
What Goes Wrong during Early Development of Artificially Reproduced European Eel Anguilla anguilla? Clues from the Larval Transcriptome and Gene Expression Patterns


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Broodstock
2.2. Induction of Gametogenesis
2.3. Fertilization and Egg Rearing
2.4. Larvae Collection
2.5. RNA-Sequencing
2.6. Gene Expression
2.6.1. Gene Description and Primer Designs
2.6.2. RNA Isolation
2.6.3. Quantitative RT-PCR
2.6.4. Statistical Analysis
3. Results
3.1. Eel Larvae Transcriptomics
3.2. Eel Larvae Temporal Expression
4. Discussion
4.1. Immune Response
4.2. Osmoregulation
4.3. Myogenesis, Neurogenesis, and Sensory Development
4.4. Digestive Function and Hyaluronan Metabolism
4.5. Activity: Movement and Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fontaine, M. Sur la maturation complète des organes génitaux de l’anguille male et l’emission spontanèe de ses produits sexuels. Comptes Rendus Acad. Sci. 1936, 202, 1312–1315. [Google Scholar]
- Fontaine, M.; Bertrand, E.; Lopez, E.; Callamand, O. Sur la maturation des organes génitaux de l’anguille femelle (Anguilla anguilla L.) et l’émission spontanée des œufs en aquarium. Comptes Rendus Acad. Sci. 1964, 259, 2907–2910. [Google Scholar]
- Boëtius, I.; Boëtius, J. Experimental maturation of female silver eels, Anguilla anguilla. Estimates of fecundity and energy reserves for migration and spawning. Dana 1980, 1, 1–28. [Google Scholar]
- Bezdenezhnykh, V.A.; Prokhorchik, G.A.; Petrikov, A.M.; Petukov, V.B.; Plyuta, M.V. Obtaining the larvae of European eel Anguilla anguilla L. (Pisces, Anguillidae) under experimental conditions. Dokl. Akad. Nauk SSSR 1983, 5, 1264–1266. [Google Scholar]
- Mordenti, O.; Di Biase, A.; Bastone, G.; Sirri, R.; Zaccaroni, A.; Parmeggiani, A. Controlled reproduction in the wild European eel (Anguilla anguilla); two populations compared. Aquac. Int. 2013, 21, 1045–1063. [Google Scholar] [CrossRef]
- Sørensen, S.R.; Tomkiewicz, J.; Munk, P.; Butts, I.A.E.; Nielsen, A.; Lauesen, P.; Graver, C. Ontogeny and growth of early life stages of captive-bred European eel. Aquaculture 2016, 456, 50–61. [Google Scholar] [CrossRef]
- Asturiano, J.F.; Sørensen, S.R.; Perez, L.; Lauesen, P.; Tomkiewicz, J. First production of larvae using cryopreserved sperm: Effects of preservation temperature and cryopreservation on European eel sperm fertilization capacity. Reprod. Domest. Anim. 2016, 51, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Jéhannet, P.; Heinsbroek, L.T.N.; Palstra, A.P. Ultrasonography to assist with timing of spawning in European eel. Theriogenology 2017, 101, 73–80. [Google Scholar] [CrossRef]
- Sørensen, S.R.; Skov, P.V.; Lauesen, P.; Tomkiewicz, J.; Bossier, P.; De Schryver, P. Microbial interference and potential control in culture of European eel (Anguilla anguilla) embryos and larvae. Aquaculture 2014, 426–427, 1–8. [Google Scholar] [CrossRef]
- Da Silva, F.F.G.; Jacobsen, C.; Kjørsvik, E.; Støttrup, J.G.; Tomkiewics, J. Oocyte and egg quality indicators in European eel: Lipid droplet coalescence and fatty acid composition. Aquaculture 2018, 496, 30–38. [Google Scholar] [CrossRef]
- Masuda, Y.; Imaizumi, H.; Oda, K.; Hashimoto, H.; Usuki, H.; Teruya, K. Artificial completion of the Japanese eel, Anguilla japonica, life cycle: Challenge to mass production. Bull. Fish. Res. Agency 2012, 35, 111–117. [Google Scholar]
- Okamura, A.; Horie, N.; Yamada, Y.; Mikawa, N.; Tsukamoto, K. Obtaining high-quality larvae for mass production of glass eels: Can we refine our approach? Aquaculture 2020, 521, 735049. [Google Scholar] [CrossRef]
- Vadstein, O.; Bergh, O.; Gatesoupe, F.G.; Galindo-Villegas, J.; Mulero, V.; Picchietti, S.; Makridis, P.; Olsen, Y.; Dierckens, K.; Defoirdt, T.; et al. Microbiology and immunology of fish larvae. Rev. Aquac. 2013, 5, S1–S25. [Google Scholar] [CrossRef]
- Vadstein, O.; Attramdal, K.J.K.; Bakke, I.; Olsen, Y. K-selection as microbial community management strategy: A method for improved viability of larvae in aquaculture. Front. Microbiol. 2018, 9, 2730. [Google Scholar] [CrossRef] [PubMed]
