
Genomics of Adaptations in Ungulates

 ,
,  , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Understanding the Genetic Basis of Ungulate Adaptations
3. The Adaptive Evolution of Ungulates in Different Habitats
3.1. Arid and Semi-Arid Habitats
3.2. High-Altitude Habitats
3.3. Savanna Habitats
3.4. Marine Habitats
3.5. Arctic Habitats
3.6. Processes Underlying Adaptive Changes in Domesticated Ungulates
3.7. Processes Underlying Adaptive Changes in Feral Ungulates

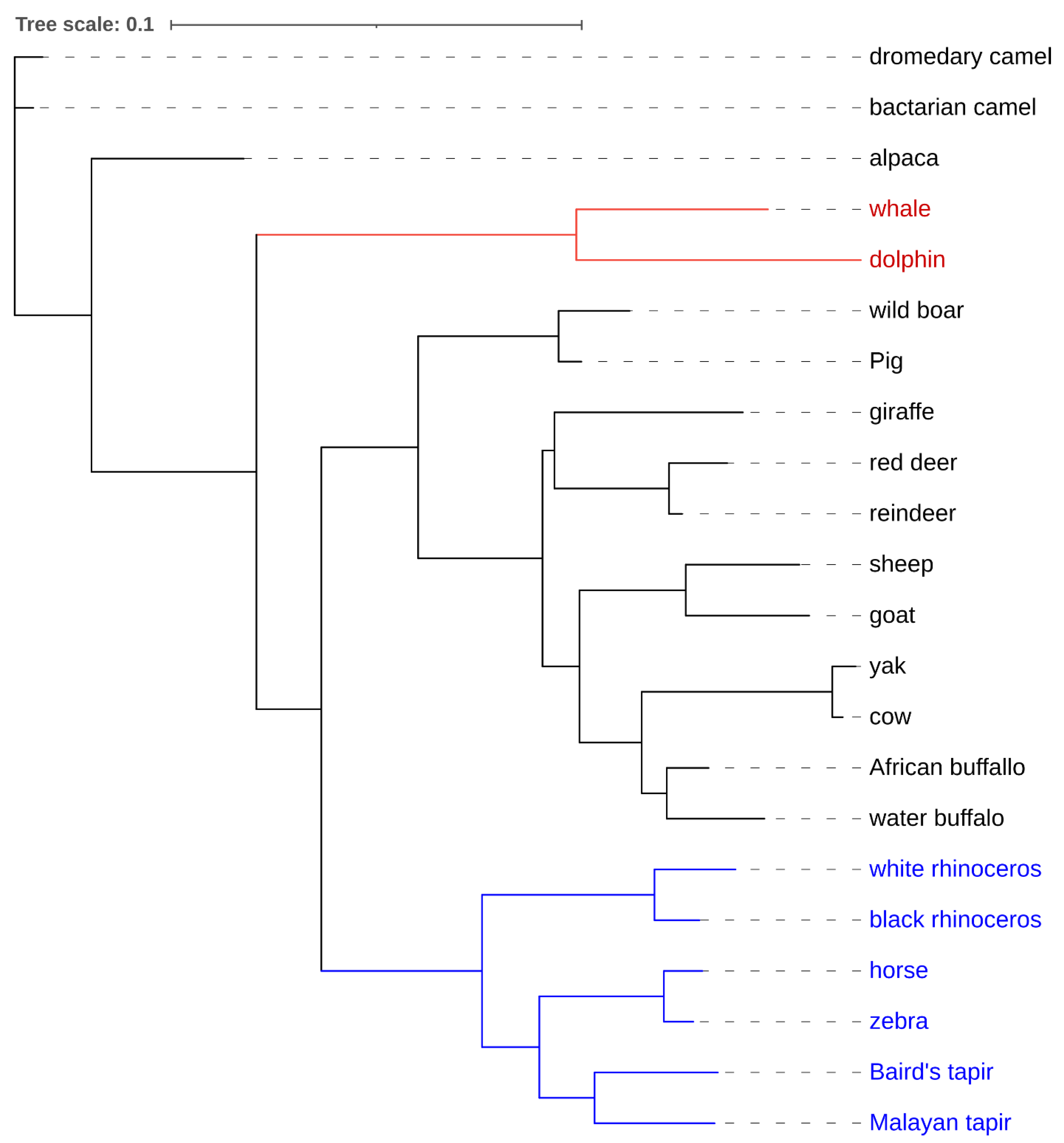{kind=link}
| Environment | Selective Pressure (s) | Selection Signature | Genes under Selection in Ungulates | Biological Functions of Candidate Genes |
|---|---|---|---|---|
| Desert/Arid | Low water availability | Population differentiation and increased frequency of derived alleles | Red deer (CP2U1) [3], Camel (CYP2J, CYP2E, AQP1, AQP2, AQP3) [5,36], sheep (NXA6, GPX3, GPX7, PTGS2, CPA3, CPVL, ECE1, CALM2, CACNA2D1, KCNJ5, and COX2) [55] | Water-salt balance, regulating water retention and reabsorption |
| Airborne dust and allergic diseases | Population differentiation, excess of long haplotype and Sequence altering mutant (rapidly evolving genes) | Red deer (TRAF2 and IL1R1) [3], camel (FOXP3, CX3CR1, CYSLTR2, and SEMA4A) [5], Fat-tail sheep (ZBP1, PRDX1, MAST2, and LURAP) [71], Bakri goat and sheep (GRIA1, IL2, IL7, IL21, IL1R1) [69] | Defend against airborne dusts | |
| High UV exposure | Excess of long haplotypes and rapidly evolving genes | Red deer (LAMB1, LAMB2, CYC, FANCF, and GPR98) [3], camel (OPN1SW, CX3CR1, and CNTFR) [5] | Ocular development, visual protection, and photoreceptor cell synapses | |
| Excess of long haplotypes | Red deer (SLX4, FANCF, FANCG, FANCI, ATR, and POLH) [3], Fat-tail sheep (PMS1, SPO11, RAD54L, MUTYH, CHEK2, POLR2D, and CMPK1) [71] | DNA repair | ||
| Sequence altering mutant (rapidly evolving genes) | Desert goat (ABCA12, ASCL4, and UVSSA) [72] | Skin barrier development and function | ||
| High temperature | Population differentiation and excess of long haplotypes | Red deer (GNAI2, FZD4, MP2K2, CREB3, CBP, GNAO, TF7L2, and GNAO) [3], goat (MTOR, and MAPK3) [114], Fat-tail sheep (ERCC3, and TGM3) [71], Bakri goat and sheep (FGF2, GNAI3, and PLCB1) [69] | Response to thermal stress | |
| Xenobiotic compounds | Population differentiation and excess of long haplotypes | Red deer (CP2U1, CP3AS, and CP3AO) [3] | Plant secondary metabolism | |
| Arctic | Long light and dark periods | Population differentiation, rapidly evolving genes and copy number variable genes | Yakutian Horse (LECT2, and FBXL21) [11], Wooly mammoth (HRH3, PER2, and BMAL1) [13], reindeer (GRIA1, and OPN4B) [12,58] | Regulation of the circadian clock |
