
No Observed Adverse Effects on Health Were Detected in Adult Beagle Dogs When Fed a High-Calcium Diet for 40 Weeks

 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Dogs and Housing
2.3. Diets
2.4. Measures
2.5. Intake, BW, and BCS
2.6. Mineral Balance and Apparent Digestibility
2.7. Urine Mineral Content and RSS
2.8. Urinalysis
2.9. Blood Based Measures
2.10. GFR
2.11. Imaging
2.12. Statistical Powering
2.13. Statistical Analysis
3. Results
3.1. General
3.2. Intake and BW
3.3. Ca, P, Regulatory Hormones and Vitamin D Metabolites in Blood
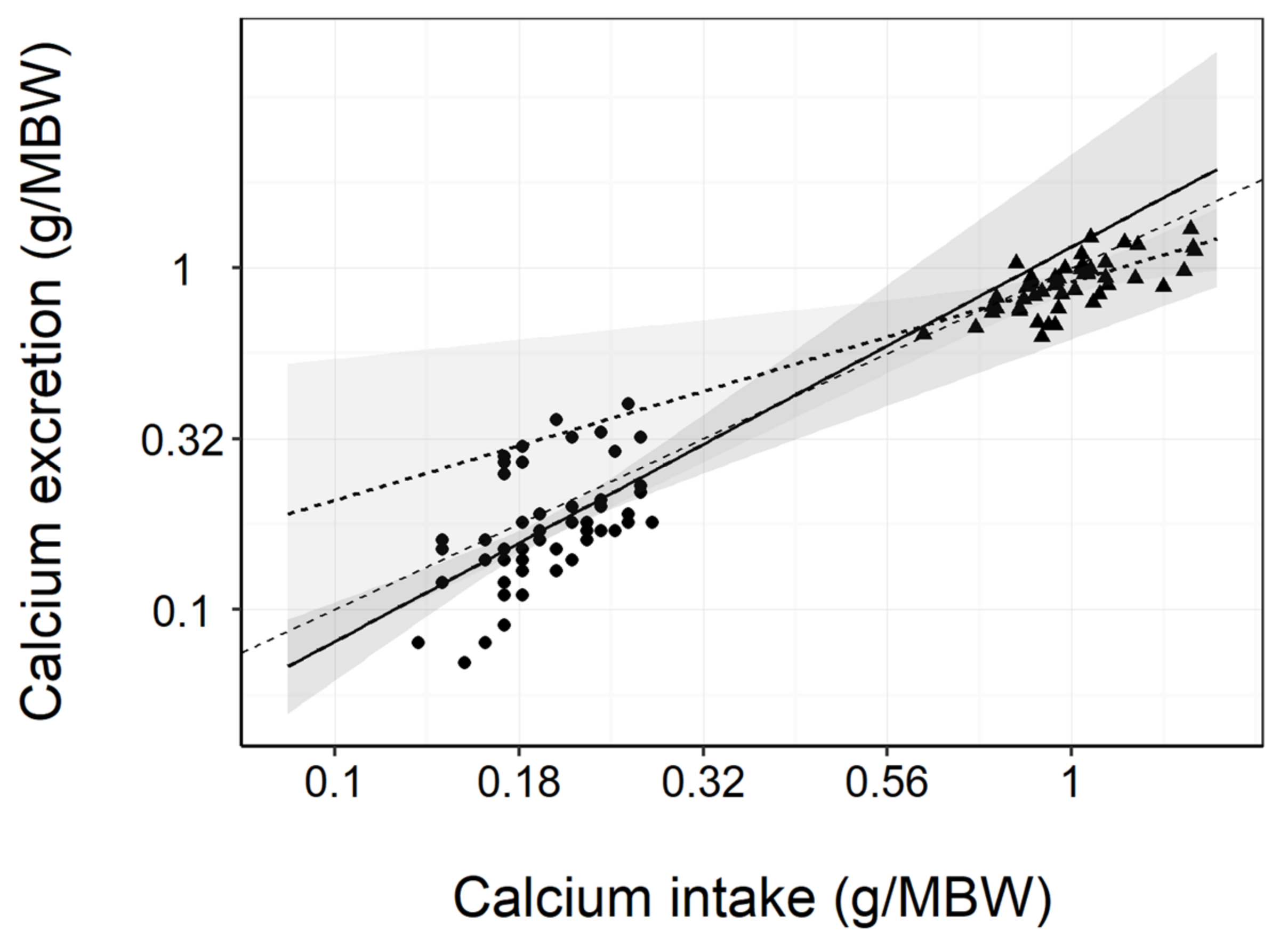
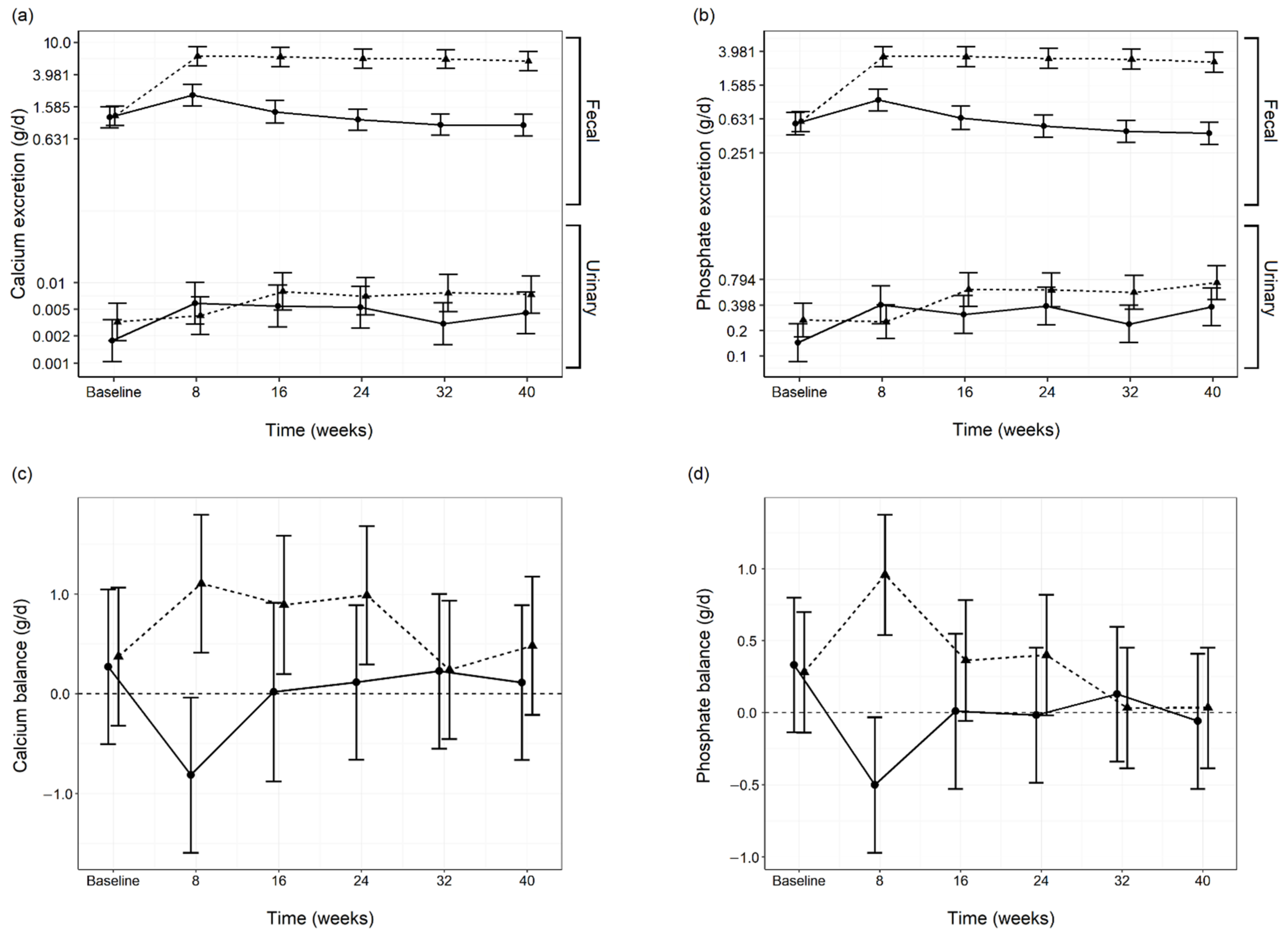
3.4. Diet Digestibility and Mineral Balance
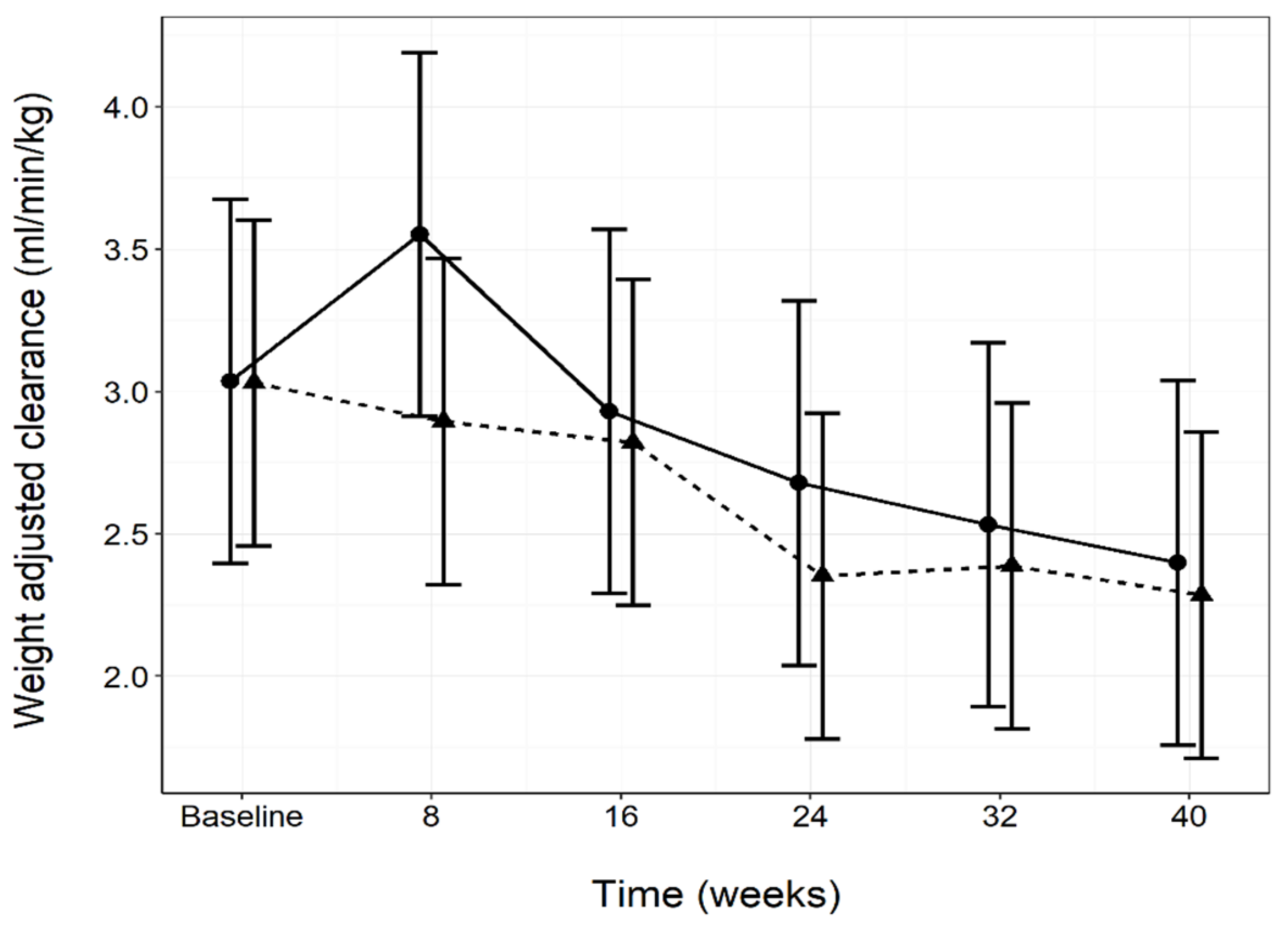
3.5. Renal and Urinary Health
3.6. Bone Mineralization and Related Marker Measurements
3.7. Imaging
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Brito Galvao, J.F.; Nagode, L.A.; Schenck, P.A.; Chew, D.J. Calcitriol, calcidiol, parathyroid hormone, and fibroblast growth factor-23 interactions in chronic kidney disease. J. Vet. Emerg. Crit. Care 2013, 23, 134–162. [Google Scholar] [CrossRef] [PubMed]
- Jeon, U.S. Kidney and calcium homeostasis. Electrolyte Blood Press 2008, 6, 68–76. [Google Scholar] [CrossRef]
- Dobenecker, B.; Kasbeitzer, N.; Flinspach, S.; Köstlin, R.; Matis, U.; Kienzle, E. Calcium-excess causes subclinical changes of bone growth in Beagles but not in Foxhound-crossbred dogs, as measured in X-rays. J. Anim. Physiol. Anim. Nutr. 2006, 90, 394–401. [Google Scholar] [CrossRef]
- Nap, R.C.; Hazewinkel, H.A. Growth and skeletal development in the dog in relation to nutrition; a review. Vet. Q. 1994, 16, 50–59. [Google Scholar] [CrossRef]
- Schoenmakers, I.; Hazewinkel, H.A.; Voorhout, G.; Carlson, C.S.; Richardson, D. Effects of diets with different calcium and phosphorus contents on the skeletal development and blood chemistry of growing great danes. Vet. Rec. 2000, 147, 652–660. [Google Scholar]
- Goedegebuure, S.A.; Hazewinkel, H.A. Morphological findings in young dogs chronically fed a diet containing excess calcium. Vet. Pathol. 1986, 23, 594–605. [Google Scholar] [CrossRef]
