
Effects of Orexin B on Swine Granulosa and Endothelial Cells

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Ovaries
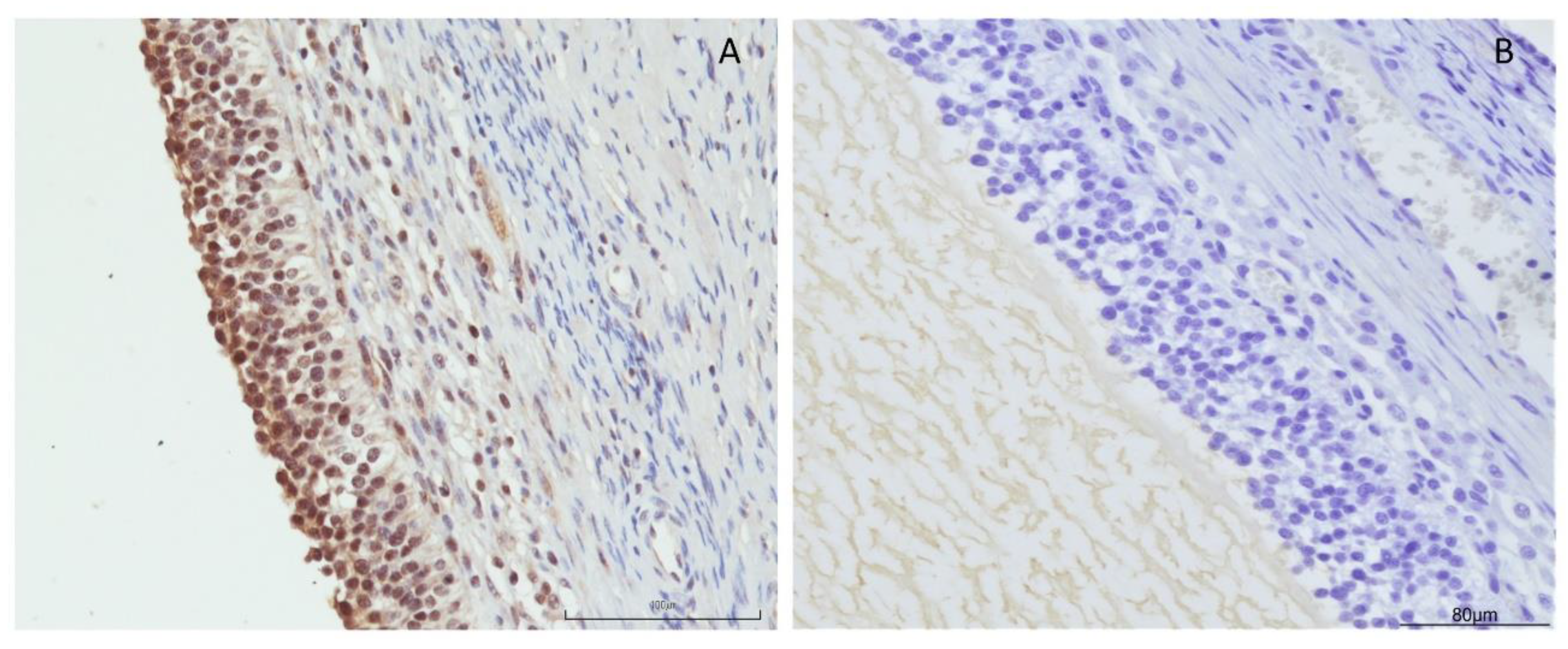
2.2. Immunolocalization of OXB
2.3. Evaluation of OXB Effect on Swine Granulosa Cell Function
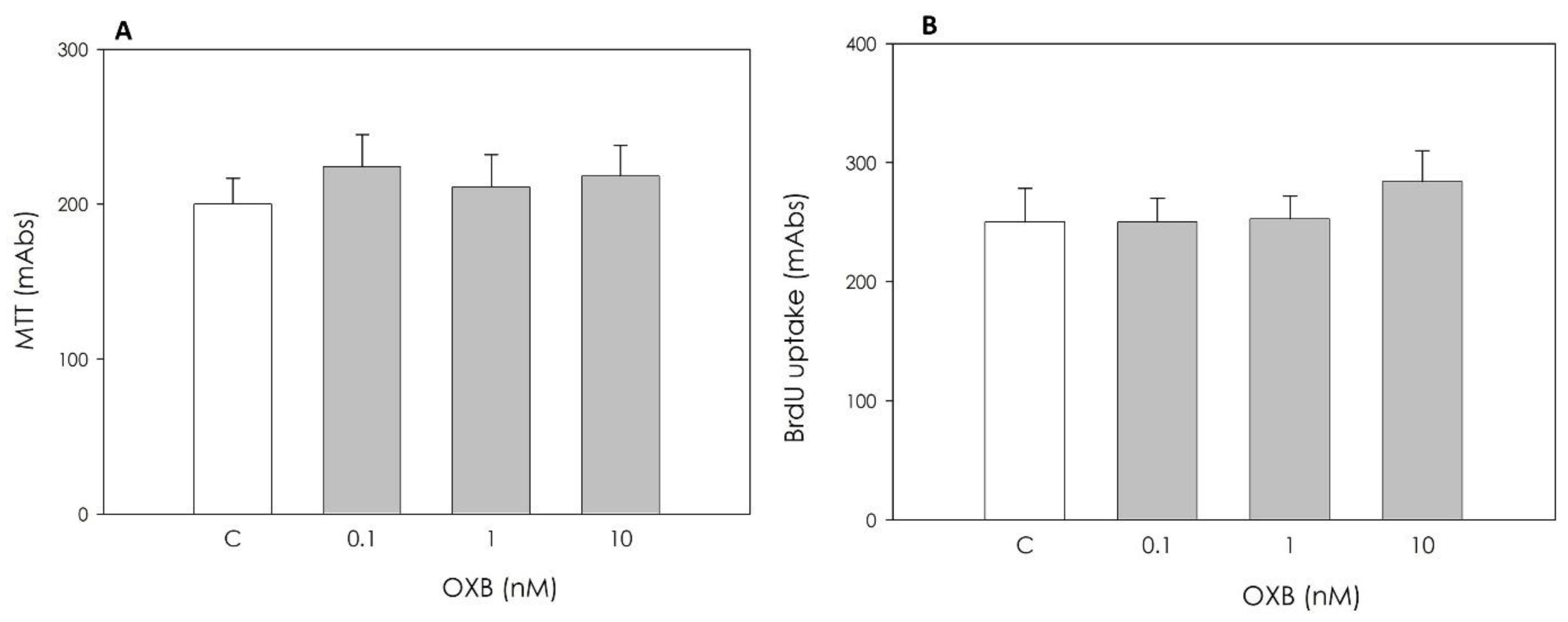
2.3.1. Granulosa Cell Viability and Proliferation
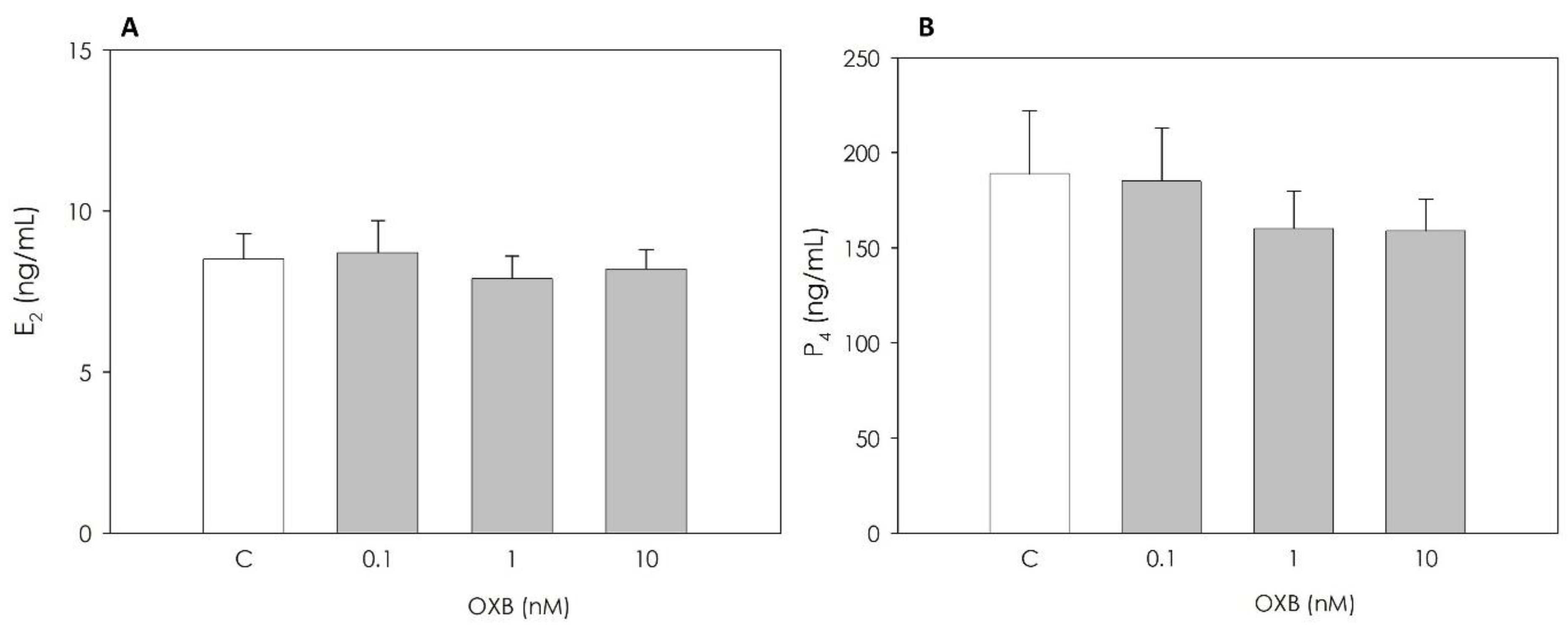
2.3.2. Granulosa Cell Steroid Production
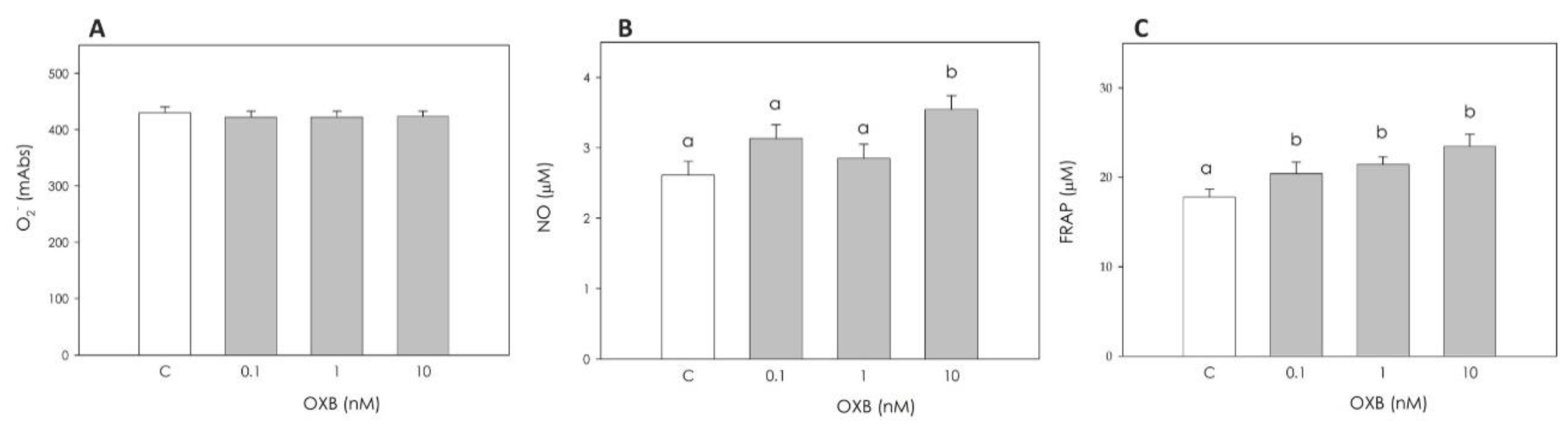
2.3.3. Granulosa Cell Redox Status
Granulosa Cell Superoxide (O2−) Production
Granulosa Cell Nitric Oxide (NO) Production
Granulosa Cell Non-Enzymatic Scavenging Activity
2.4. Evaluation of OXB Effects on Swine Aortic Endothelial Cell (AOC) Function
2.4.1. AOC Culture
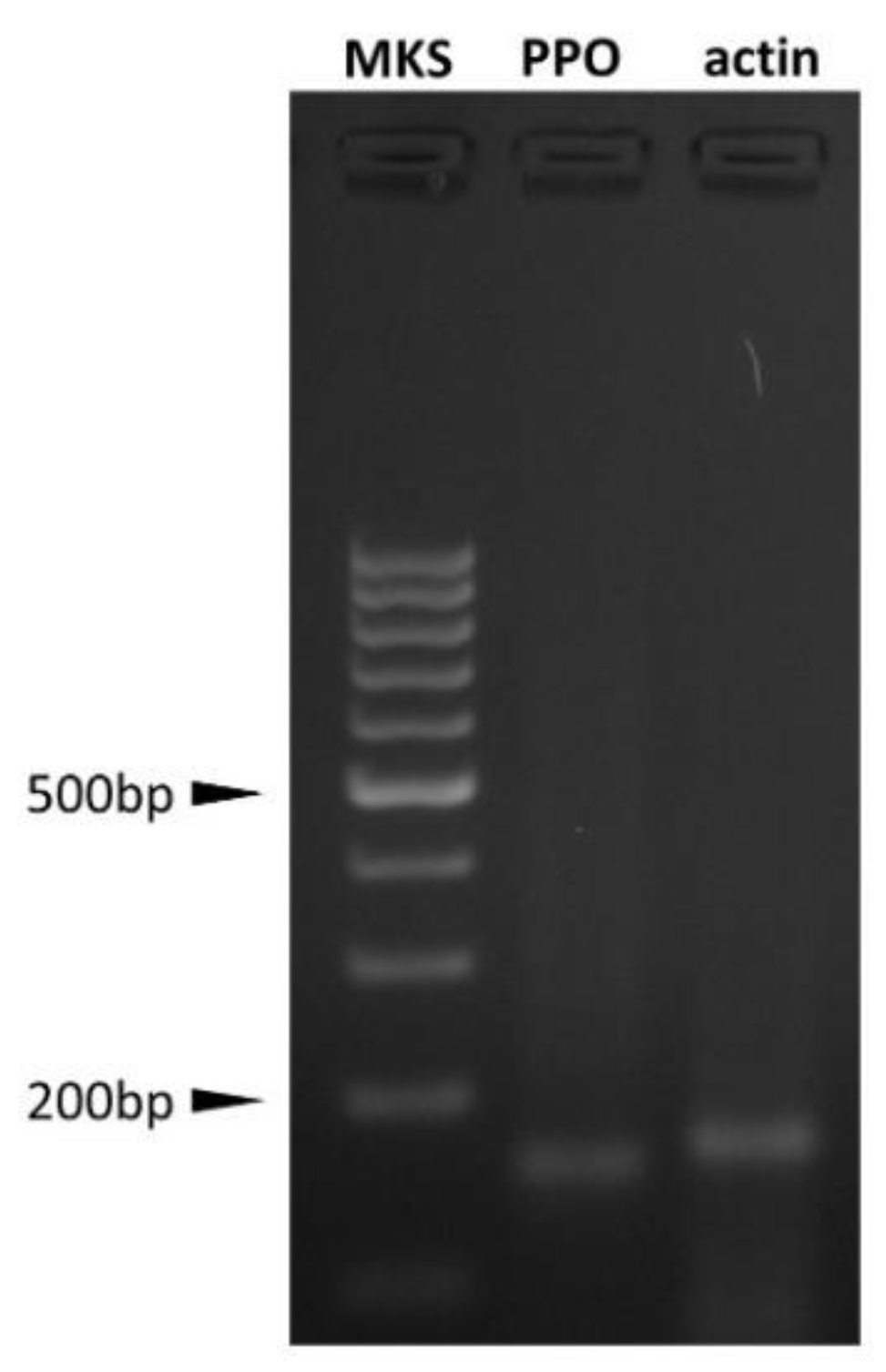
2.4.2. AOC Prepro-Orexin Expression Evaluation
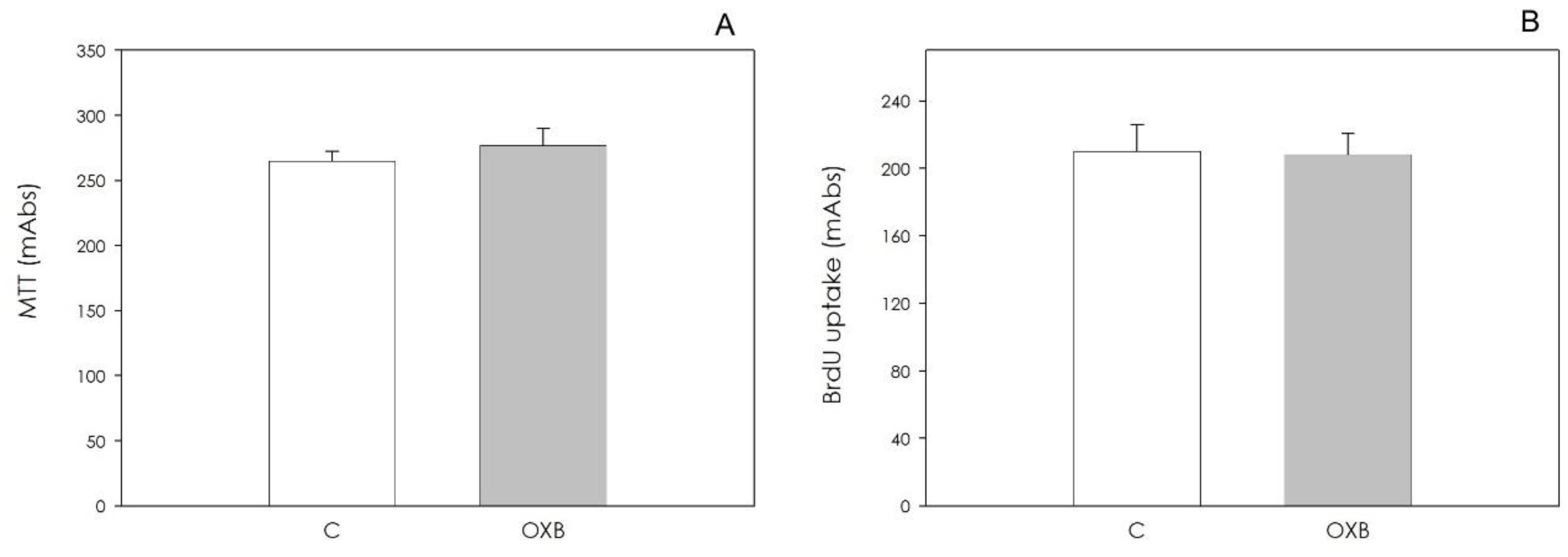
2.4.3. AOC Viability and Proliferation
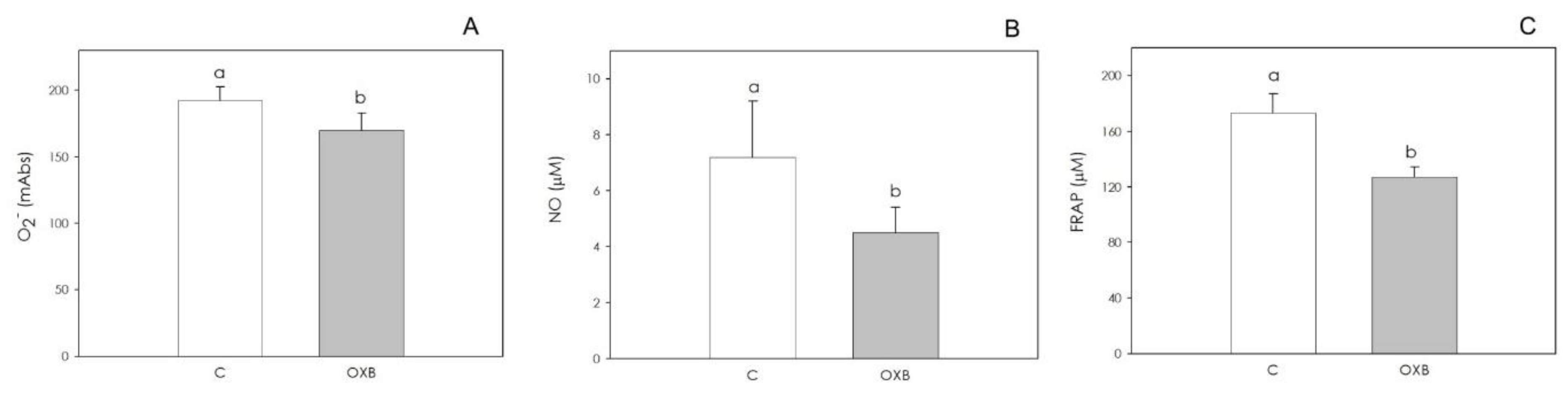
2.4.4. AOC Redox Status
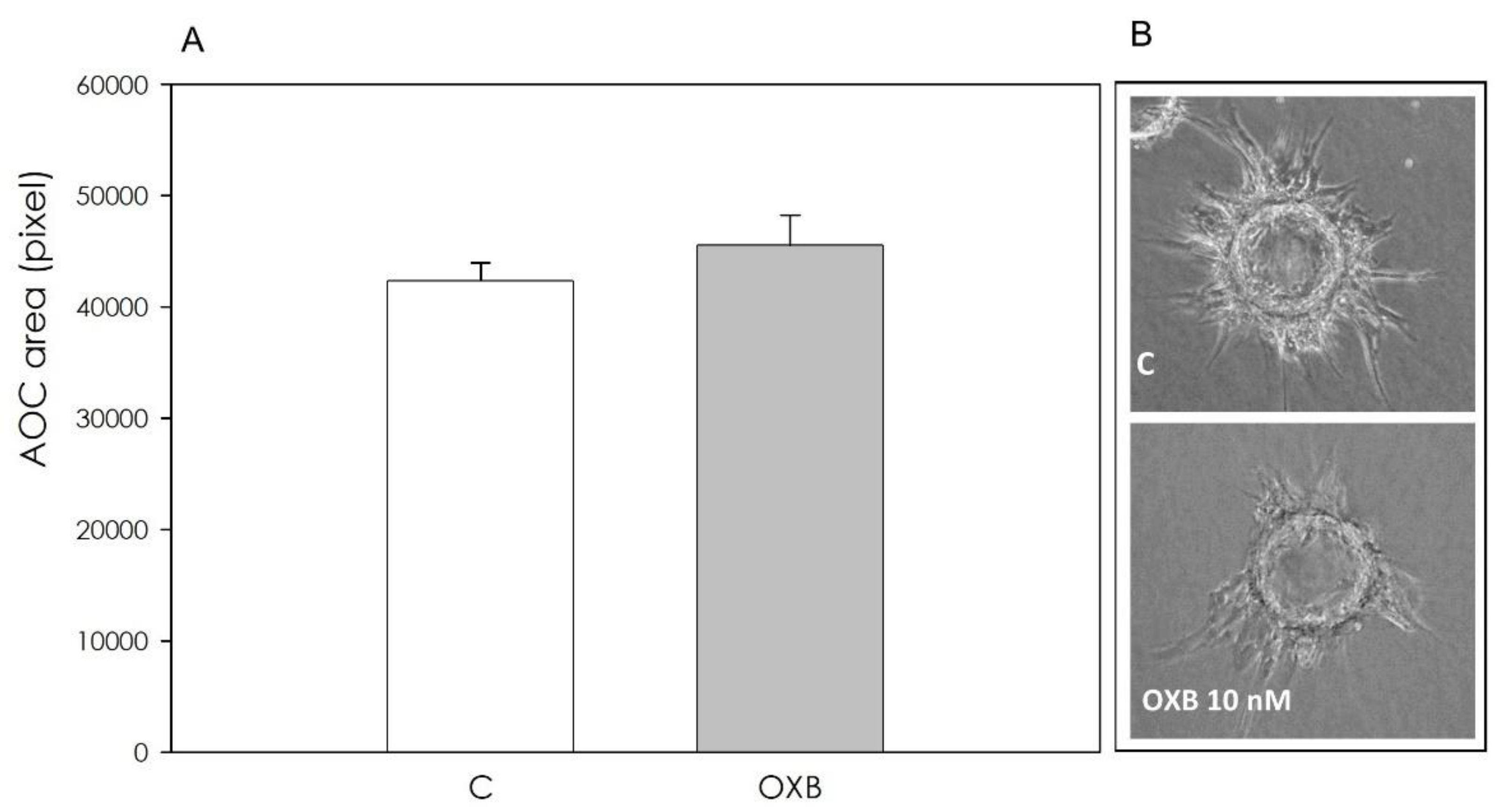
2.4.5. Angiogenesis Bioassay
2.5. Statistical Analysis
3. Results
3.1. Immunolocalization of OXB and Its Effects on Swine Granulosa Cell Function
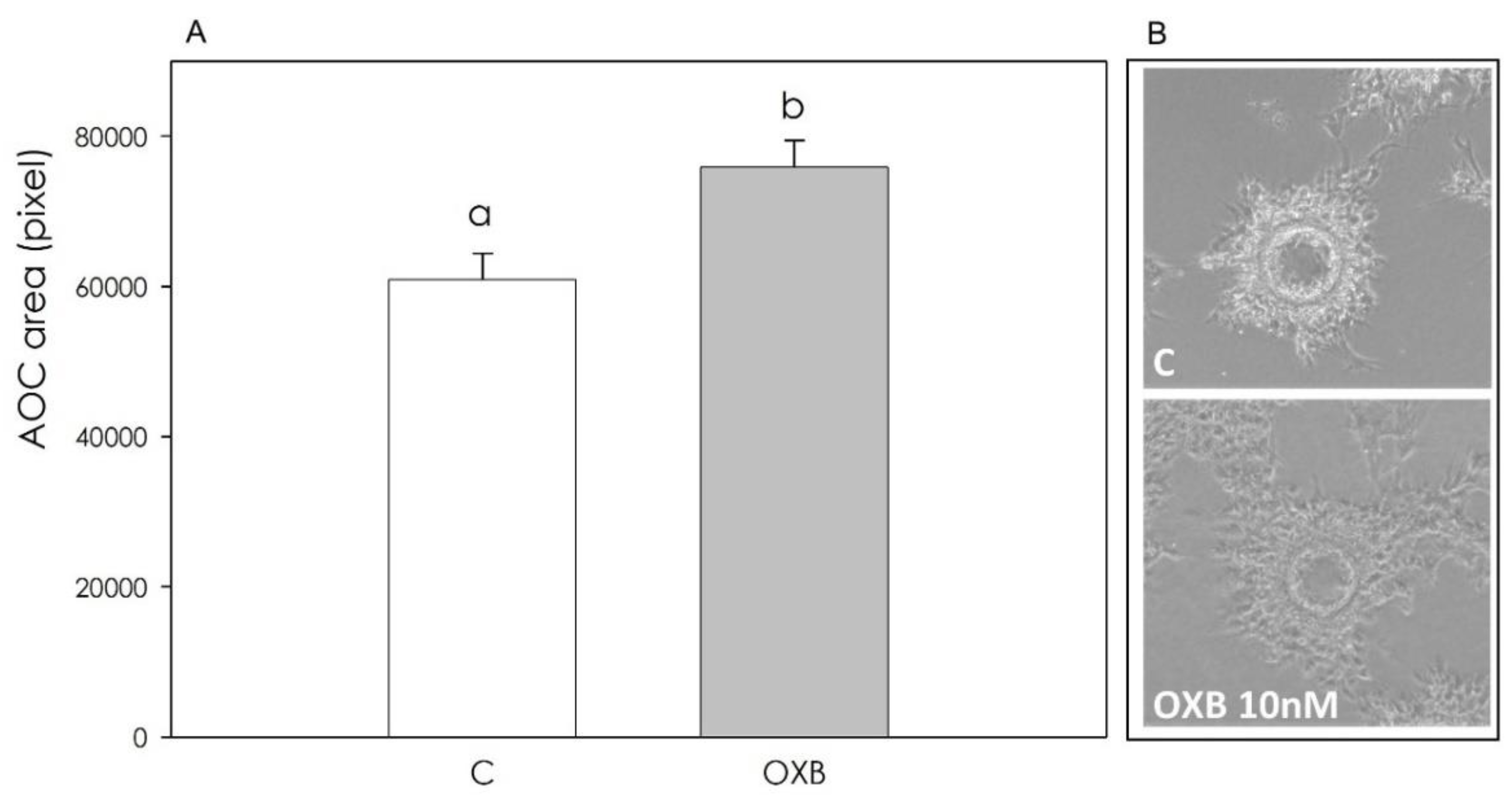
3.2. PPO Expression and OXB Effects on Swine Aortic Endothelial Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Lecea, L.; Kilduff, T.S.; Peyron, C.; Gao, X.; Foye, P.E.; Danielson, P.E.; Fukuhara, C.; Battenberg, E.L.; Gautvik, V.T.; Bartlett, F.S., 2nd; et al. The hypocretins: Hypothalamus-specific peptides with neuroexcitatory activity. Proc. Natl. Acad. Sci. USA 1998, 95, 322–327. [Google Scholar] [CrossRef]