| Low temperatures | Population differentiation and copy number variable genes, Sequence altering mutant (rapidly evolving genes) | Yakutian Horse (ACADSB, ATP1A2, CYP11B2, HSPG2, and PRKG1) [11], Wooly mammoth (DLK1, and TRPV3) [13], Yakutian cattle (DNAJC9, SOCS3, TRPC7, SLC8A1 GLP1R, PKLR, and TCF7L2) [42], Reindeer (SCN11A, and SILT2) [58] | Thermoregulation | |
| Lipid metabolism | Sequence altering mutant (rapidly evolving genes) | Reindeer (APOB and FASN), woolly mammoth (CRH) [13] | Lipid metabolism | |
| High-altitude | Hypoxia | Population differentiation, sequence altering mutant (rapidly evolving genes), excess of long haplotypes | Tibetan wild boar (ALB, ECE1, GNG2, and PIK3C2G) [52], Yak (ADAM17, ARG2, and MMP3) [8], goat (CDK2, SOCS2, NOXA1, and ENPEP) [157], Tibetan sheep (EPAS1, CRYAA, LONP1, NF1, DPP4, SOD1, PPARG, and SOCS2) [78], Tibetan pig (EPAS1, HIF1A, RNF4, TNFSF1, PDE1A, and PDE3) [77] | Hypoxia response |
| Low temperature | Population differentiation and sequence altering mutant (rapidly evolving genes) | Tibetan wild boar (AEBP1, DGAT1, FABP2, LEPR, and PTPN1) [52], Yak (GCNT3, HSD17B12, WHSC1, and GLUL) [8], Tibetan sheep (DPP4, and PPARG) [78] | Tolerance to cold | |
| Intense UV radiations | Sequence altering mutant (rapidly evolving genes) | Tibetan wild boar (BCL3, ERCC4, ERCC6, REV1, USF1, and ZRANB3) [52] | DNA repair, response to radiations | |
| Marine | Low oxygen levels | Sequence altering mutant (rapidly evolving genes) | Whales (PRDX1, PRDX2, and GPX2) [96], Minke whale (GPX2, ODC1, GSR, GGT6, GGT7, GCLC, and ANPEP) [67], cetaceans (ALDOA, ENO2, CS, ATP6V0A4, LHPP, NDUFA9, and NDUFV3) [158] | Response to hypoxia |
| Salty water | Sequence altering mutant (rapidly evolving genes) | Minke whale (AGTR1, ANPEP, LNPEP, MME, and THOP1) [67], dolphin (TSPO2, EPGN, PLN, EDN2, PLA2G5, and KCNJ2) [15] | Salt water balance | |
| Aquatic environment | Sequence altering mutant (rapidly evolving genes) | Minke whale (HOXA5, HOXB1, HOXB2, HOXB5, HOXD12 and HOXD13) [67] | Morphological adaptation to swimming | |
| Prolonged, deep diving | Sequence altering mutant (rapidly evolving genes) | Whales and dolphins (GSTA1) [96], dolphin (APOA2, APOC4, APOO, FABP4, SERINC4, CCDC129, PLA2G5, PNLIPRP3, RARRES2, and NR1I3) [15], cetaceans/whales (LDHA, LDHD, PC, PCK1, FBP1, and GPI) [158] | Energy metabolism | |
| Cold temperature | Sequence altering mutation (rapidly evolving genes) | Minke whale (NPY) [66] | Thermoregulation | |