- Cortadellas, O.; Fernández del Palacio, M.J.; Talavera, J.; Bayón, A. Calcium and Phosphorus Homeostasis in Dogs with Spontaneous Chronic Kidney Disease at Different Stages of Severity. J. Vet. Intern. Med. 2010, 24, 73–79. [Google Scholar] [CrossRef]
- Lulich, J.P.; Osborne, C.A.; Nagode, L.A.; Polzin, D.J.; Parke, M.L. Evaluation of urine and serum metabolites in miniature schnauzers with calcium oxalate urolithiasis. Am. J. Vet. Res. 1991, 52, 1583–1590. [Google Scholar]
- Dobenecker, B. Factors that modify the effect of excess calcium on skeletal development in puppies. Br. J. Nutr. 2011, 106, S142–S145. [Google Scholar] [CrossRef] [PubMed]
- Kiefer-Hecker, B.; Kienzle, E.; Dobenecker, B. Effects of low phosphorus supply on the availability of calcium and phosphorus, and musculoskeletal development of growing dogs of two different breeds. J. Anim. Physiol. Anim. Nutr. 2018, 102, 789–798. [Google Scholar] [CrossRef]
- Laflamme, G.H.; Jowsey, J. Bone and soft tissue changes with oral phosphate supplements. J. Clin. Investig. 1972, 51, 2834–2840. [Google Scholar] [CrossRef]
- Cook, S.D.; Skinner, H.B.; Haddad, R.J. A quantitative histologic study of osteoporosis produced by nutritional secondary hyperparathyroidism in dogs. Clin. Orthop. Relat. Res. 1983, 175, 105–120. [Google Scholar] [CrossRef]
- Saville, P.D.; Krook, L. Gravimetric and isotopic studies in nutritional hyperparathyroidism in beagles. Clin. Orthop. Relat. Res. 1969, 62, 15–24. [Google Scholar] [CrossRef]
- Cloutier, M.; Gascon-Barré, M.; D’Amour, P. Chronic adaptation of dog parathyroid function to a low-calcium-high-Sodium-Vitamin D-deficient diet. J. Bone Miner. Res. 1992, 7, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- Stockman, J.; Villaverde, C.; Corbee, R.J. Calcium, Phosphorus, and Vitamin D in Dogs and Cats: Beyond the Bones. Vet. Clin. N. Am. Small Anim. Pract. 2021, 51, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Mack, J.K.; Alexander, L.G.; Morris, P.J.; Dobenecker, B.; Kienzle, E. Demonstration of uniformity of calcium absorption in adult dogs and cats. J. Anim. Physiol. Anim. Nutr. 2015, 99, 801–809. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Dogs and Cats; National Academies Press: Washington, DC, USA, 2006.