- Miest, J.J.; Politis, S.N.; Adamek, M.; Tomkiewicz, J.; Butts, I.A.E. Molecular ontogeny of larval immunity in European eel at increasing temperatures. Fish Shellfish Immun. 2019, 87, 105–119. [Google Scholar] [CrossRef]
- Varsamos, S.; Nebel, C.; Charmantier, G. Ontogeny of osmoregulation in postembryonic fish: A review. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2005, 141, 401–429. [Google Scholar] [CrossRef] [PubMed]
- Sasai, S.; Kaneko, T.; Tsukamoto, K. Extrabranchial chloride cells in early life stages of the Japanese eel, Anguilla japonica. Ichthyol. Res. 1998, 45, 95–98. [Google Scholar] [CrossRef]
- Ahn, H.; Lee, K.M.; Inokuchi, M.; Watanabe, S.; Okamura, A.; Tsukamoto, K.; Kaneko, T. Observations of initial water ingestion and ion absorption in the digestive tract of Japanese eel larvae. Fish Sci. 2015, 81, 283–290. [Google Scholar] [CrossRef]
- Bagarino, T. Yolk resorption, onset of feeding and survival potential of larvae of three tropical marine fish species reared in the hatchery. Mar. Biol. 1986, 91, 449–459. [Google Scholar] [CrossRef]
- Lasker, R. Efficiency and rate of yolk utilization by developing embryos and larvae of the Pacific sardine, Sardinops caerulea (Girard). J. Fish. Res. Board Can. 1962, 19, 867–875. [Google Scholar] [CrossRef]
- Avila, E.M.; Juario, J.V. Yolk and oil globule utilization and developmental morphology of the digestive tract epithelium in larval rabbitfish, Siganus guttatus (Bloch). Aquaculture 1987, 65, 319–331. [Google Scholar] [CrossRef]
- Xia, J.H.; Liu, J.X.; Zhou, L.; LI, Z.; Gui, J.F. Apo-14 is required for digestive system organogenesis during fish embryogenesis and larval development. Int. J. Dev. Biol. 2008, 52, 1089–1098. [Google Scholar] [CrossRef]
- Mazurais, D.; Darias, M.; Zambonino-Infante, J.L.; Cahu, C.L. Transcriptomics for understanding marine fish larval development. Can. J. Zool. 2011, 89, 599–611. [Google Scholar] [CrossRef]
- Yamauchi, K.; Nakamura, M.; Takahashi, H.; Takano, K. Cultivation of larvae of Japanese eel. Nature 1976, 263, 412. [Google Scholar] [CrossRef]
- Lokman, P.M.; Young, G. Induced spawning and early ontogeny of New Zealand freshwater eels (Anguilla dieffenbachii and A. australis). N. Z. J. Mar. Freshw. Res. 2000, 34, 135–145. [Google Scholar] [CrossRef]
- Zambonino-Infante, J.L.; Gisbert, E.; Sarasquete, S.; Navarro, I.; Gutierrez, J.; Cahu, C.L. Ontogeny and physiology of the digestive system of marine fish larvae. In Feeding and Digestive Functions in Fishes; Cyrino, J.E.P., Bureau, D., Kapoor, B.D., Eds.; Science Publishers: Enfield, NH, USA, 2008; pp. 281–348. [Google Scholar]
- Kurokawa, T.; Suzuki, T.; Ohta, H.; Kagawa, H.; Tanaka, H.; Unuma, T. Expression of pancreatic enzyme genes during the early larval stage of Japanese eel Anguilla japonica. Fish. Res. 2002, 68, 736–744. [Google Scholar] [CrossRef]
- Mazurais, D.; Kjørsvik, E.; World, P.A.; Politis, S.N.; Cahu, C.; Tomkiewicz, J.; Zambonino-Infante, J. Biochemical, histological and molecular study of digestive tract development in European eel larvae (Anguilla anguilla) prior to exogenous feeding. In Proceedings of the Aquaculture Europe 13, Trondheim, Norway, 9–12 August 2013. [Google Scholar]
- Bouillart, M.; Tomkiewicz, J.; Lauesen, P.; De Kegel, B.; Adriaens, D. Musculoskeletal anatomy and feeding performance of pre-feeding engyodontic larvae of the European eel (Anguila anguilla). J. Anat. 2015, 227, 325–340. [Google Scholar] [CrossRef]
- Masuda, Y.; Shima, Y.; Tamaru, O.; Takahashi, Y.; Ohmura, Y.; Takashi, I.; Kamoshida, M.; Arimoto, M.; Yamano, K.; Yatabe, T. Japanese eel jaw and vertebra ossification occurring respectively during the larval stage and metamorphosis. Fish. Sci. 2019, 85, 1045–1054. [Google Scholar] [CrossRef]
- Politis, S.N.; Sørensen, S.R.; Mazurais, D.; Servili, A.; Zambonino-Infante, J.; Miest, J.J.; Clemmesen, C.M.; Tomkiewicz, J.; Butts, I.A.E. Molecular ontogeny of first-feeding European eel larvae. Front. Physiol. 2018, 9, 1477. [Google Scholar] [CrossRef]
- Parmeggiani, A.; Zannoni, A.; Tubon, I.; Casalini, A.; Emmanuele, P.; Forni, M.; Mordenti, O. Initial ontogeny of digestive enzymes in the early life stages of captive-bred European eels during fasting: A partial characterization. Res. Vet. Sci. 2020, 132, 54–56. [Google Scholar] [CrossRef]