| Domesticated | Productivity | Population differentiation, rapidly evolving genes, copy number variable genes | Goat (LRP1, PLIN4 and FASN) [4], pig (ACACA, ANKRD23, GM2A, KIT, MOGAT2, MTTP, FASN, SGMS1, SLC27A6, and RETSAT) [132], donkey (TBX3, NCAPG, LOCR, BCOR, CDKL5, and ACSL4) [125], cattle (MUC1) [42] | Regulation body weight, body size, milk production |
| Domestication | Population differentiation, sequence altering mutant (rapidly evolving genes), copy number variable genes and excess of long haplotypes | Goat (FGF9, IGF1, ASIP, KITLG, HTT, GNA11, OSTM1, ATRN, GNAQ, HELLS, MUTED, VPS33A, ADAMTS20, MITF and OCA2) [4,114,145], donkey (ASIP and KTLG) [125], sheep and goat (KITLG, HMGI-C, NBEA, and MTMR7) [105], pigs (ESR1, and AHR) [159,160] | Coat color, litter size, fatty acid composition, wool crimping | |
| Domestication | Population differentiation, excess of long haplotypes, sequence altering mutant (rapidly evolving genes), and copy number variable genes | Goat (HTR3A, STMI, and PRMI) [4,49], horse (VDAC1, and GRID1) [127], pigs (NRTN, SEMA3C, PLXNC1, AAK1, RAB35 FRS2, APBA2, MC4R, RCAN1, and BAIAP3) [161] and domestic yak (GRIN2D and NTN5) [130] | Tameness, less aggressiveness, reduced fear to humans | |
| Increased pathogens | Excess of long haplotypes, sequence altering mutant (rapidly evolving genes), population differentiation and copy number variable genes, adaptive introgression | Goat (IL10RB, IFNLR1, BCL2L1, ERBB2, ENO1, CFH, TRIM5, and MUC6) [4,49,114], cattle (IFNAR1, IFNAR2, IL10RB, NOD2, CD96, CD14, GZMB, IL17A, PFKM, ADAM17, SIRPA IFNAR2, IFNG, CD34, TREM1, TREML1, FCER1A, IL23R, IL24, IL15, and LEAP2) [41,42], pig (IL1B, CD36, CD68, CD163, CRP, and IFIT1) [121] | Host innate immune response, gastrointestinal pathogen resistance, disease resistance |
4. Future Perspectives and Directions
5. Conclusions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Applegate, R.D. Groves, C., and P. Grubb. 2011. Ungulate Taxonomy. Johns Hopkins University Press, Baltimore, Maryland, 317 pp. ISBN-13 978-1-4214-0093-8 and ISBN-10 1-4214-0093-6, Price (Hardbound), $100.00. J. Mammal. 2013, 94, 245–246. [Google Scholar] [CrossRef]
- Thewissen, J.G.M.; Cooper, L.N.; George, J.C.; Bajpai, S. From Land to Water: The Origin of Whales, Dolphins, and Porpoises. Evol. Educ. Outreach 2009, 2, 272–288. [Google Scholar] [CrossRef]
- Ababaikeri, B.; Abduriyim, S.; Tohetahong, Y.; Mamat, T.; Ahmat, A.; Halik, M. Whole-Genome Sequencing of Tarim Red Deer (Cervus Elaphus Yarkandensis) Reveals Demographic History and Adaptations to an Arid-Desert Environment. Front. Zool. 2020, 17, 31. [Google Scholar] [CrossRef]