- Schmitt, S.; Mack, J.; Kienzle, E.; Alexander, L.G.; Morris, P.J.; Colyer, A.; Dobenecker, B. Faecal calcium excretion does not decrease during long-term feeding of a low-calcium diet in adult dogs. J. Anim. Physiol. Anim. Nutr. 2018, 102, e798–e805. [Google Scholar] [CrossRef] [PubMed]
- Wilkens, M.R.; Richter, J.; Fraser, D.R.; Liesegang, A.; Breves, G.; Schroder, B. In contrast to sheep, goats adapt to dietary calcium restriction by increasing intestinal absorption of calcium. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2012, 163, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Liesegang, A.; Sassi, M.; Risteli, J.; Kraenzlin, M.; Riond, J.; Wanner, M. Influence of low calcium, high calcium and energy-rich diets on bone markers in ovariectomized Beagle dogs. J. Anim. Physiol. Anim. Nutr. 1999, 81, 223–231. [Google Scholar]
- Stephens, L.C.; Norrdin, R.W.; Benjamin, S.A. Effects of calcium supplementation and sunlight exposure on growing beagle dogs. Am. J. Vet. Res. 1985, 46, 2037–2042. [Google Scholar]
- Furrow, E.; Patterson, E.E.; Armstrong, P.J.; Osborne, C.A.; Lulich, J.P. Fasting urinary calcium-to-creatinine and oxalate-to-creatinine ratios in dogs with calcium oxalate urolithiasis and breed-matched controls. J. Vet. Intern. Med. 2015, 29, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Lulich, J.P.; Osborne, C.A.; Polzin, D.J.; Johnston, S.D.; Parker, M.L. Urine metabolite values in fed and nonfed clinically normal beagles. Am. J. Vet. Res. 1991, 52, 1573–1578. [Google Scholar] [PubMed]
- Slater, M.R.; Scarlett, J.M.; Donoghue, S.; Kaderly, R.E.; Bonnett, B.N.; Cockshutt, J.; Erb, H.N. Diet and exercise as potential risk factors for osteochondritis dissecans in dogs. Am. J. Vet. Res. 1992, 53, 2119–2124. [Google Scholar] [PubMed]
- Association of American Feed Control Officials (AAFCO). 2018 Official Publication; The Association of Feed Control Officials Inc.: Washington, DC, USA, 2018. [Google Scholar]
- European Pet Food Industry Federation (FEDIAF). Nutritional Guidelines for Complete and Complementary Pet Food for Cats and Dogs; FEDIAF: Brussels, Belgium, 2019. [Google Scholar]
- Stockman, J.; Watson, P.; Gilham, M.; Allaway, D.; Atwal, J.; Haydock, R.; Colyer, A.; Renfrew, H.; Morris, P.J. Adult dogs are capable of regulating calcium balance, with no adverse effects on health, when fed a high-calcium diet. Br. J. Nutr. 2017, 117, 1235–1243. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Matsuzaki, H.; Kikuchi, T.; Kajita, Y.; Masuyama, R.; Uehara, M.; Goto, S.; Suzuki, K. Comparison of various phosphate salts as the dietary phosphorus source on nephrocalcinosis and kidney function in rats. J. Nutr. Sci. Vitam. 1999, 45, 595–608. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pastoor, F.J.; Van’t Klooster, A.T.; Mathot, J.N.; Beynen, A.C. Increasing phosphorus intake reduces urinary concentrations of magnesium and calcium in adult ovariectomized cats fed purified diets. J. Nutr. 1995, 125, 1334–1341. [Google Scholar]
- Nadkarni, G.N.; Uribarri, J. Phosphorus and the Kidney: What Is Known and What Is Needed. Adv. Nutr. 2014, 5, 98–103. [Google Scholar] [CrossRef] [PubMed]
- German, A.J.; Holden, S.L.; Moxham, G.L.; Holmes, K.L.; Hackett, R.M.; Rawlings, J.M. A simple, reliable tool for owners to assess the body condition of their dog or cat. J. Nutr. 2006, 136, 2031s–2033s. [Google Scholar] [CrossRef]
- Association of American Feed Control Officials (AAFCO). 2013 Official Publication; The Association of Feed Control Officials Inc.: Washington, DC, USA, 2013. [Google Scholar]
- Kienzle, E.; Wilms-Eilers, S. Struvite Diet in Cats: Effect of Ammonium Chloride and Carbonates on Acid Base Balance of Cats. J. Nutr. 1994, 124, 2652S–2659S. [Google Scholar] [CrossRef]
- Laflamme, D.P. Determining metabolizable energy content in commercial pet foods. J. Anim. Physiol. Anim. Nutr. 2001, 85, 222–230. [Google Scholar] [CrossRef]
- European Union, Regulation (EC) N °152/of the commission of 27 January 2009. Off. J. Eur. Union 2009, 152, L54/1.
