andraszek, K.; Banaszewska, D. Influence of the age of the individual on the stability of boar sperm genetic material. Theriogenology 2020, 147, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Kuhlgatz, D.; Kuhlgatz, C.; Aepli, M.; Schumann, B.; Grossfeld, R.; Bortfeldt, R.; Jakop, U.; Jung, M.; Schulze, M. Development of predictive models for boar semen quality. Theriogenology 2019, 134, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Trudeau, V.; Sanford, L.M. Effect of Season and Social Environment on Testis Size and Semen Quality of the Adult Landrace Boar2. J. Anim. Sci. 1986, 63, 1211–1219. [Google Scholar] [CrossRef]
- Murase, T.; Imaeda, N.; Yamada, H.; Miyazawa, K. Seasonal Changes in Semen Characteristics, Composition of Seminal Plasma and Frequency of Acrosome Reaction Induced by Calcium and Calcium Ionophore A23187 in Large White Boars. J. Reprod. Dev. 2007, 53, 853–865. [Google Scholar] [CrossRef] [Green Version]
- Ciereszko, A.; Ottobre, J.; Glogowski, J. Effects of season and breed on sperm acrosin activity and semen quality of boars. Anim. Reprod. Sci. 2000, 64, 89–96. [Google Scholar] [CrossRef]
- Kunavongkrit, A.; Suriyasomboon, A.; Lundeheim, N.; Heard, T.W.; Einarsson, S. Management and sperm production of boars under differing environmental conditions. Theriogenology 2005, 63, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, B.; Wang, X.; Liu, X.; Zhang, Q.; Chen, Y. Estimation of genetic parameters and season effects for semen traits in three pig breeds of South China. J. Anim. Breed. Genet. 2019, 136, 183–189. [Google Scholar] [CrossRef]
- Sancho, S.; Pinart, E.; Briz, M.; Garcia-Gil, N.; Badia, E.; Bassols, J.; Kádár, E.; Pruneda, A.; Bussalleu, E.; Yeste, M.; et al. Semen quality of postpubertal boars during increasing and decreasing natural photoperiods. Theriogenology 2004, 62, 1271–1282. [Google Scholar] [CrossRef] [PubMed]
- Petrocelli, H.; Batista, C.; Gosalvez, J. Seasonal variation in sperm characteristics of boars in southern Uruguay. Rev. Bras. Zootec. 2015, 44, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Rivera, M.M.; Moreno, A.A.Q.; Barrera, X.; Palomo, M.J.; Rigau, T.; Rodríguez-Gil, J.E. Natural Mediterranean photoperiod does not affect the main parameters of boar-semen quality analysis. Theriogenology 2005, 64, 934–946. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Yi, Y. Comparison of semen characteristics, sperm freezability and testosterone concentration between Duroc and Yorkshire boars during seasons. Anim. Reprod. Sci. 2002, 73, 53–61. [Google Scholar] [CrossRef]
- Argenti, L.E.; Parmeggiani, B.; Leipnitz, G.; Weber, A.; Pereira, G.R.; Bustamante-Filho, I.C. Effects of season on boar semen parameters and antioxidant enzymes in the south subtropical region in Brazil. Andrologia 2018, 50, e12951. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, H.; Welch, G.; Long, J. Mitochondrial function and reactive oxygen species action in relation to boar motility. Theriogenology 2008, 70, 1209–1215. [Google Scholar] [CrossRef]
- Fernández-Gago, R.; Domínguez, J.C.; Martínez-Pastor, F. Seminal plasma applied post-thawing affects boar sperm physiology: A flow cytometry study. Theriogenology 2013, 80, 400–410. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Lower Hinge | Q1 | Median | Q3 | Higher Hinge |
|---|---|---|---|---|---|
| Total motility (%) | 18.0 | 34.0 | 44.0 | 51.0 | 70.0 |
| Progressive motility (%) | 12.0 | 28.0 | 36.0 | 45.0 | 64.0 |
| VCL (µm/s) | 50.5 | 66.8 | 74.3 | 84.7 | 106.3 |
| VSL (µm/s) | 22.8 | 40.2 | 47.9 | 52.8 | 71.0 |
| VAP (µm/s) | 39.8 | 51.4 | 56.6 | 64.5 | 82.8 |