- Sakurai, T.; Amemiya, A.; Ishii, M.; Matsuzaki, I.; Chemelli, R.M.; Tanaka, H.; Williams, S.C.; Richardson, J.A.; Kozlowski, G.P.; Wilson, S.; et al. Orexins and orexin receptors: A family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 1998, 92, 573–585. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Q.; Liu, A.; Lan, X.; Huang, Y.; Zhao, Z.; Jie, H.; Chen, J.; Zhao, Y. Physiological Implications of Orexins/Hypocretins on Energy Metabolism and Adipose Tissue Development. ACS Omega 2019, 5, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T. The neuralcircuit of orexin (hypocretin): Maintaining sleep and wakefulness. Nat. Rev. Neurosci. 2007, 8, 171–181. [Google Scholar] [CrossRef]
- Chieffi, S.; Carotenuto, M.; Monda, V.; Valenzano, A.; Villano, I.; Precenzano, F.; Tafuri, D.; Salerno, M.; Filippi, N.; Nuccio, F.; et al. Orexin System: The Key for a Healthy Life. Front. Physiol. 2017, 8, 357. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.P.; Liu, C.; Liu, M.F.; Wang, Y.; Bian, K.; Xue, Y.; Chen, L. Involvement of orexin-A in the regulation of neuronal activity and emotional behaviors in central amygdala in rats. Neuropeptides 2020, 80, 102019. [Google Scholar] [CrossRef]
- Basini, G.; Bussolati, S.; Grolli, S.; Ragionieri, L.; Di Lecce, R.; Grasselli, F. Orexin B inhibits viability and differentiation of stromal cells from swine adipose tissue. Domest. Anim. Endocrinol. 2021, 75, 106594. [Google Scholar] [CrossRef]
- Gigante, P.; Berni, M.; Bussolati, S.; Grasselli, F.; Grolli, S.; Ramoni, R.; Basini, G. Glyphosate affects swine ovarian and adipose stromal cell functions. Anim. Reprod. Sci. 2018, 195, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Berni, M.; Gigante, P.; Bussolati, S.; Grasselli, F.; Grolli, S.; Ramoni, R.; Basini, G. Bisphenol S, a Bisphenol A alternative, impairs swine ovarian and adipose cell functions. Domest. Anim. Endocrinol. 2019, 66, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Silveyra, P.; Cataldi, N.I.; Lux-Lantos, V.A.; Libertun, C. Role of orexins in the hypothalamic-pituitary-ovarian relationships. Acta Physiol. 2010, 198, 355–360. [Google Scholar] [CrossRef]