- Robertson, W.G.; Jones, J.S.; Heaton, M.A.; Stevenson, A.E.; Markwell, P.J. Predicting the Crystallization Potential of Urine from Cats and Dogs with Respect to Calcium Oxalate and Magnesium Ammonium Phosphate (Struvite). J. Nutr. 2002, 132, 1637S–1641S. [Google Scholar] [CrossRef]
- Estepa, J.C.; Lopez, I.; Felsenfeld, A.J.; Gao, P.; Cantor, T.; Rodríguez, M.; Aguilera-Tejero, E. Dynamics of secretion and metabolism of PTH during hypo- and hypercalcaemia in the dog as determined by the ‘intact’ and ‘whole’ PTH assays. Nephrol. Dial. Transplant. 2003, 18, 1101–1107. [Google Scholar] [CrossRef][Green Version]
- Mooney, C.T.; Shiel, R.E.; Fawcett, K.; Matthews, E.; Gunn, E. A comparison of canine whole and intact parathyroid hormone concentrations as measured by different assays. J. Small Anim. Pract. 2019, 60, 507–513. [Google Scholar] [CrossRef]
- Mooney, C.T.; Shiel, R.E.; Sekiya, M.; Dunning, M.; Gunn, E. A Preliminary Study of the Effect of Hyperadrenocorticism on Calcium and Phosphate Concentrations, Parathyroid Hormone and Markers of Bone Turnover in Dogs. Front. Vet. Sci. 2020, 7, 311. [Google Scholar] [CrossRef]
- Harjes, L.M.; Parker, V.J.; Dembek, K.; Young, G.S.; Giovaninni, L.H.; Kogika, M.M.; Chew, D.J.; Toribio, R.E. Fibroblast Growth Factor-23 Concentration in Dogs with Chronic Kidney Disease. J. Vet. Intern. Med. 2017, 31, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.C.Y.; Nicholls, H.; Piec, I.; Washbourne, C.J.; Dutton, J.J.; Jackson, S.; Greeves, J.; Fraser, W.D. Reference intervals for serum 24,25-Dihydroxyvitamin D and the ratio with 25-Hydroxyvitamin established using a newly developed LC-MS/MS method. J. Nutr. Biochem. 2017, 46, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Bexfield, N.H.; Heiene, R.; Gerritsen, R.J.; Risoen, U.; Eliassen, K.A.; Herrtage, M.E.; Michell, A.R. Glomerular filtration rate estimated by 3-sample plasma clearance of iohexol in 118 healthy dogs. J. Vet. Intern. Med. 2008, 22, 66–73. [Google Scholar] [CrossRef]
- Munday, H.S.; Booles, D.; Anderson, P.; Poore, D.W.; Earle, K.E. The Repeatability of Body Composition Measurements in Dogs and Cats using Dual Energy X-Ray Absorptiometry. J. Nutr. 1994, 124, 2619S–2621S. [Google Scholar] [CrossRef] [PubMed]
- Speakman, J.; Booles, D.; Butterwick, R. Validation of dual energy X-ray absorptiometry (DXA) by comparison with chemical analysis of dogs and cats. Int. J. Obes. 2001, 25, 439–447. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on April 2019).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. arXiv 2015, arXiv:1406.5823. Available online: https://arxiv.org/abs/1406.5823 (accessed on April 2019).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Bronner, F. Mechanisms of intestinal calcium absorption. J. Cell. Biochem. 2003, 88, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Coltherd, J.C.; Staunton, R.; Colyer, A.; Thomas, G.; Gilham, M.; Logan, D.W.; Butterwick, R.; Watson, P. Not all forms of dietary phosphorus are equal: An evaluation of postprandial phosphorus concentrations in the plasma of the cat. Br. J. Nutr. 2019, 121, 270–284. [Google Scholar] [CrossRef]
- Siedler, S.; Dobenecker, B. The source of phosphorus influences serum PTH, apparent digestibility and blood levels of calcium and phosphorus in dogs fed high phosphorus diets with balanced Ca/P ratio. In Proceedings of the Waltham International Nutritional Sciences Symposium, Chicago, IL, USA, 18–21 October 2016. [Google Scholar]
- Liesegang, A.; Reutter, R.; Sassi, M.L.; Risteli, J.; Kraenzlin, M.; Riond, J.L.; Wanner, M. Diurnal variation in concentrations of various markers of bone metabolism in dogs. Am. J. Vet. Res. 1999, 60, 949–953. [Google Scholar]
- López, I.; Aguilera-Tejero, E.; Estepa, J.C.; Bas, S.; Mayer-Valor, R.; Jiménez, A.; Rodríguez, M. Diurnal variations in the plasma concentration of parathyroid hormone in dogs. Vet. Rec. 2005, 157, 344–347. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.C.; Allen, M.J.; Breur, G.J.; Hoffmann, W.E.; Richardson, D.C. A comparison of two techniques for the determination of serum bone-specific alkaline phosphatase activity in dogs. Res. Vet. Sci. 2000, 68, 231–235. [Google Scholar] [CrossRef]
- Menkes, A.; Mazel, S.; Redmond, R.A.; Koffler, K.; Libanati, C.R.; Gundberg, C.M.; Zizic, T.M.; Hagberg, J.M.; Pratley, R.E.; Hurley, B.F. Strength training increases regional bone mineral density and bone remodeling in middle-aged and older men. J. Appl. Physiol. 1993, 74, 2478–2484. [Google Scholar] [CrossRef]
- Vrbanac, Z.; Brkljaca Bottegaro, N.; Skrlin, B.; Bojanic, K.; Kusec, V.; Stanin, D.; Belic, M. The Effect of a Moderate Exercise Program on Serum Markers of Bone Metabolism in Dogs. Animals 2020, 10, 1481. [Google Scholar] [CrossRef]