- Bishop, R.E.; Torres, J.J. Leptocephalus energetics: Metabolism and excretion. J. Exp. Biol. 1999, 202, 2485–2493. [Google Scholar] [CrossRef]
- Pfeiler, E.; Govoni, J. Metabolic rates in early life history stages of Elopomorph fishes. Biol. Bull. 1993, 185, 277–283. [Google Scholar] [CrossRef]
- Politis, S.N.; Butts, I.A.E.; Tomkiewicz, J. Light impacts embryonic and early larval development of the European eel, Anguilla anguilla. J. Exp. Mar. Biol. Ecol. 2014, 461, 407–415. [Google Scholar] [CrossRef]
- Politis, S.N.; Mazurais, D.; Servili, A.; Zambonino-Infante, J.L.; Miest, J.J.; Sorensen, S.R.; Tomkiewicz, J.; Butts, I.A.E. Temperature effects on gene expression and morphological development of European eel, Anguilla anguilla larvae. PLoS ONE 2017, 12, e0182726. [Google Scholar] [CrossRef] [PubMed]
- Furuita, H.; Ohta, H.; Unuma, T.; Tanaka, H.; Kagawa, H.; Suzuki, N.; Yamamoto, T. Biochemical composition of eggs in relation to egg quality in the Japanese eel, Anguilla japonica. Fish Physiol. Biochem. 2003, 29, 37–46. [Google Scholar] [CrossRef]
- Furuita, H.; Unuma, T.; Nomura, K.; Tanaka, H.; Okuzawa, K.; Sugita, T.; Yamamoto, T. Lipid and fatty acid composition of eggs producing larvae with high survival rate in the Japanese eel. J. Fish Biol. 2006, 69, 1178–1189. [Google Scholar] [CrossRef]
- Kurokawa, T.; Okamoto, T.; Gen, K.; Uji, S.; Murashita, K.; Unuma, T.; Nomura, K.; Matsuraba, H.; Kim, S.K.; Ohta, H.; et al. Influence of water temperature on morphological deformities in cultured larvae of Japanese eel, Anguilla japonica, at completion of yolk resorption. J. World Aquac. Soc. 2008, 39, 726–735. [Google Scholar] [CrossRef]
- Okamura, A.; Yamada, Y.; Horie, N.; Utoh, T.; Mikawa, N.; Tanaka, S.; Tsukamoto, K. Effects of water temperature on early development of Japanese eel Anguilla japonica. Fish. Sci. 2007, 73, 1241–1248. [Google Scholar]
- Okamura, A.; Yamada, Y.; Mikawa, N.; Horie, N.; Utoh, T.; Kaneko, T.; Tanaka, S.; Tsukamoto, K. Growth and survival of eel leptocephali (Anguilla japonica) in low-salinity water. Aquaculture 2009, 296, 367–372. [Google Scholar] [CrossRef]
- Hsu, H.Y.; Chen, S.H.; Cha, Y.R.; Tsukamoto, K.; Lin, C.Y.; Han, Y.S. De Novo Assembly of the Whole Transcriptome of the Wild Embryo, Preleptocephalus, Leptocephalus, and Glass Eel of Anguilla japonica and Deciphering the Digestive and Absorptive Capacities during Early Development. PLoS ONE 2015, 10, e0139105. [Google Scholar] [CrossRef]
- Izumi, H.; Gen, K.; Lokman, P.M.; Hagihara, S.; Horiuchi, M.; Tanaka, T.; Ijiri, S.; Adachi, S. Maternal transcripts in good and poor quality eggs from Japanese eel, Anguilla japonica, their identification by large-scale quantitative analysis. Mol. Reprod. Dev. 2019, 86, 1846–1864. [Google Scholar] [CrossRef]
- Chai, Y.; Tosaka, R.; Sago, K.; Hatanaka, R.; Ijiri, S.; Adachi, S. The relationship between the developmental stage of oocytes in various seasons and the quality of the egg obtained by artificial maturation in the feminized Japanese eel Anguilla Japonica. Aquac. Sci. 2010, 58, 269–278. [Google Scholar]
- Mes, D.; Dirks, R.P.; Palstra, A.P. Simulated migration under mimicked photothermal conditions enhances sexual maturation of farmed European eel (Anguilla anguilla). Aquaculture 2016, 452, 367–372. [Google Scholar] [CrossRef]
- Lokman, P.M.; Wylie, M.J.; Downes, M.; Di Biase, A.; Damsteegt, E. Artificial induction of maturation in female eels, Anguilla australis: The benefits of androgen pre-treatment. Aquaculture 2015, 437, 111–119. [Google Scholar] [CrossRef]
- Thomson-Laing, G.; Damsteegt, E.L.; Nagata, J.; Ijiri, S.; Adachi, S.; Todo, T.; Hiramatsu, N.; Lokman, P.M. Synergistic effects of estradiol and 11-ketotestosterone on vitellogenin physiology in the shortfinned eel (Anguilla australis). Biol. Reprod. 2019, 100, 1319–1332. [Google Scholar] [CrossRef]