- Nitkiewicz, A.; Smolinska, N.; Przala, J.; Kaminski, T. Expression of orexin receptors 1 (OX1R) and 2 (OX2R) in the porcine ovary during the oestrous cycle. Regul. Pept. 2010, 165, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Nitkiewicz, A.; Smolinska, N.; Maleszka, A.; Chojnowska, K.; Kaminski, T. Expression of orexins and their precursor in the porcine ovary and the influence of orexins on ovarian steroidogenesis in pigs. Anim. Reprod. Sci. 2014, 148, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, T.; Smolinska, N.; Nitkiewicz, A.; Przala, J. Expression of orexin receptors 1 (OX1R) and 2 (OX2R) in the porcine hypothalamus during the oestrous cycle. J. Physiol. Pharmacol. 2010, 61, 363–371. [Google Scholar]
- Levanti, M.; Germanà, A.; Abbate, F. Orexin A expression in the ovary of dog and cat. Reprod. Domest. Anim. 2015, 50, 247–250. [Google Scholar] [CrossRef]
- Ragionieri, L.; Ravanetti, F.; Di Lecce, R.; Botti, M.; Ciccimarra, R.; Bussolati, S.; Basini, G.; Gazza, F.; Cacchioli, A. Immunolocalization of Orexin A and its receptors in the different structures of the porcine ovary. Ann. Anat. 2018, 218, 214–226. [Google Scholar] [CrossRef]
- Basini, G.; Ciccimarra, R.; Bussolati, S.; Grolli, S.; Ragionieri, L.; Ravanetti, F.; Botti, M.; Gazza, F.; Cacchioli, A.; Di Lecce, R.; et al. Orexin A in swine corpus luteum. Domest. Anim. Endocrinol. 2018, 64, 38–48. [Google Scholar] [CrossRef]
- Ciccimarra, R.; Bussolati, S.; Grasselli, F.; Grolli, S.; Ragionieri, L.; Ravanetti, F.; Botti, M.; Gazza, F.; Cacchioli, A.; Di Lecce, R.; et al. Orexin system in swine ovarian follicles. Domest. Anim. Endocrinol. 2018, 62, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, T.; Smolinska, N.; Kiezun, M.; Dobrzyn, K.; Szeszko, K.; Maleszka, A. Effect of orexin B on CYP17A1 and CYP19A3 expression and oestradiol, oestrone and testosterone secretion in the porcine uterus during early pregnancy and the oestrous cycle. Animal 2018, 12, 1921–1932. [Google Scholar] [CrossRef]