- Belic, M.; Kusec, V.; Svetina, A.; Grizelj, J.; Robic, M.; Vrbanac, Z.; Benic, M.; Turk, R. The influence of sex on biochemical markers of bone turnover in dogs. Res. Vet. Sci. 2012, 93, 918–920. [Google Scholar] [CrossRef]
- Lucas, P.W.; Fan, T.M.; Garrett, L.D.; Griffon, D.J.; Wypij, J.M. A comparison of five different bone resorption markers in osteosarcoma-bearing dogs, normal dogs, and dogs with orthopedic diseases. J. Vet. Intern. Med. 2008, 22, 1008–1013. [Google Scholar] [CrossRef] [PubMed]
- Osborne, C.A.; Lulich, J.P.; Polzin, D.J.; Sanderson, S.L.; Koehler, L.A.; Ulrich, L.K.; Bird, K.A.; Swanson, L.L.; Pederson, L.A.; Sudo, S.Z. Analysis of 77,000 canine uroliths: Perspectives from the Minnesota Urolith Center. Vet. Clin. Small Anim. Pract. 1999, 29, 17–38. [Google Scholar] [CrossRef]
- Luskin, A.C.; Lulich, J.P.; Gresch, S.C.; Furrow, E. Bone resorption in dogs with calcium oxalate urolithiasis and idiopathic hypercalciuria. Res. Vet. Sci. 2019, 123, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Groth, E.M.; Lulich, J.P.; Chew, D.J.; Parker, V.J.; Furrow, E. Vitamin D metabolism in dogs with and without hypercalciuric calcium oxalate urolithiasis. J. Vet. Intern. Med. 2019, 33, 758–763. [Google Scholar] [CrossRef] [PubMed]
- Allaway, D.; Gilham, M.; Wagner-Golbs, A.; Maldonado, S.G.; Haydock, R.; Colyer, A.; Stockman, J.; Watson, P. Metabolomic profiling to identify effects of dietary calcium reveal the influence of the individual and postprandial dynamics on the canine plasma metabolome. J. Nutr. Sci. 2019, 8, e13. [Google Scholar] [CrossRef] [PubMed]
- Nybroe, S.; Astrup, A.; Bjørnvad, C.R. Dietary supplementation with flaxseed mucilage alone or in combination with calcium in dogs: Effects on apparent digestibility of fat and energy and fecal characteristics. Int. J. Obes. 2016, 40, 1884–1890. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, R.; Lorenzen, J.K.; Toubro, S.; Krog-Mikkelsen, I.; Astrup, A. Effect of short-term high dietary calcium intake on 24-h energy expenditure, fat oxidation, and fecal fat excretion. Int. J. Obes. 2005, 29, 292–301. [Google Scholar] [CrossRef]
- Papakonstantinou, E.; Flatt, W.P.; Huth, P.J.; Harris, R.B. High dietary calcium reduces body fat content, digestibility of fat, and serum vitamin D in rats. Obes. Res. 2003, 11, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Appleton, G.V.N.; Owen, R.W.; Williamson, R.C.N. The effect of dietary calcium supplementation on intestinal lipid metabolism. J. Steroid Biochem. Mol. Biol. 1992, 42, 383–387. [Google Scholar] [CrossRef]
- Wood, R.J.; Zheng, J.J. High dietary calcium intakes reduce zinc absorption and balance in humans. Am. J. Clin. Nutr. 1997, 65, 1803–1809. [Google Scholar] [CrossRef]
- Gutierrez, O.M. Sodium- and phosphorus-based food additives: Persistent but surmountable hurdles in the management of nutrition in chronic kidney disease. Adv. Chronic Kidney Dis. 2013, 20, 150–156. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, D.G.; Elliott, J.; Church, D.B.; McGreevy, P.D.; Thomson, P.C.; Brodbelt, D.C. Chronic kidney disease in dogs in UK veterinary practices: Prevalence, risk factors, and survival. J. Vet. Intern. Med. 2013, 27, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Lulich, J.; Osborne, C.; O’brien, T.; Polzin, D. Feline renal failure: Questions, answers, questions. Compend. Contin. Educ. Pract. Vet. 1992, 14, 127–153. [Google Scholar]
- Alexander, J.; Stockman, J.; Atwal, J.; Butterwick, R.; Colyer, A.; Elliott, D.; Gilham, M.; Morris, P.; Staunton, R.; Renfrew, H.; et al. Effects of the long-term feeding of diets enriched with inorganic phosphorus on the adult feline kidney and phosphorus metabolism. Br. J. Nutr. 2018, 121, 249–269. [Google Scholar] [CrossRef]
- Böswald, L.F.; Kienzle, E.; Dobenecker, B. Observation about phosphorus and protein supply in cats and dogs prior to the diagnosis of chronic kidney disease. J. Anim. Physiol. Anim. Nutr. 2018, 102, 31–36. [Google Scholar] [CrossRef]
- Tsuda, T.; Ide, M.; Iigo, M. Influences of season and of temperature, photoperiod, and subcutaneous melatonin infusion on the glomerular filtration rate of ewes. J. Pineal. Res. 1995, 19, 166–172. [Google Scholar] [CrossRef]
- Masugata, H.; Senda, S.; Inukai, M.; Himoto, T.; Murao, K.; Hosomi, N.; Iwado, Y.; Noma, T.; Kohno, M.; Goda, F. Seasonal Variation in Estimated Glomerular Filtration Rate Based on Serum Creatinine Levels in Hypertensive Patients. Tohoku J. Exp. Med. 2011, 224, 137–142. [Google Scholar] [CrossRef]
- Von Hendy-Willson, V.E.; Pressler, B.M. An overview of glomerular filtration rate testing in dogs and cats. Vet. J. 2011, 188, 156–165. [Google Scholar] [CrossRef]
- Schneider, P.; Pappritz, G.; Muller-Peddinghaus, R.; Bauer, M.; Lehmann, H.; Ueberberg, H.; Trautwein, G. Potassium hydrogen phosphate induced nephropathy in the dog. I. Pathogenesis of tubular atrophy (author’s transl). Vet. Pathol. 1980, 17, 699–719. [Google Scholar] [CrossRef]