- Palstra, A.P.; Cohen, E.G.H.; Niemantsverdriet, P.R.W.; van Ginneken, V.J.T.; van den Thillart, G.E.E.J.M. Artificial maturation and reproduction of European silver eel: Development of oocytes during final maturation. Aquaculture 2005, 249, 533–547. [Google Scholar] [CrossRef]
- Ohta, H.; Kagawa, H.; Tanaka, H.; Okuzawa, K.; Iinuma, N.; Hirose, K. Artificial induction of maturation and fertilization in the Japanese eel, Anguilla Japonica. Aquaculture 1996, 139, 291–301. [Google Scholar] [CrossRef]
- Kahn, I.A.; Lopez, E.; Leloup-Hatey, J. Induction of spermatogenesis and spermiation by a single injection of human chorionic gonadotropin in intact and hypophysectomised immature European eel (Anguilla anguilla L.). Gen. Comp. Endocrinol. 1987, 68, 91–103. [Google Scholar] [CrossRef]
- Pérez, L.; Asturiano, J.F.; Tomás, A.; Zegrari, S.; Barrera, R.; Espinós, F.J.; Jover, M. Induction of maturation and spermiation in the male European eel: Assessment of sperm quality throughout treatment. J. Fish Biol. 2005, 57, 1488–1504. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N. 1000 Genome project data processing subgroup. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq-a Python framework to work with high-throughput sequencing data. Bioinformatics 2014, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Jéhannet, P.; Kruijt, L.; Damsteegt, E.L.; Swinkels, W.; Heinsbroek, L.T.N.; Lokman, P.M.; Palstra, A.P. A mechanistic model for studying the initiation of anguillid vitellogenesis by comparing the European eel (Anguilla anguilla) and the shortfinned eel (A. australis). Gen. Comp. Endocrinol. 2019, 279, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Setiawan, A.N.; Lokman, P.M. The use of reference gene selection programs to study the silvering transformation in a freshwater eel Anguilla australis: A cautionary tale. BMC Mol. Biol. 2010, 11, 75. [Google Scholar] [CrossRef]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3–new capabilities and interfaces. Nucl. Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Henkel, C.V.; Burgerhout, E.; de Wijze, D.L.; Dirks, R.P.; Minegishi, Y.; Jansen, H.J.; Spaink, H.P.; Dufour, S.; Weltzien, F.-A.; Tsukamoto, K.; et al. Primitive duplicate hox clusters in the European eel’s genome. PLoS ONE 2012, 7, e32231. [Google Scholar] [CrossRef]
- Jansen, H.J.; Liem, M.; Jong-Raadsen, S.A.; Dufour, S.; Weltzien, F.A.; Swinkels, W.; Koelewijn, A.; Palstra, A.P.; Pelster, B.; Spaink, H.P. Rapid de novo assembly of the European eel genome from nanopore sequencing reads. Sci. Rep. 2017, 7, 7213. [Google Scholar] [CrossRef]
- Bustin, A.S.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Satoh, K.; Yamada, H.; Takebe, T.; Nikaido, H.; Shiozawa, S. Assessment of the nutritional status of field-caught larval Pacific bluefin tuna by RNA/DNA ratio based on a starvation experiment of hatchery-reared fish. J. Exp. Mar. Biol. Ecol. 2008, 4, 56–64. [Google Scholar] [CrossRef]
- Bobe, J.; Labbé, C. Egg and sperm quality in fish. Gen. Comp. Endocrinol. 2010, 165, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liu, Y.; Luo, Y.; Zhong, H.; Huang, T.; Liang, W.; Xiao, J.; Wu, W.; Li, L.; Chen, M. Large-scale profiling of the proteome and dual transcriptome in Nile tilapia (Oreochromis niloticus) challenged with low- and high-virulence strains of Streptococcus agalactiae. Fish Shellfish Immunol. 2020, 100, 386–396. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, J.; Xu, X.; Fu, J.; Li, J. Expression of complement component C7 and involvement in innate immune responses to bacteria in grass carp. Fish Shellfish Immunol. 2012, 33, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Gao, Y.; Shu, C.; Xu, T. Characterization and evolutionary analysis of duplicated C7 in miiuy croaker. Fish Shellfish Immunol. 2015, 45, 672–679. [Google Scholar] [CrossRef]
- Guo, B.; Wu, C.; Lv, Z.; Liu, C. Characterization and expression analysis of two terminal complement components: C7 and C9 from large yellow croaker, Larimichthys crocea. Fish Shellfish Immunol. 2016, 51, 211–219. [Google Scholar] [CrossRef]
- Sun, G.; Li, H.; Wang, Y.; Zhang, B.; Zhang, S. Zebrafish complement factor H and its related genes: Identification, evolution, and expression. Funct. Integr. Genom. 2010, 10, 577–587. [Google Scholar] [CrossRef]