- Kisielewska, K.; Rytelewska, E.; Gudelska, M.; Kiezun, M.; Dobrzyn, K.; Szeszko, K.; Bors, K.; Wyrebek, J.; Kaminski, T.; Smolinska, N. The effect of orexin B on steroidogenic acute regulatory protein, P450 side-chain cleavage enzyme, and 3 beta-hydroxysteroid dehydrogenase gene expression, and progesterone and androstenedione secretion by the porcine uterus during early pregnancy and the estrous cycle. J. Anim. Sci. 2019, 97, 851–864. [Google Scholar] [PubMed]
- Liguori, G.; Tafuri, S.; Miyoshi, C.; Yanagisawa, M.; Squillacioti, C.; De Pasquale, V.; Mirabella, N.; Vittoria, A.; Costagliola, A. Localization of orexin B and orexin-2 receptor in the rat epididymis. Acta Histochem. 2018, 120, 292–297. [Google Scholar] [CrossRef]
- Cataldi, N.I.; Lux-Lantos, V.A.; Libertun, C. Effects of orexins A and B on expression of orexin receptors and progesterone release in luteal and granulosa ovarian cells. Regul. Pept. 2012, 178, 56–63. [Google Scholar] [CrossRef]
- Basini, G.; Bussolati, S.; Ciccimarra, R.; Grasselli, F. Melatonin potentially acts directly on swine ovary by modulating granulosa cell function and angiogenesis. Reprod. Fertil. Dev. 2017, 29, 2305–2312. [Google Scholar] [CrossRef] [PubMed]
- Basini, G.; Bussolati, S.; Santini, S.E.; Grasselli, F. Sanguinarine inhibits VEGF-induced angiogenesis in a fibrin gel matrix. BioFactors 2007, 29, 11–18. [Google Scholar] [CrossRef]
- Babalola, G.O.; Shapiro, B.H. Correlation of follicular steroid hormone profiles with ovarian cyclicity in sows. Reproduction 1988, 84, 79–87. [Google Scholar] [CrossRef]