- Dobenecker, B.; Reese, S.; Herbst, S. Effects of dietary phosphates from organic and inorganic sources on parameters of phosphorus homeostasis in healthy adult dogs. PLoS ONE 2021, 16, e0246950. [Google Scholar] [CrossRef] [PubMed]
- Dobenecker, B.; Hertel-Böhnke, P.; Webel, A.; Kienzle, E. Renal phosphorus excretion in adult healthy cats after the intake of high phosphorus diets with either calcium monophosphate or sodium monophosphate. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1759–1765. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| - | Control | Test |
|---|---|---|
| Dry matter (g/100 g) | 89.76 | 90.10 |
| Moisture (g/100 g) | 10.24 | 9.90 |
| Protein (g/100 g) | 30.53 | 25.40 |
| Fat (g/100 g) | 12.22 | 12.20 |
| Ash (g/100 g) | 4.73 | 10.30 |
| Total dietary fiber (g/100 g) | 6.10 | 5.41 |
| Calcium (g/4184 kJ (1000 kcal)) | 1.44 | 7.19 |
| Phosphorus (g/4184 kJ (1000 kcal)) | 1.05 | 4.25 |
| Calcium: Phosphorus ratio | 1.38 | 1.69 |
| Sodium (g/4184 kJ (1000 kcal)) | 1.12 | 1.16 |
| Chloride (g/4184 kJ (1000 kcal)) | 2.02 | 2.11 |
| Potassium (g/4184 kJ (1000 kcal)) | 2.28 | 2.56 |
| Magnesium (mg/4184 kJ (1000 kcal)) | 184 | 338 |
| Iron (mg/4184 kJ (1000 kcal)) | 25.70 | 55.23 |
| Copper (mg/4184 kJ (1000 kcal)) | 1.72 | 2.82 |
| Manganese (mg/4184 kJ (1000 kcal)) | 19.75 | 16.06 |
| Zinc (mg/4184 kJ (1000 kcal)) | 43.56 | 55.51 |
| Iodine (mg/4184 kJ (1000 kcal)) | 1.10 | 1.18 |
| Selenium (µg/4184 kJ (1000 kcal)) | 115.49 † | 121.16 |
| Vitamin A (µg/4184 kJ (µg/1000 kcal)) | 1441 | 1454 |
| Vitamin D3 (µg/4184 kJ (µg/1000 kcal)) | 4.24 | 4.40 |
| Vitamin E (mg/4184 kJ (mg/1000 kcal)) | 70 | 108 |
| Thiamin (mg/4184 kJ (1000 kcal)) | 2.26 | 1.86 |
| Riboflavin (mg/4184 kJ (1000 kcal)) | 1.44 | 1.04 |
| Pantothenic acid (mg/4184 kJ (1000 kcal)) | 6.88 | 7.86 |
| Niacin/nicotinic acid (mg/4184 kJ (1000 kcal)) | 13.12 | 13.27 |
| Pyridoxine (mg/4184 kJ (1000 kcal)) | 2.26 | 2.48 |
| Folic acid (µg/4184 kJ (1000 kcal)) | 354 | 405 |
| Biotin (µg/4184 kJ (1000 kcal)) | 276 | 310 |
| Cyanocobalamin (µg/4184 kJ (1000 kcal)) | 15 | 14 |
| Choline (g/4184 kJ (1000 kcal)) | 607 | 648 |
| Base excess (mEq/kg) ‡ | 96 | 443 |
| PME § (kcal/100 g) | 424.47 | 393.90 |
| - | Weeks | Control | Test | p | ||
|---|---|---|---|---|---|---|
| Mean | 95% Cl | Mean | 95% Cl | |||
| Bodyweight (kg) | Baseline | 13.14 | 11.07, 15.20 | 14.00 | 12.15, 15.85 | 0.997 |
| 8 | 13.25 | 11.19, 15.31 | 13.99 | 12.14, 15.84 | 0.999 | |
| 16 | 13.41 | 11.35, 15.48 | 13.75 | 11.90, 15.60 | 1.000 | |
| 24 | 13.53 | 11.46, 15.59 | 14.05 | 12.20, 15.90 | 1.000 | |
| 32 | 13.64 | 11.57, 15.70 | 13.80 | 11.95, 15.65 | 1.000 | |
| 40 | 13.14 | 11.07, 15.20 | 13.78 | 11.93, 15.63 | 1.000 | |
| Energy intake (kJ/kg 0.75) | Baseline | 628 | 532, 724 | 663 | 577, 748 | 0.998 |
| 8 | 608 | 512, 704 | 665 | 579, 751 | 0.924 | |
| 16 | 592 | 496, 688 | 627 | 542, 713 | 0.998 | |
| 24 | 532 | 436, 628 | 593 | 508, 679 | 0.888 | |
| 32 | 515 * | 419, 611 | 539 * | 453, 625 | 1.000 | |
| 40 | 453 * | 358, 549 | 530 * | 444, 616 | 0.680 | |
| Calcium intake (mg/kg 0.75) † | Baseline | 216 | 183, 249 | 228 | 199, 258 | - |
| 8 | 209 | 176, 242 | 1143 | 995, 1290 | - | |
| 16 | 204 | 171, 237 | 1078 | 931, 1225 | - | |
| 24 | 183 | 150, 216 | 1020 | 872, 1167 | - | |
| 32 | 177 | 144, 210 | 927 | 779, 1074 | - | |
| 40 | 156 | 123, 189 | 911 | 763, 1058 | - | |
| Phosphorus intake (mg/kg 0.75) † | Baseline | 158 | 134, 182 | 166 | 145, 188 | - |
| 8 | 153 | 128, 177 | 675 | 588, 762 | - | |
| 16 | 149 | 125, 173 | 637 | 550, 724 | - | |
| 24 | 134 | 110, 158 | 603 | 516, 690 | - | |
| 32 | 129 | 105, 153 | 548 | 461, 635 | - | |
| 40 | 114 | 90, 138 | 538 | 451, 625 | ||
| - | Weeks | Control | Test | p | ||
|---|---|---|---|---|---|---|
| Mean | 95% Cl | Mean | 95% Cl | |||
| Plasma calcium (mmol/L) | Baseline | 2.48 | 2.42, 2.54 | 2.40 | 2.35, 2.45 | 0.098 |
| 8 | 2.48 | 2.42, 2.54 | 2.44 | 2.38, 2.49 | 0.757 | |
| 16 | 2.50 | 2.44, 2.56 | 2.44 | 2.39, 2.49 | 0.292 | |
| 24 | 2.53 * | 2.47, 2.59 | 2.46 * | 2.41, 2.52 | 0.245 | |
| 32 | 2.54 * | 2.48, 2.61 | 2.49 * | 2.44, 2.54 | 0.505 | |
| 40 | 2.54 * | 2.48, 2.60 | 2.47 * | 2.42, 2.53 | 0.236 | |
| Plasma phosphorus (mmol/L) | Baseline | 1.24 | 1.11, 1.39 | 1.30 | 1.18, 1.44 | 0.992 |
| 8 | 1.21 | 1.08, 1.36 | 1.20 | 1.08, 1.33 | 1.000 | |
| 16 | 1.29 | 1.15, 1.45 | 1.19 | 1.08, 1.32 | 0.817 | |
| 24 | 1.22 | 1.08, 1.36 | 1.20 | 1.08, 1.33 | 1.000 | |
| 32 | 1.26 | 1.12, 1.41 | 1.14 * | 1.03, 1.26 | 0.558 | |
| 40 | 1.17 | 1.04, 1.31 | 1.15 * | 1.04, 1.27 | 1.000 | |
| Ionized calcium (mmol/L) | Baseline | 1.29 | 1.25, 1.33 | 1.28 | 1.24, 1.32 | 0.999 |
| 8 | 1.18 * | 1.14, 1.22 | 1.29 | 1.26, 1.33 | <0.001 ** | |
| 16 | 1.30 | 1.26, 1.34 | 1.31 | 1.27, 1.34 | 1.000 | |