- Qi, P.; Wu, B.; Guo, B.; Zhanf, C.; Xu, K. The complement factor H (CFH) and its related protein 2 (CFHR2) mediating immune response in large yellow croaker Larimichthys crocea. Dev. Comp. Immunol. 2018, 84, 241–249. [Google Scholar] [CrossRef]
- Forlenza, M.; Fink, I.R.; Raes, G.; Wiegertjes, G.F. Heterogeneity of macrophage activation in fish. Dev. Comp. Immunol. 2011, 35, 1246–1255. [Google Scholar] [CrossRef]
- Chen, L.; Huang, R.; Zhu, D.; Wang, Y.; Mehjabin, R.; Li, Y.; Liao, L.; He, L.; Zhu, Z.; Wang, Y. Cloning of six serpin genes and their responses to GCRV infection in grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2019, 86, 93–100. [Google Scholar] [CrossRef]
- Zhu, J.; Fu, Q.; Ao, Q.; Tan, Y.; Luo, Y.; Jiang, H.; Li, C.; Gan, X. Transcriptomic profiling analysis of tilapia (Oreochromis niloticus) following Streptococcus agalactiae challenge. Fish Shellfish Immun. 2017, 62, 202–212. [Google Scholar] [CrossRef]
- Li, M.F.; Sun, L. Characterization of a teleost membrane-associated protein that is involved in the regulation of complement activation and bacterial infection. Dev. Comp. Immunol. 2018, 79, 142–149. [Google Scholar] [CrossRef]
- Koronega, H.; Kono, T.; Sakai, M. Isolation of seven IL-17 family genes from the Japanese pufferfish Takifugu rubripes. Fish Shellfish Immun. 2010, 28, 809–818. [Google Scholar] [CrossRef]
- Wang, T.; Martin, S.A.M.; Secombes, C.J. Two interleukin-17C-like genes exist in rainbow trout Oncorhynchus mykiss that are differentially expressed and modulated. Dev. Comp. Immunol. 2010, 34, 491–500. [Google Scholar] [CrossRef]
- Ding, Y.; Ao, J.; Chen, X. Comparative study of interleukin-17C (IL-17C) and IL-17D in large yellow croaker Larimichthys crocea reveals their similar but differential functional activity. Dev. Comp. Immunol. 2017, 76, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Hulpke, S.; Tampé, R. The MHC I loading complex: A multitasking machinery in adaptive immunity. Trends Biochem. Sci. 2013, 8, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Müller-Eberhard, H.J. The membrane attack complex of complement. Annu. Rev. Immunol. 1986, 4, 503–528. [Google Scholar]
- Wang, T.; Zhang, J. CsPTX1, a pentraxin of Cynoglossus semilaevis, is an innate immunity factor with antibacterial effects. Fish Shellfish Immunol. 2016, 56, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Barbé, F.; Douglas, T.; Saleh, M. Advances in Nod-like receptors (NLR) biology. Cytokine Growth Factor Rev. 2014, 25, 681–697. [Google Scholar] [CrossRef]
- Holland, M.C.H.; Lambris, D. The complement system in teleosts. Fish Shellfish Immun. 2002, 12, 399–420. [Google Scholar] [CrossRef] [PubMed]
- Cordero, H.; Brinchmann, M.F.; Cuesta, A.; Meseguer, J.; Esteban, M.A. Skin mucus proteome map of European sea bass (Dicentrarchus labrax). Proteomics 2015, 15, 4007–4020. [Google Scholar] [CrossRef] [PubMed]
- Wiegertjes, G.F.; Wentzel, A.S.; Spaink, H.P.; Elks, P.M.; Fink, I.R. Polarization of immune responses in fish: The ‘macrophages first’ point of view. Mol. Immunol. 2016, 69, 146–156. [Google Scholar] [CrossRef]
- Fischer, U.; Dijkstra, J.M.; Kollner, B.; Kiryu, I.; Koppang, E.O.; Hordvik, I.; Sawamoto, Y.; Ototake, M. The ontogeny of MHC class I expression in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2005, 18, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, P.N.; Hermsen, T.T.; Van Maanen, A.; Taverne-Thiele, A.A.; Rombout, J.H.; Dixon, B.; Stet, R.J. Expression of MhcCyca class I and class II molecules in the early life history of the common carp (Cyprinus carpio L.). Dev. Comp. Immunol. 1998, 22, 493–506. [Google Scholar] [CrossRef]
- Lee, K.M.; Yamada, Y.; Okamura, A.; Tsukamoto, K.; Kaneko, T. Hyposmoregulatory ability and ion- and water-regulatory mechanisms during the leptocephalus stages of Japanese eel Anguilla japonica. Fish. Sci. 2013, 79, 77–86. [Google Scholar] [CrossRef]
- Okamura, A.; Yamada, Y.; Mikawa, N.; Horie, N.; Tsukamoto, K. Effect of salinity on occurrence of notochord deformities in Japanese eel Anguilla japonica larvae. Aquac. Int. 2016, 24, 549–555. [Google Scholar] [CrossRef]