- Akins, E.L.; Morrissette, M.C. Gross ovarian changes during estrous cycle of swine. Am. J. Vet. Res. 1968, 29, 1953–1957. [Google Scholar]
- Dall’Aglio, C.; Zannoni, A.; Forni, M.; Bacci, M.L.; Ceccarelli, P.; Boiti, C. Orexin system expression in the gastrointestinal tract of pigs. Res. Vet. Sci. 2013, 95, 8–14. [Google Scholar] [CrossRef]
- Grasselli, F.; Basini, G.; Tirelli, M.; Cavalli, V.; Bussolati, S.; Tamanini, C. Angiogenic activity of porcine granulosa cells cocultured with endothelial cells in a microcarrier-based three-dimensional fibrin gel. J. Physiol. Pharmacol. 2003, 54, 361–370. [Google Scholar] [PubMed]
- Basini, G.; Baioni, L.; Bussolati, S.; Grolli, S.; Grasselli, F. Prolactin is a potential physiological modulator of swine ovarian follicle function. Regul. Pept. 2014, 189, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Basini, G.; Ragionieri, L.; Bussolati, S.; Di Lecce, R.; Cacchioli, A.; Dettin, M.; Cantoni, A.M.; Grolli, S.; La Bella, O.; Zamuner, A.; et al. Expression and function of the stromal cell-derived factor-1 (SDF-1) and CXC chemokine receptor 4 (CXCR4) in the swine ovarian follicle. Domest. Anim. Endocrinol. 2020, 71, 106404. [Google Scholar] [CrossRef] [PubMed]
- Foxcroft, G.R.; Hunter, M.G. Basic physiology of follicular maturation in the pig. J. Reprod. Fertil. Suppl. 1985, 33, 1–19. [Google Scholar]
- Kaminski, T.; Nitkiewicz, A.; Smolinska, N. Changes in plasma orexinA and orexin B concentrations during the oestrous cycle of the pig. Peptides 2013, 39, 175–177. [Google Scholar]
- Basini, G.; Bussolati, S.; Iannarelli, M.; Ragionieri, L.; Grolli, S.; Ramoni, R.; Dodi, A.; Gazza, F.; Grasselli, F. The myokine irisin: Localization and effects in swine late medium and large antral ovarian follicle. Domest. Anim. Endocrinol. 2021, 74, 106576. [Google Scholar] [CrossRef] [PubMed]