- Dong, Y.; Zhang, X.; Xie, M.; Arefnezhad, B.; Wang, Z.; Wang, W.; Feng, S.; Huang, G.; Guan, R.; Shen, W.; et al. Reference Genome of Wild Goat (Capra Aegagrus) and Sequencing of Goat Breeds Provide Insight into Genic Basis of Goat Domestication. BMC Genom. 2015, 16, 431. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Guang, X.; Al-Fageeh, M.B.; Cao, J.; Pan, S.; Zhou, H.; Zhang, L.; Abutarboush, M.H.; Xing, Y.; Xie, Z.; et al. Camelid Genomes Reveal Evolution and Adaptation to Desert Environments. Nat. Comm. 2014, 5, 5188. [Google Scholar] [CrossRef]
- Agaba, M.; Ishengoma, E.; Miller, W.C.; McGrath, B.C.; Hudson, C.N.; Bedoya Reina, O.C.; Ratan, A.; Burhans, R.; Chikhi, R.; Medvedev, P.; et al. Giraffe Genome Sequence Reveals Clues to Its Unique Morphology and Physiology. Nat. Commun. 2016, 7, 11519. [Google Scholar] [CrossRef] [PubMed]
- Ishengoma, E.; Agaba, M.; Cavener, D.R. Evolutionary Analysis of Vision Genes Identifies Potential Drivers of Visual Differences between Giraffe and Okapi. PeerJ 2017, 5, e3145. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Q.; Zhang, G.; Ma, T.; Qian, W.; Wang, J.; Ye, Z.; Cao, C.; Hu, Q.; Kim, J.; Larkin, D.M.; et al. The Yak Genome and Adaptation to Life at High Altitude. Nat. Genet. 2012, 44, 946–949. [Google Scholar] [CrossRef]
- Ge, R.L.; Cai, Q.; Shen, Y.Y.; San, A.; Ma, L.; Zhang, Y.; Yi, X.; Chen, Y.; Yang, L.; Huang, Y.; et al. Draft Genome Sequence of the Tibetan Antelope. Nat. Commun. 2013, 4, 1858. [Google Scholar] [CrossRef]
- Blix, A.S. Adaptations to Polar Life in Mammals and Birds. J. Exp. Biol. 2016, 219, 1093. [Google Scholar] [CrossRef]
- Librado, P.; Der Sarkissian, C.; Ermini, L.; Schubert, M.; Jónsson, H.; Albrechtsen, A.; Fumagalli, M.; Yang, M.A.; Gamba, C.; Seguin-Orlando, A.; et al. Tracking the Origins of Yakutian Horses and the Genetic Basis for Their Fast Adaptation to Subarctic Environments. Proc. Natl. Acad. Sci. USA 2015, 112, E6889. [Google Scholar] [CrossRef]
- Lin, Z.; Chen, L.; Chen, X.; Zhong, Y.; Yang, Y.; Xia, W.; Liu, C.; Zhu, W.; Wang, H.; Yan, B.; et al. Biological Adaptations in the Arctic Cervid, the Reindeer (Rangifer Tarandus). Science 2019, 364, eaav6312. [Google Scholar] [CrossRef] [PubMed]
- Lynch, V.J.; Bedoya-Reina, O.C.; Ratan, A.; Sulak, M.; Drautz-Moses, D.I.; Perry, G.H.; Miller, W.; Schuster, S.C. Elephantid Genomes Reveal the Molecular Bases of Woolly Mammoth Adaptations to the Arctic. Cell Rep. 2015, 12, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Árnason, U.; Lammers, F.; Kumar, V.; Nilsson, M.A.; Janke, A. Whole-Genome Sequencing of the Blue Whale and Other Rorquals Finds Signatures for Introgressive Gene Flow. Sci. Adv. 2018, 4, eaap9873. [Google Scholar] [CrossRef] [PubMed]