- Politis, S.N.; Mazurais, D.; Servili, A.; Zambonino-Infante, J.L.; Miest, J.J.; Tomkiewicz, J.; Butts, I.A.E. Salinity reduction benefits European eel larvae: Insights at the morphological and molecular level. PLoS ONE 2018, 13, e0198294. [Google Scholar] [CrossRef]
- Okamoto, T.; Kurokawa, T.; Gen, K.; Murashita, K.; Nomura, K.; Kim, S.K.; Matsubara, H.; Ohta, H.; Tanaka, H. Influence of salinity on morphological deformities in cultured larvae of Japanese eel, Anguilla japonica, at completion of yolk resorption. Aquaculture 2009, 293, 113–118. [Google Scholar] [CrossRef]
- Fiol, D.F.; Kültz, D. Osmotic stress sensing and signaling in fishes. FEBS J. 2007, 274, 5790–5798. [Google Scholar] [CrossRef]
- Comrie, M.M.; Cutler, C.P.; Cramb, G. Cloning and expression of guanylin from the European eel (Anguilla anguilla). Biochem. Biophys. Res. Commun. 2001, 281, 1078–1085. [Google Scholar] [CrossRef]
- Yuge, S.; Inoue, K.; Hyodo, S.; Takei, Y. A novel guanylin family (guanylin, uroguanylin, and renoguanylin) in eels: Possible osmoregulatory hormones in intestine and kidney. J. Biol. Chem. 2003, 278, 22726–22733. [Google Scholar] [CrossRef] [PubMed]
- Kalujnaia, S.; Wilson, G.D.; Feilen, A.L.; Cramb, G. Guanylin-like peptides, guanylate cyclase and osmoregulation in the European eel (Anguilla anguilla). Gen. Comp. Endocrinol. 2009, 161, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Marshall, S.; Breves, J.P.; Doohan, E.M.; Tipsmark, C.K.; Kelly, S.P.; Robertson, G.N.; Schulte, P.M. Claudin-10 isoform expression and cation selectivity change with salinity in salt-secreting epithelia of Fundulus heteroclitus. J. Exp. Biol. 2018, 221, jeb168906. [Google Scholar] [PubMed]
- Schneider, S. Inositol transport proteins. FEBS Lett. 2015, 589, 1049–1058. [Google Scholar] [CrossRef] [PubMed]
- Tipsmark, C.K.; Luckenbach, J.A.; Madsen, S.S.; Kiilerich, P.; Borski, R.J. Osmoregulation and expression of ion transport proteins and putative claudins in the fill of Southern Flounder (Paralichthys lethostigma). Comp. Biochem. Physiol. Part A 2008, 150, 265–273. [Google Scholar] [CrossRef]
- Agarkova, I.; Perriard, J.C. The M-band: An elastic web that crosslinks thick filaments in the center of the sarcomere. Trends Cell. Biol. 2005, 15, 477–785. [Google Scholar] [CrossRef]
- Farah, C.S.; Reinach, F.C. The troponin complex and regulation of muscle contraction. FASEB J. 1995, 9, 755–767. [Google Scholar] [CrossRef]
- Lincoln, T.M.; Dey, N.; Sellak, H. Invited review: cGMP-dependent protein kinase signaling mechanisms in smooth muscle: From the regulation of tone to gene expression. J. Appl. Psychol. 2001, 3, 1421–1430. [Google Scholar] [CrossRef]
- Souza, B.R.; Romano-Silva, M.A.; Tropepe, V. Dopamine D2 receptor activity modulates Akt signaling and alters GABAergic neuron development and motor behavior in zebrafish larvae. J. Neurosci. 2011, 31, 5512–5525. [Google Scholar] [CrossRef]
- Shontz, E.C.; Soulders, C.L.; Scmidt, J.T.; Martyniuk, C.J. Domperidone upregulates dopamine receptor expression and stimulates locomotor activity in larval zebrafish (Danio rerio). Genes Brain Behav. 2018, 17, e12460. [Google Scholar] [CrossRef]
- Zhdanova, I.V.; Wang, S.Y.; Leclair, O.U.; Danilova, N.P. Melatonin promotes sleep-like state in zebrafish. Brain Res. 2001, 903, 263–268. [Google Scholar] [CrossRef]
- Bae, Y.K.; Shimizu, T.; Yabe, T.; Kim, C.H.; Hirata, T.; Nojima, H.; Muraoka, O.; Hirano, T.; Hibi, M. A homeobox gene, pnx, is involved in the formation of posterior neurons in zebrafish. Development 2003, 130, 1853–1865. [Google Scholar] [CrossRef] [PubMed]
- Kimura-Yoshida, C.; Kitajima, K.; Oda-Ishii, I.; Tian, E.; Suzuki, M.; Yamamoto, M.; Suzuki, T.; Kobayashi, M.; Aizawam, S.; Matsuo, I. Characterization of the pufferfish Otx2 cis-regulators reveals evolutionarily conserved genetic mechanisms for vertebrate head specification. Development 2004, 131, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Fricker, F.R.; Bennett, D.L. The role of neuregulin-1 in the response to nerve injury. Future Neurol. 2011, 6, 809–822. [Google Scholar] [CrossRef] [PubMed]