- Basini, G.; Tamanini, C. Selenium stimulates estradiol production in bovine granulosa cells: Possible involvement of nitric oxide. Domest. Anim. Endocrinol. 2000, 18, 1–17. [Google Scholar] [CrossRef]
- Pacentra, A.; Grasselli, F.; Bussolati, S.; Grolli, S.; Di Lecce, R.; Cantoni, A.M.; Basini, G. The effect of pathogen-associated molecular patterns on the swine granulosa cells. Theriogenology 2020, 145, 207–216. [Google Scholar] [CrossRef]
- Basini, G.; Bussolati, S.; Baioni, L.; Grasselli, F. Gossypol, a polyphenolic aldehyde from cotton plant, interferes with swine granulosa cell function. Domest. Anim. Endocrinol. 2009, 37, 30–36. [Google Scholar] [CrossRef]
- Basini, G.; Baioni, L.; Bussolati, S.; Grolli, S.; Kramer, L.H.; Wagner, G.F.; Grasselli, F. Expression and localization of stanniocalcin 1 in swine ovary. Gen. Comp. Endocrinol. 2010, 166, 404–408. [Google Scholar] [CrossRef]
- Butcher, J.T.; Tressel, S.; Johnson, T.; Turner, D.; Sorescu, G.; Jo, H.; Nerem, R.M. Transcriptional Profiles of Valvular and Vascular Endothelial Cells Reveal Phenotypic Differences. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Basini, G.; Bussolati, S.; Santini, S.E.; Bianchi, F.; Careri, M.; Mangia, A.; Musci, M.; Grasselli, F. Hydroxyestrogens inhibit angiogenesis in swine ovarian follicles. J. Endocrinol. 2008, 199, 127–135. [Google Scholar] [CrossRef]
- Russell, S.H.; Small, C.J.; Kennedy, A.R.; Stanley, S.A.; Seth, A.; Murphy, K.G.; Taheri, S.; Ghatei, M.A.; Bloom, S.R. Orexin A interactions in the hypothalamus-pituitary-gonadal axis. Endocrinology 2001, 142, 5294–5302. [Google Scholar] [CrossRef] [PubMed]
- Kok, S.W.; Roelfsema, F.; Overeem, S.; Lammers, G.J.; Frölich, M.; Meinders, A.E.; Pijl, H. Pulsatile LH release is diminished, whereas FSH secretion is normal, in hypocretin-deficient narcoleptic men. Am. J. Physiol. Endocrinol. Metab. 2004, 287, E630–E636. [Google Scholar] [CrossRef] [PubMed]