| 24 | 1.30 | 1.26, 1.34 | 1.32 | 1.28, 1.36 | 0.989 | |
| 32 | 1.31 | 1.26, 1.35 | 1.30 | 1.27, 1.34 | 1.000 | |
| 40 | 1.28 | 1.24, 1.32 | 1.29 | 1.25, 1.32 | 1.000 | |
| Parathyroid hormone (pg/mL) | Baseline | 18.27 | 11.72, 28.47 | 12.18 | 8.19, 18.11 | 0.488 |
| 8 | 12.38 | 7.94, 19.29 | 11.06 | 7.44, 16.45 | 1.000 | |
| 16 | 12.32 | 7.90, 19.19 | 14.13 | 9.50, 21.01 | 1.000 | |
| 24 | 10.33 | 6.63, 16.09 | 9.99 | 6.72, 14.86 | 1.000 | |
| 32 | 10.58 | 6.79, 16.49 | 9.61 | 6.46, 14.29 | 1.000 | |
| 40 | 15.26 | 9.79, 23.78 | 13.72 | 9.23, 20.41 | 1.000 | |
| Fibroblast growth factor 23 (pg/mL) | Baseline | 336 | 253, 447 | 411 | 319, 530 | 0.808 |
| 8 | 351 | 264, 466 | 392 | 304, 505 | 0.996 | |
| 16 | 311 | 234, 414 | 384 | 298, 496 | 0.759 | |
| 24 | 293 | 220, 390 | 348 | 270, 449 | 0.912 | |
| 32 | 323 | 243, 429 | 339 | 263, 438 | 1.000 | |
| 40 | 178 * | 134, 237 | 209 * | 162, 269 | 0.952 | |
| Total calcidiol (25OHD) (nmol/L) | Baseline | 70.30 | 56.97, 86.75 | 79.10 | 65.54, 95.47 | 0.956 |
| 8 | 73.46 | 59.53, 90.65 | 82.98 | 68.76, 100.15 | 0.946 | |
| 16 | 69.37 | 56.21, 85.60 | 79.06 | 65.51, 95.42 | 0.918 | |
| 24 | 75.28 | 61.01, 92.89 | 80.81 | 66.96, 97.53 | 0.999 | |
| 32 | 82.49 * | 66.85, 101.80 | 86.89 | 71.99, 104.87 | 1.000 | |
| 40 | 78.52 | 63.63, 96.89 | 86.44 | 71.63, 104.33 | 0.990 | |
| Calcitriol (1,25(OH)2D) (pmol/L) | Baseline | 25.10 | 18.25, 34.53 | 25.43 | 19.25, 33.60 | 1.000 |
| 8 | 17.49 * | 12.55, 24.38 | 30.40 | 23.01, 40.17 | 0.008 ** | |
| 16 | 16.80 * | 12.05, 23.41 | 27.61 | 20.90, 36.48 | 0.027 ** | |
| 24 | 21.01 | 15.23, 29.00 | 26.72 | 20.22, 35.31 | 0.757 | |
| 32 | 17.96 * | 13.02, 24.79 | 23.38 | 17.61, 31.05 | 0.660 | |
| 40 | 17.80 * | 12.89, 24.56 | 25.17 | 19.05, 33.25 | 0.284 | |
| 24,25-dihydroxycholecalciferol (24,25(OH)2D3) (nmol/L) | Baseline | 31.12 | 23.08, 41.98 | 35.05 | 26.82, 45.80 | 0.997 |
| 8 | 30.84 | 22.87, 41.59 | 33.76 | 25.83, 44.11 | 1.000 | |
| 16 | 25.56 * | 18.95, 34.47 | 31.23 | 23.90, 40.81 | 0.884 | |
| 24 | 23.21 * | 17.21, 31.30 | 26.88 * | 20.57, 35.13 | 0.984 | |
| 32 | 23.48 * | 17.41, 31.66 | 24.22 * | 18.54, 31.65 | 1.000 | |
| 40 | 23.23 * | 17.22, 31.33 | 25.65 * | 19.63, 33.51 | 0.999 | |
| 24,25-dihydroxyergocalciferol (24,25(OH)D2) (nmol/L) | Baseline | 9.19 | 6.33, 13.33 | 7.06 | 5.06, 9.85 | 0.857 |
| 8 | 9.27 | 6.39, 13.45 | 7.89 | 5.66, 11.01 | 0.995 | |
| 16 | 7.57 * | 5.21, 10.98 | 6.74 | 4.83, 9.40 | 1.000 | |
| 24 | 6.77 * | 4.67, 9.83 | 5.97 * | 4.28, 8.32 | 0.999 | |
| 32 | 6.25 * | 4.31, 9.07 | 5.72 * | 4.10, 7.98 | 1.000 | |
| 40 | 6.79 * | 4.68, 9.85 | 5.69 * | 4.08, 7.94 | 0.989 | |
| Total dihydroxycalcidiol (24,25(OH)2D) (nmol/L) | Baseline | 40.83 | 31.33, 53.21 | 42.72 | 33.71, 54.14 | 1.000 |
| 8 | 40.69 | 31.22, 53.03 | 42.53 | 33.56, 53.90 | 1.000 | |
| 16 | 33.58 * | 25.77, 43.77 | 38.57 | 30.44, 48.88 | 0.974 | |
| 24 | 30.51 * | 23.41, 39.77 | 33.45 * | 26.39, 42.39 | 0.999 | |
| 32 | 30.11 * | 23.10, 39.24 | 30.60 * | 24.14, 38.77 | 1.000 | |
| 40 | 30.33 * | 23.27, 39.53 | 31.95 * | 25.21, 40.48 | 1.000 | |
| Calcidiol (25OHD3) (nmol/L) | Baseline | 68.97 | 55.79, 85.26 | 77.59 | 64.19, 93.79 | 0.959 |
| 8 | 72.41 | 58.58, 89.52 | 81.86 | 67.72, 98.95 | 0.947 | |
| 16 | 68.24 | 55.20, 84.36 | 78.10 | 64.60, 94.40 | 0.907 | |
| 24 | 73.12 | 59.15, 90.39 | 79.40 | 65.68, 95.98 | 0.997 | |
| 32 | 78.99 | 63.90, 97.65 | 83.44 | 69.03, 100.87 | 1.000 | |
| 40 | 73.25 | 59.26, 90.56 | 83.73 | 69.27, 101.22 | 0.912 | |
| Ergocalcidiol (25OHD2) (nmol/L) | Baseline | 1.26 | 0.79, 1.99 | 1.47 | 0.97, 2.22 | 0.999 |
| 8 | 1.03 | 0.65, 1.64 | 1.09 | 0.72, 1.64 | 1.000 | |
| 16 | 1.09 | 0.69, 1.72 | 0.92 | 0.61, 1.40 | 0.998 | |
| 24 | 1.85 | 1.17, 2.93 | 1.27 | 0.84, 1.92 | 0.649 | |
| 32 | 2.88 * | 1.81, 4.56 | 2.65 * | 1.75, 4.00 | 1.000 | |
| 40 | 4.34 * | 2.74, 6.89 | 2.51 * | 1.66, 3.80 | 0.165 | |
| Total calcidiol: total dihydroxycalcidiol ratio (25OHD:24,25(OH)2D) | Baseline | 1.72 | 1.49, 1.98 | 1.85 | 1.63, 2.10 | 0.970 |
| 8 | 1.81 | 1.57, 2.08 | 1.95 | 1.72, 2.21 | 0.954 | |
| 16 | 2.07 * | 1.79, 2.38 | 2.05 | 1.81, 2.33 | 1.000 | |
| 24 | 2.47 * | 2.14, 2.84 | 2.42 * | 2.13, 2.74 | 1.000 | |
| 32 | 2.74 * | 2.38, 3.16 | 2.84 * | 2.50, 3.22 | 1.000 | |
| 40 | 2.59 * | 2.25, 2.98 | 2.71 * | 2.38, 3.07 | 0.999 | |
| - | Weeks | Control | Test | p | ||
|---|---|---|---|---|---|---|
| Mean | 95% Cl | Mean | 95% Cl | |||
| Plasma creatinine (µmol/L) | Baseline | 78.67 | 71.35, 86.76 | 85.43 | 78.27, 93.23 | 0.674 |
| 8 | 78.00 | 70.73, 86.01 | 85.46 | 78.30, 93.27 | 0.541 ** | |
| 16 | 81.15 | 73.59, 89.49 | 81.13 * | 74.34, 88.55 | 1.000 | |
| 24 | 87.62 * | 79.45, 96.62 | 91.99 * | 84.29, 100.40 | 0.982 | |
| 32 | 88.25 * | 80.03, 97.31 | 91.68 * | 84.01, 100.06 | 0.997 | |
| 40 | 89.73 * | 81.37, 98.94 | 94.69 * | 86.76, 103.35 | 0.964 | |