| LIN (%) | 41.4 | 59.4 | 68.7 | 76.3 | 87.7 |
| STR (%) | 74.6 | 86.7 | 92.8 | 95.5 | 99.4 |
| WOB (%) | 59.9 | 71.4 | 80.5 | 84.5 | 89.7 |
| ALH (µm) | 1.6 | 2.1 | 2.3 | 2.7 | 3.5 |
| BCF (Hz) | 6.0 | 7.0 | 8.0 | 8.0 | 9.0 |
| DNC (µm2/s) | 77.8 | 137.6 | 169.8 | 205.8 | 304.3 |
| DNCm (µm) | 2.1 | 2.9 | 3.7 | 5.0 | 8.1 |
| Parameter | Lower Hinge | Q1 | Median | Q3 | Higher Hinge |
|---|---|---|---|---|---|
| Viability (%) | 34.0 | 51.3 | 60.0 | 65.5 | 74.0 |
| Damaged acrosomes (ratio viable) (%) | 0.2 | 0.4 | 0.6 | 1.0 | 1.5 |
| Damaged acrosomes (%) | 9.2 | 13.5 | 18.4 | 21.4 | 32.3 |
| Apoptotic sperm (ratio) (%) | 1.1 | 2.7 | 4.9 | 8.7 | 17.0 |
| Cytoplasmic ROS (ratio viable) (%) | 4.3 | 11.4 | 19.6 | 34.0 | 58.5 |
| Active mitochondria (ratio viable) (%) | 59.9 | 75.1 | 87.0 | 93.9 | 98.0 |
| SD-DFI | 11.5 | 13.5 | 15.1 | 17.0 | 22.1 |
| %DFI (%) | 0.0 | 0.4 | 0.7 | 1.0 | 1.4 |
| %HDS (%) | 5.3 | 8.6 | 10.0 | 11.4 | 15.1 |
| Parameter | Lower Hinge | Q1 | Median | Q3 | Higher Hinge |
|---|---|---|---|---|---|
| Total motility (%) | 5.0 | 19.5 | 28.5 | 39.5 | 59.0 |
| Progressive motility (%) | 3.0 | 15.0 | 23.0 | 33.0 | 55.0 |
| VCL (µm/s) | 40.7 | 56.5 | 63.8 | 70.5 | 91.2 |
| VSL (µm/s) | 21.9 | 32.1 | 41.9 | 49.2 | 68.8 |
| VAP (µm/s) | 30.9 | 40.0 | 49.5 | 56.0 | 77.1 |
| LIN (%) | 50.6 | 61.4 | 71.7 | 78.7 | 84.2 |
| STR (%) | 81.9 | 89.4 | 92.0 | 95.4 | 99.2 |
| WOB (%) | 65.9 | 72.8 | 79.6 | 83.8 | 88.6 |
| ALH (µm) | 1.5 | 1.9 | 2.1 | 2.3 | 2.7 |
| BCF (Hz) | 5.8 | 7.0 | 7.8 | 8.0 | 9.0 |
| DNC (µm2/s) | 60.3 | 108.8 | 130.8 | 170.1 | 229.6 |
| DNCm (µm) | 2.1 | 2.7 | 3.3 | 3.9 | 5.1 |
| Parameter | Lower Hinge | Q1 | Median | Q3 | Higher Hinge |
|---|---|---|---|---|---|
| Viability (%) | 30.6 | 44.4 | 53.3 | 59.2 | 68.6 |
| Damaged acrosomes (ratio viable) (%) | 0.0 | 1.0 | 1.7 | 2.6 | 4.4 |
| Damaged acrosomes (%) | 0.0 | 17.9 | 24.0 | 31.8 | 47.9 |
| Apoptotic sperm (ratio) (%) | 1.1 | 6.6 | 12.1 | 17.0 | 31.7 |
| Cytoplasmic ROS (ratio viable) (%) | 2.0 | 11.9 | 16.8 | 33.8 | 61.7 |
| Active mitochondria (ratio viable) (%) | 38.5 | 67.1 | 81.0 | 88.3 | 97.1 |
| SD-DFI | 11.9 | 14.3 | 15.9 | 17.6 | 21.6 |
| %DFI (%) | 0.1 | 0.3 | 0.6 | 0.8 | 1.4 |
| %HDS (%) | 6.7 | 10.6 | 12.0 | 13.6 | 16.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caamaño, J.N.; Tamargo, C.; Parrilla, I.; Martínez-Pastor, F.; Padilla, L.; Salman, A.; Fueyo, C.; Fernández, Á.; Merino, M.J.; Iglesias, T.; et al. Post-Thaw Sperm Quality and Functionality in the Autochthonous Pig Breed Gochu Asturcelta. Animals 2021, 11, 1885. https://doi.org/10.3390/ani11071885
Caamaño JN, Tamargo C, Parrilla I, Martínez-Pastor F, Padilla L, Salman A, Fueyo C, Fernández Á, Merino MJ, Iglesias T, et al. Post-Thaw Sperm Quality and Functionality in the Autochthonous Pig Breed Gochu Asturcelta. Animals. 2021; 11(7):1885. https://doi.org/10.3390/ani11071885
Chicago/Turabian StyleCaamaño, José Néstor, Carolina Tamargo, Inmaculada Parrilla, Felipe Martínez-Pastor, Lorena Padilla, Amer Salman, Carmen Fueyo, Ángel Fernández, María José Merino, Tania Iglesias, and et al. 2021. "Post-Thaw Sperm Quality and Functionality in the Autochthonous Pig Breed Gochu Asturcelta" Animals 11, no. 7: 1885. https://doi.org/10.3390/ani11071885
APA StyleCaamaño, J. N., Tamargo, C., Parrilla, I., Martínez-Pastor, F., Padilla, L., Salman, A., Fueyo, C., Fernández, Á., Merino, M. J., Iglesias, T., & Hidalgo, C. O. (2021). Post-Thaw Sperm Quality and Functionality in the Autochthonous Pig Breed Gochu Asturcelta. Animals, 11(7), 1885. https://doi.org/10.3390/ani11071885