- Barb, C.R.; Matteri, R. Orexin-B modulates luteinizing hormone and growth hormone secretion from porcine pituitary cells in culture. Domest. Anim. Endocrinol. 2005, 28, 331–337. [Google Scholar] [CrossRef]
- Scammell, T.E.; Winrow, C.J. Orexin receptors: Pharmacology and therapeutic opportunities. Annu. Rev. Pharmacol. Toxicol. 2011, 51, 243–266. [Google Scholar] [CrossRef]
- Sokołowska, P.; Urbańska, A.; Biegańska, K.; Wagner, W.; Ciszewski, W.; Namiecińska, M.; Zawilska, J.B. Orexins protect neuronal cell cultures against hypoxic stress: An involvement of Akt signaling. J. Mol. Neurosci. 2014, 52, 48–55. [Google Scholar] [CrossRef]
- Couvineau, A.; Voisin, T.; Nicole, P.; Gratio, V.; Abad, C.; Tan, Y.V. Orexins as Novel Therapeutic Targets in Inflammatory and Neurodegenerative Diseases. Front. Endocrinol. 2019, 22, 709. [Google Scholar] [CrossRef] [PubMed]
- Basini, G.; Grasselli, F. Nitric oxide in follicle development and oocyte competence. Reproduction 2015, 150, R1–R9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Abdukerim, M.; Abilailieti, M.; Tang, L.; Ling, Y.; Pan, S. The protective effects of orexin a against high glucose-induced activation of NLRP3 inflammasome in human vascular endothelial cells. Arch. Biochem. Biophys. 2019, 672, 108052. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.W.; Byzova, T.V. Oxidative stress in angiogenesis and vascular disease. Blood 2014, 123, 625–631. [Google Scholar] [CrossRef]
- Johren, O.; Neidert, S.J.; Kummer, M.; Dendorfer, A.; Dominiak, P. Prepro-orexin and orexin receptor mRNAs are differentially expressed in peripheral tissues of male and female rats. Endocrinology 2001, 142, 3324–3331. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Park, H.J.; Kim, S.R.; Choi, Y.K.; Shin, H.K.; Jeon, J.H.; Jang, H.O.; Yun, I.; Bae, S.K.; Bae, M.K. Angiogenic role of orexin-A via the activation of extracellular signal-regulated kinase in endothelial cells. Biochem. Biophys. Res. Commun. 2010, 403, 59–65. [Google Scholar] [CrossRef]