| Plasma urea (mmol/L) | Baseline | 5.20 | 4.52, 5.98 | 6.41 | 5.66, 7.27 | 0.028 |
| 8 | 5.54 | 4.81, 6.37 | 5.14 * | 4.54, 5.83 | 0.965 ** | |
| 16 | 6.05 * | 5.26, 6.97 | 5.19 * | 4.57, 5.88 | 0.253 ** | |
| 24 | 5.69 | 4.94, 6.55 | 5.54 * | 4.89, 6.28 | 1.000 ** | |
| 32 | 6.16 * | 5.35, 7.08 | 5.26 * | 4.64, 5.96 | 0.235 ** | |
| 40 | 5.45 | 4.74, 6.27 | 5.26 * | 4.64, 5.96 | 1.000 ** | |
| Urine specific gravity | Baseline | 1.034 | 1.034, 1.035 | 1.034 | 1.034, 1.035 | 0.841 |
| 8 | 1.034 | 1.034, 1.035 | 1.034 | 1.034, 1.035 | 1.000 | |
| 16 | 1.035 | 1.034, 1.035 | 1.035 | 1.034, 1.035 | 1.000 | |
| 24 | 1.034 | 1.034, 1.035 | 1.035 | 1.034, 1.035 | 0.794 | |
| 32 | 1.035 * | 1.034, 1.035 | 1.035 | 1.034, 1.035 | 1.000 | |
| 40 | 1.034 | 1.034, 1.035 | 1.035 | 1.034, 1.035 | 0.429 | |
| Urine RSS struvite | Baseline | 0.26 | 0.07, 0.93 | 1.41 | 0.46, 4.35 | 0.075 |
| 8 | 0.75 | 0.21, 2.66 | 0.05 * | 0.02, 0.16 | <0.001 ** | |
| 16 | 0.70 | 0.20, 2.48 | 0.31 * | 0.10, 0.97 | 0.888 ** | |
| 24 | 1.01 | 0.28, 3.55 | 0.41 | 0.13, 1.28 | 0.811 ** | |
| 32 | 1.10 | 0.31, 3.89 | 0.47 | 0.15, 1.45 | 0.846 ** | |
| 40 | 2.42 * | 0.68, 8.53 | 0.75 | 0.24, 2.32 | 0.477 ** | |
| Urine RSS calcium oxalate | Baseline | 2.18 | 1.31, 3.65 | 5.87 | 3.71, 9.30 | 0.001 |
| 8 | 3.39 | 2.02, 5.66 | 3.07 * | 1.94, 4.86 | 1.000 ** | |
| 16 | 3.50 | 2.09, 5.85 | 5.70 | 3.60, 9.02 | 0.452 | |
| 24 | 3.84 | 2.30, 6.42 | 4.83 | 3.05, 7.65 | 0.989 | |
| 32 | 2.68 | 1.60, 4.47 | 4.76 | 3.01, 7.54 | 0.228 | |
| 40 | 3.25 | 1.94, 5.44 | 5.11 | 3.22, 8.09 | 0.558 | |
| Urine albumin: creatinine ratio (mg/g) | Baseline | 12.16 | 4.47, 33.07 | 5.50 | 2.25, 13.45 | 0.738 |
| 8 | 8.81 | 3.24, 23.96 | 8.46 | 3.46, 20.71 | 1.000 | |
| 16 | 6.49 * | 2.38, 17.64 | 13.06 * | 5.34, 31.97 | 0.851 ** | |
| 24 | 6.29 * | 2.31, 17.10 | 8.67 | 3.54, 21.23 | 0.999 ** | |
| 32 | 8.44 | 3.10, 22.96 | 7.73 | 3.16, 18.92 | 1.000 | |
| 40 | 5.56 * | 2.04, 15.11 | 8.27 | 3.38, 20.25 | 0.997 ** | |
| Weight adjusted clearance (ml/min/kg) | Baseline | 3.04 | 2.40, 3.68 | 3.03 | 2.46, 3.60 | 1.000 |
| 8 | 3.55 | 2.91, 4.19 | 2.90 | 2.32, 3.47 | 0.335 | |
| 16 | 2.93 | 2.29, 3.57 | 2.82 | 2.25, 3.39 | 1.000 | |
| 24 | 2.68 | 2.04, 3.32 | 2.35 * | 1.78, 2.92 | 0.971 | |
| 32 | 2.53 | 1.89, 3.17 | 2.39 | 1.82, 2.96 | 1.000 | |
| 40 | 2.40 | 1.76, 3.04 | 2.29 * | 1.71, 2.86 | 1.000 | |
| - | Weeks | Control | Test | p | ||
|---|---|---|---|---|---|---|
| Mean | 95% Cl | Mean | 95% Cl | |||
| DXA: Bone mineral content (g) | Baseline | 393 | 331, 467 | 422 | 362, 492 | 0.961 |
| 24 | 404 * | 340, 479 | 429 | 368, 501 | 0.979 | |
| 40 | 403 | 340, 478 | 429 | 368, 500 | 0.978 | |
| DXA: Bone mineral density (g/cm2) | Baseline | 0.820 | 0.787, 0.855 | 0.826 | 0.796, 0.857 | 0.999 |
| 24 | 0.822 | 0.788, 0.856 | 0.829 | 0.799, 0.860 | 0.997 | |
| 40 | 0.819 | 0.785, 0.853 | 0.831 | 0.801, 0.862 | 0.972 | |
| Bone alkaline phosphatase (U/L) | Baseline | 14.56 | 10.69, 19.85 | 14.90 | 11.30, 19.66 | 1.000 |
| 8 | 11.49 | 8.43, 15.66 | 16.56 | 12.56, 21.84 | 0.179 | |
| 16 | 10.17 * | 7.46, 13.85 | 15.63 | 11.85, 20.62 | 0.058 | |
| 24 | 10.54 * | 7.74, 14.37 | 14.09 | 10.68, 18.58 | 0.477 | |
| 32 | 9.33 * | 6.85, 12.71 | 11.00 * | 8.34, 14.51 | 0.962 | |
| 40 | 9.59 * | 7.04, 13.06 | 10.99 * | 8.34, 14.50 | 0.990 | |
| Serum cross-laps (ng/mL) | Baseline | 0.40 | 0.22, 0.71 | 0.47 | 0.28, 0.79 | 1.000 |
| 8 | 0.27 * | 0.15, 0.49 | 0.43 | 0.26, 0.72 | 0.730 | |
| 16 | 0.27 * | 0.15, 0.48 | 0.34 * | 0.20, 0.57 | 0.997 | |
| 24 | 0.28 | 0.16, 0.50 | 0.32 * | 0.19, 0.54 | 1.000 | |
| 32 | 0.25 * | 0.14, 0.44 | 0.33 * | 0.20, 0.56 | 0.976 | |
| 40 | 0.25 * | 0.14, 0.44 | 0.26 * | 0.15, 0.43 | 1.000 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atwal, J.; Stockman, J.; Gilham, M.; Allaway, D.; Renfrew, H.; Bakke, A.M.; Watson, P. No Observed Adverse Effects on Health Were Detected in Adult Beagle Dogs When Fed a High-Calcium Diet for 40 Weeks. Animals 2021, 11, 1799. https://doi.org/10.3390/ani11061799
Atwal J, Stockman J, Gilham M, Allaway D, Renfrew H, Bakke AM, Watson P. No Observed Adverse Effects on Health Were Detected in Adult Beagle Dogs When Fed a High-Calcium Diet for 40 Weeks. Animals. 2021; 11(6):1799. https://doi.org/10.3390/ani11061799
Chicago/Turabian StyleAtwal, Jujhar, Jonathan Stockman, Matthew Gilham, David Allaway, Helen Renfrew, Anne Marie Bakke, and Phillip Watson. 2021. "No Observed Adverse Effects on Health Were Detected in Adult Beagle Dogs When Fed a High-Calcium Diet for 40 Weeks" Animals 11, no. 6: 1799. https://doi.org/10.3390/ani11061799
APA StyleAtwal, J., Stockman, J., Gilham, M., Allaway, D., Renfrew, H., Bakke, A. M., & Watson, P. (2021). No Observed Adverse Effects on Health Were Detected in Adult Beagle Dogs When Fed a High-Calcium Diet for 40 Weeks. Animals, 11(6), 1799. https://doi.org/10.3390/ani11061799