- Hall, Z.J.; Tropepe, V. Movement maintains forebrain neurogenesis via peripheral neural feedback in larval zebrafish. Elife 2018, 7, e31045. [Google Scholar] [CrossRef] [PubMed]
- Soni, B.G.; Foster, R.G. A novel and ancient vertebrate opsin. FEBS Lett. 1997, 406, 1873–3468. [Google Scholar] [CrossRef]
- Minamoto, T.; Shimizu, I. A novel isoform of vertebrate ancient opsin in a smelt fish, Plecoglossus altivelis. Biochem. Biophys. Res. Commun. 2002, 290, 280–286. [Google Scholar] [CrossRef]
- Kojima, D.; Mano, H.; Fukada, Y. Vertebrate ancient-long opsin: A green sensitive photoreceptive molecule present in zebrafish deep brain and retinal horizontal cells. J. Neurosci. 2000, 20, 2845–2851. [Google Scholar] [CrossRef]
- Philip, A.E.; Garcia-Fernandez, J.M.; Soni, B.G.; Lucas, R.J.; Foster, B.R.G. Vertebrate ancient (VA) opsin and extraretinal photoreception in the Atlantic salmon (Salmo salar). J. Exp. Biol. 2000, 203, 1925–1936. [Google Scholar] [CrossRef]
- Sarropoulou, E.; Kotoulas, G.; Power, D.; Geisler, R. Gene expression profiling of gilthead sea bream during early development and detection of stress-related genes by the application of cDNA microarray technology. Physiol. Genom. 2005, 23, 182–191. [Google Scholar] [CrossRef]
- Blaxter, J.H.S. Light intensity, vision and feeding in young plaice. J. Exp. Mar. Biol. Ecol. 1968, 2, 293–307. [Google Scholar] [CrossRef]
- Yahaya, S.; Lim, L.H.; Shaleh, S.R.M.; Mukai, Y.; Anraku, K.; Kawamura, G. Ontogenetic eye development and related behavioural changes in larvae and juveniles of barramundi Lates calcarifer (Bloch). Mar. Freshw. Behav. Physiol. 2011, 44, 339–348. [Google Scholar] [CrossRef]
- Okamura, A.; Sakamoto, Y.; Yamada, Y.; Tsukamoto, K. Accumulation of hyaluronan in reared Japanese eel Anguilla japonica during early ontogeny. Aquaculture 2018, 497, 220–225. [Google Scholar] [CrossRef]
- Tsukamoto, K.; Yamada, Y.; Okamura, A.; Kaneko, T.; Tanaka, H.; Miller, M.J.; Horie, N.; Mikawa, N.; Utoh, T.; Tanaka, S. Positive buoyancy in eel leptocephali: An adaptation for life in the ocean surface layer. Mar. Biol. 2009, 156, 835–846. [Google Scholar] [CrossRef]
- Butts, I.A.E.; Sørensen, S.R.; Politis, S.N.; Tomkiewicz, J. First-feeding by European eel larvae: A step towards closing the life cycle in captivity. Aquaculture 2016, 464, 451–458. [Google Scholar] [CrossRef]
- Li, D.Y.; Smith, D.G.; Hardeland, R.; Yang, M.Y.; Xu, H.L.; Zhang, L.; Yin, H.D.; Zhu, Q. Melatonin receptor genes in vertebrates. Int. J. Mol. Sci. 2013, 14, 11208–11223. [Google Scholar] [CrossRef] [PubMed]
- Byun, J.H.; Hyeon, J.Y.; Kim, E.S.; Kim, S.K.; Hur, S.P.; Kim, S.J.; Takemura, A. Daily variation of D2 dopamine receptor transcription in the brain of the Japanese eel Anguilla japonica and its regulation with dopamine and melatonin. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 240, 11051. [Google Scholar] [CrossRef]
- Mohanty, B.P.; Mahanty, A.; Mitra, T.; Parija, S.C.; Mohanty, S. Heat shock proteins in stress in teleosts. In Regulation of Heat Shock Protein Responses; Asea, A., Kaur, P., Eds.; Springer: Cham, Switzerland, 2018; Volume 13, pp. 71–94. [Google Scholar]
- Yengkokpam, S.; Pal, A.K.; Sahu, N.P.; Jain, K.K.; Dalvi, R.; Misra, S.; Debnath, D. Metabolic modulation in Labeo rohita fingerlings during starvation: Hsp70 expression and oxygen consumption. Aquaculture 2008, 285, 234–237. [Google Scholar] [CrossRef]
- Dar, S.A.; Srivastava, P.P.; Varghese, T.; Nazir, M.I.; Gupta, S.; Krishna, G. Temporal changes in superoxide dismutase, catalase, and heat shock protein 70 gene expression, cortisol and antioxidant enzymes activity of Labeo rohita fingerlings subjected to starvation and refeeding. Gene 2019, 692, 94–101. [Google Scholar] [CrossRef]



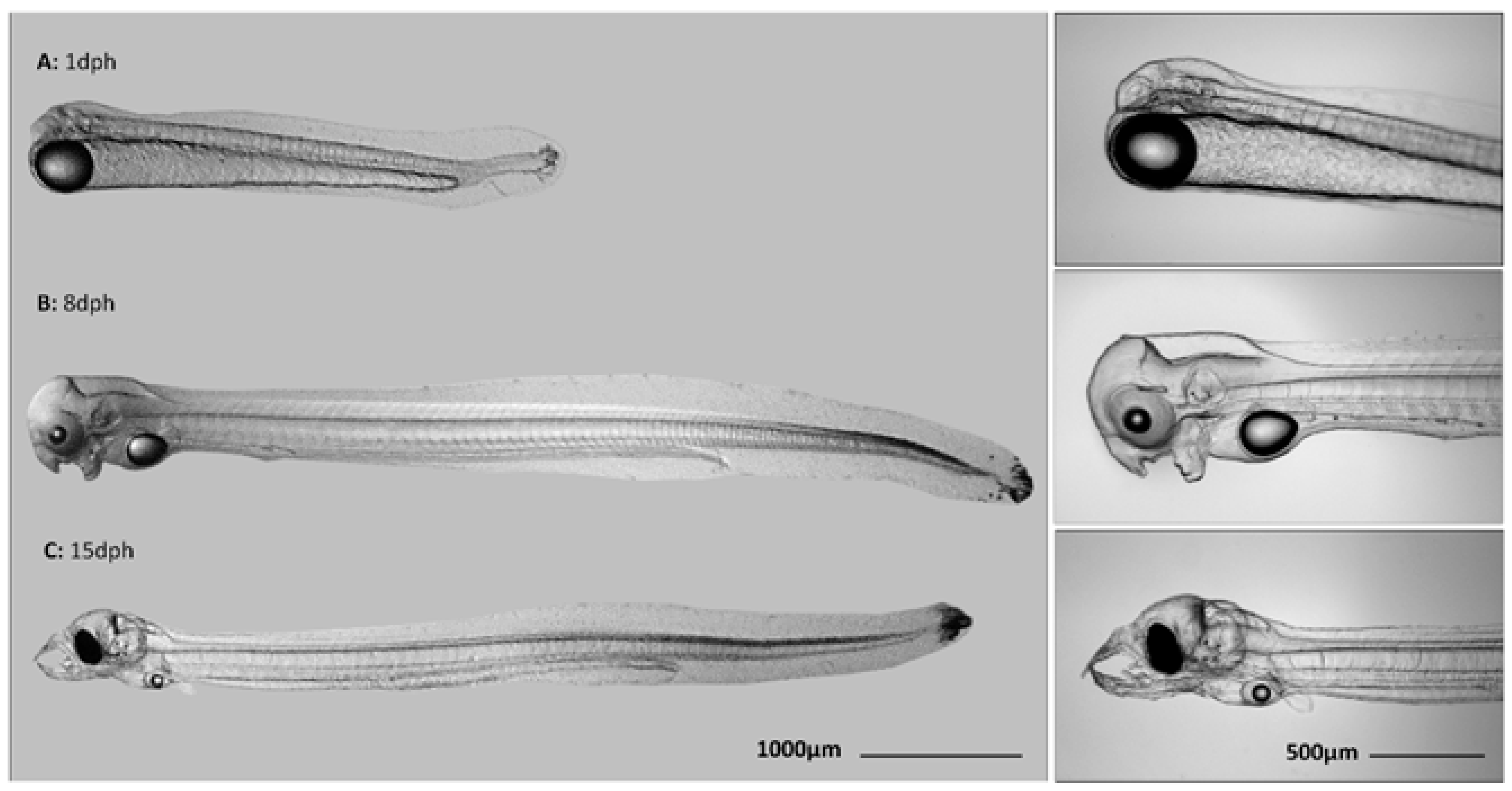{kind=link}
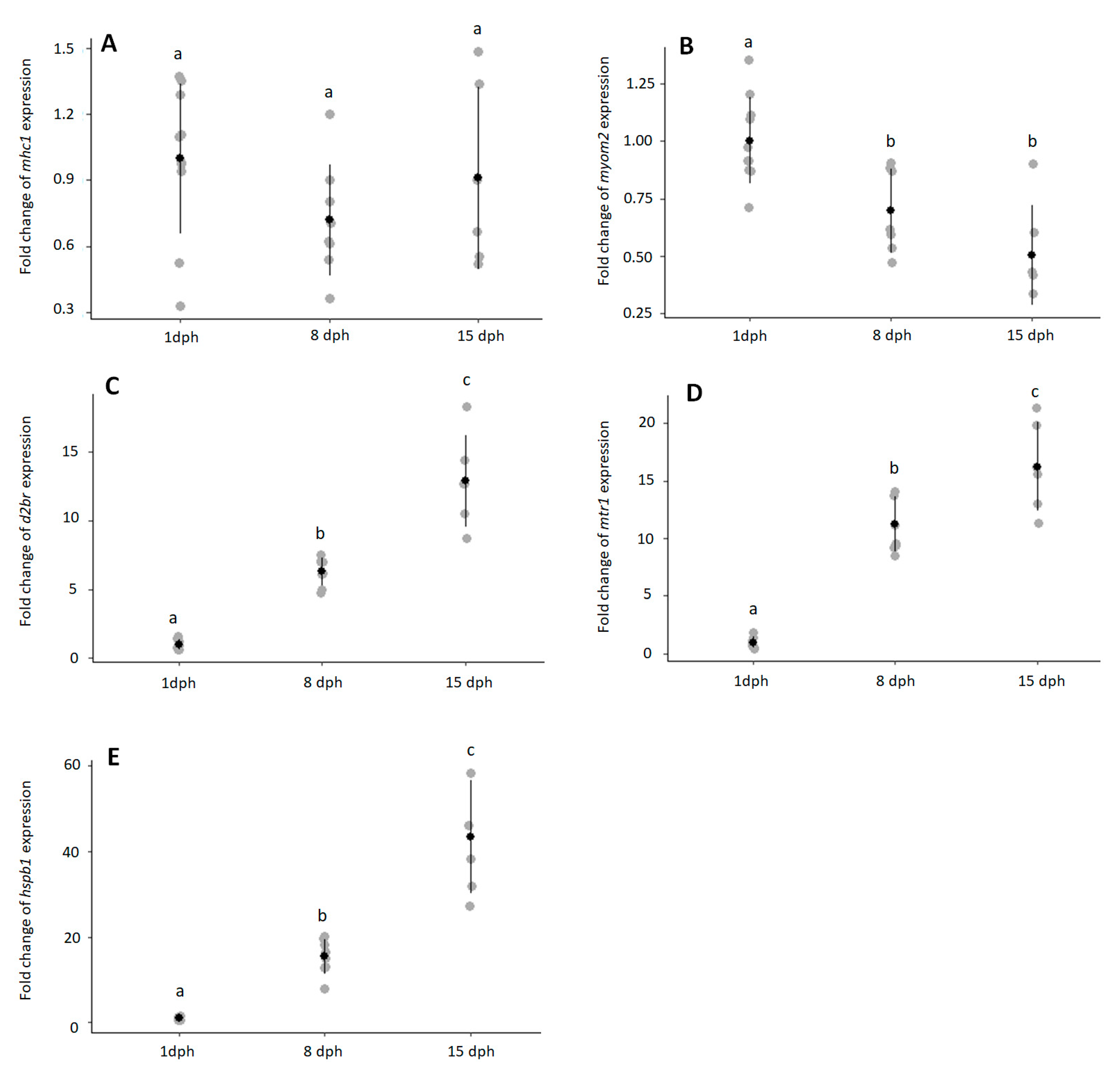{kind=link}
| Female ID | Hatch (hpf) | Longevity (dph) | Larvae Viability |
|---|---|---|---|
| 847C | 51 | 1 | Non-viable |
| B43C | 51 | 3 | Non-viable |
| F8E1 | 72 | 1 | Non-viable |
| 044B | 52 | 16 | Viable |
| D785 | 60 | 22 | Viable |
| AB79 | 59 | 7 | Viable |