- Heinonen, M.V.; Purhonen, A.K.; Mäkelä, K.A.; Herzig, K.H. Functions of orexins in peripheral tissues. Acta Physiol. 2008, 192, 471–485. [Google Scholar] [CrossRef]
- Kim, M.K.; Park, H.J.; Kim, S.R.; Choi, Y.K.; Bae, S.K.; Bae, M.K. Involvement of Heme Oxygenase-1 in Orexin-A-induced Angiogenesis in Vascular Endothelial Cells. Korean J. Physiol. Pharmacol. 2015, 19, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Berisha, B.; Schams, D.; Rodler, D.; Pfaffl, M.W. Angiogenesis in The Ovary—The Most Important Regulatory Event for Follicle and Corpus Luteum Development and Function in Cow—An Overview. Anat. Histol. Embryol. 2016, 45, 124–130. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grasselli, F.; Bussolati, S.; Grolli, S.; Di Lecce, R.; Dall’Aglio, C.; Basini, G. Effects of Orexin B on Swine Granulosa and Endothelial Cells. Animals 2021, 11, 1812. https://doi.org/10.3390/ani11061812
Grasselli F, Bussolati S, Grolli S, Di Lecce R, Dall’Aglio C, Basini G. Effects of Orexin B on Swine Granulosa and Endothelial Cells. Animals. 2021; 11(6):1812. https://doi.org/10.3390/ani11061812
Chicago/Turabian StyleGrasselli, Francesca, Simona Bussolati, Stefano Grolli, Rosanna Di Lecce, Cecilia Dall’Aglio, and Giuseppina Basini. 2021. "Effects of Orexin B on Swine Granulosa and Endothelial Cells" Animals 11, no. 6: 1812. https://doi.org/10.3390/ani11061812
APA StyleGrasselli, F., Bussolati, S., Grolli, S., Di Lecce, R., Dall’Aglio, C., & Basini, G. (2021). Effects of Orexin B on Swine Granulosa and Endothelial Cells. Animals, 11(6), 1812. https://doi.org/10.3390/ani11061812