| Target Genes | Abv. | Accession Number | Primer Sequences | T° | Product Size (bp) | Efficiency (%) |
|---|---|---|---|---|---|---|
| 60s ribosomal protein l36 | L36 | G | FW: CCTGACCAAGCAGACCAAGT | 62 | 160 | 91 |
| RV: TCTCTTTGCACGGATGTGAG | ||||||
| Dopamine receptor-2B | D2br | DQ789977 | FW: CACGCTACAGCTCCAAAAGAA | 60 | 186 | 92 |
| RV: TGAAGGGGACATAGAAGGACAC | ||||||
| Melatonin receptor | MT1R | Contig sequence | FW: CGACAAGAACTCCCTGTGCTA | 62 | 175 | 92 |
| RV: CAGGATGAAGTGGAAGAAGACC | ||||||
| M-protein | Myom2 | Contig sequence | FW: GATGCAAAGATCACTCAGTCCA | 62 | 160 | 106 |
| RV: CGTATTTGCCTTTGTCCTTCTC | ||||||
| MHC-I | MHCI | Contig sequence | FW: CATGTGACCGGATTCTACCC | 64 | 200 | 91 |
| RV: TGTTTCAGGCTCTTGTGCTGT | ||||||
| Hspb1 | Hspb1 | Contig sequence | FW: GGGGCATATCCGAGATCAA | 62 | 104 | 93 |
| RV: GACTCCATCCCTGGTCTTCAC |
| Description | Fold Change |
|---|---|
| Interleukin-17C | 29.0 |
| Complement decay-accelerating factor isoform X1 | 5.2 |
| Pentraxin fusion protein-like | 2.9 |
| Nlrp 12 (NACHT, LRR and PYD domains-containing protein 12-like) | 2.8 |
| Leukocyte elastase inhibitor | 2.6 |
| Arginase-2, mitochondrial | 2.3 |
| Complement factor H-like | 2.3 |
| Complement component C7 | −7.6 |
| MHC1 (Class I histocompatibility antigen, F10 alpha chain-like) | −17.7 |
| Description | Fold Change |
|---|---|
| Extracellular calcium-sensing receptor-like | 39.4 |
| Claudin-4 | 7.1 |
| Sodium/myo-inositol cotransporter-like | 3.9 |
| Guanylin precursor | 2.9 |
| Claudin-10-like isoform X2 | 2.8 |
| Claudin-16-like isoform X1 | −7.9 |
| Description | Fold Change |
|---|---|
| Muscle development | |
| Heat-shock protein beta-1 like | 5.9 |
| cGMP-dependent protein kinase 1-like | −4.0 |
| Troponin I slow skeletal muscle-like | −4.8 |
| Myom2 (M-protein, striated muscle-like isoform X1) | −10.5 |
| Neural development | |
| Pro-neuregulin-1, membrane-bound isoform X1 | 6.7 |
| Csmd3 (CUB and sushi domain-containing protein 3) | 2.6 |
| Homeobox protein OTX2 | −2.3 |
| Homeobox protein pnx-like isoform X1 | −2.3 |
| Adam22 (disintegrin and metalloproteinase domain-containing protein 22) | −2.5 |
| MT1R (MT1 melatonin receptor) | −3.4 |
| Protocadherin-16-like, partial | −5.6 |
| D2br (dopamine D2B receptor) | −9.4 |
| Sensory system | |
| Vertebrate ancient opsin-like | 3.2 |
| LIM/homeobox protein Lhx2 | −2.2 |
| Norrin-like | −2.5 |
| Tmc1 (putative transmembrane channel-like protein 1) | −3.0 |
| Wnt signaling | |
| Norrin-like | −2.5 |
| Ptpro (Receptor-type tyrosine-protein phosphatase O, partial) | −4.3 |
| CXXC-type zinc finger protein 4 | −9.0 |
| Various functions | |
| T-box transcription factor TBX1 isoform X4 | 2.3 |
| Homeobox protein DLX-6 | −2.4 |
| Homeobox protein DLX-2 | −2.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jéhannet, P.; Palstra, A.P.; Heinsbroek, L.T.N.; Kruijt, L.; Dirks, R.P.; Swinkels, W.; Komen, H. What Goes Wrong during Early Development of Artificially Reproduced European Eel Anguilla anguilla? Clues from the Larval Transcriptome and Gene Expression Patterns. Animals 2021, 11, 1710. https://doi.org/10.3390/ani11061710
Jéhannet P, Palstra AP, Heinsbroek LTN, Kruijt L, Dirks RP, Swinkels W, Komen H. What Goes Wrong during Early Development of Artificially Reproduced European Eel Anguilla anguilla? Clues from the Larval Transcriptome and Gene Expression Patterns. Animals. 2021; 11(6):1710. https://doi.org/10.3390/ani11061710
Chicago/Turabian StyleJéhannet, Pauline, Arjan P. Palstra, Leon T. N. Heinsbroek, Leo Kruijt, Ron P. Dirks, William Swinkels, and Hans Komen. 2021. "What Goes Wrong during Early Development of Artificially Reproduced European Eel Anguilla anguilla? Clues from the Larval Transcriptome and Gene Expression Patterns" Animals 11, no. 6: 1710. https://doi.org/10.3390/ani11061710
APA StyleJéhannet, P., Palstra, A. P., Heinsbroek, L. T. N., Kruijt, L., Dirks, R. P., Swinkels, W., & Komen, H. (2021). What Goes Wrong during Early Development of Artificially Reproduced European Eel Anguilla anguilla? Clues from the Larval Transcriptome and Gene Expression Patterns. Animals, 11(6), 1710. https://doi.org/10.3390/ani11061710