- McGowen, M.R.; Grossman, L.I.; Wildman, D.E. Dolphin Genome Provides Evidence for Adaptive Evolution of Nervous System Genes and a Molecular Rate Slowdown. Proc. R. Soc. B: Biol. Sci. 2012, 279, 3643–3651. [Google Scholar] [CrossRef]
- Sun, Y.B.; Zhou, W.P.; Liu, H.Q.; Irwin, D.M.; Shen, Y.Y.; Zhang, Y.P. Genome-Wide Scans for Candidate Genes Involved in the Aquatic Adaptation of Dolphins. Genome Biol. Evol. 2013, 5, 130–139. [Google Scholar] [CrossRef]
- Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2016, 44, D67–D72. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A Multiple Sequence Alignment Method with Reduced Time and Space Complexity. BMC Bioinform. 2004, 5, 1–19. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021. [Google Scholar] [CrossRef]
- Roffler, G.H.; Amish, S.J.; Smith, S.; Cosart, T.; Kardos, M.; Schwartz, M.K.; Luikart, G. SNP Discovery in Candidate Adaptive Genes Using Exon Capture in a Free-Ranging Alpine Ungulate. Mol. Ecol. Resour. 2016, 16, 1147–1164. [Google Scholar] [CrossRef]
- Pardo-Diaz, C.; Salazar, C.; Jiggins, C.D. Towards the Identification of the Loci of Adaptive Evolution. Methods Ecol. Evol. 2015, 6, 445–464. [Google Scholar] [CrossRef] [PubMed]
- Lewontin, R.C.; Krakauer, J. Distribution of Gene Frequency as a Test of the Theory of the Selective Neutrality of Polymorphisms. Genetics 1973, 74, 175–195. [Google Scholar] [CrossRef] [PubMed]
- Tajima, F. Statistical Method for Testing the Neutral Mutation Hypothesis by DNA Polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Sabeti, P.C.; Reich, D.E.; Higgins, J.M.; Levine, H.Z.P.; Richter, D.J.; Schaffner, S.F.; Gabriel, S.B.; Platko, J.V.; Patterson, N.J.; McDonald, G.J.; et al. Detecting Recent Positive Selection in the Human Genome from Haplotype Structure. Nature 2002, 419, 832–837. [Google Scholar] [CrossRef]
- Vitti, J.J.; Grossman, S.R.; Sabeti, P.C. Detecting Natural Selection in Genomic Data. Annu. Rev. Genet. 2013, 47, 97–120. [Google Scholar] [CrossRef]
- Hudson, R.R.; Kreitman, M.; Aguadé, M. A Test of Neutral Molecular Evolution Based on Nucleotide Data. Genetics 1987, 116, 153–159. [Google Scholar] [CrossRef]
- McDonald, J.H.; Kreitman, M. Adaptive Protein Evolution at the Adh Locus in Drosophila. Nature 1991, 351, 652–654. [Google Scholar] [CrossRef]
- Goldman, N.; Yang, Z. A Codon-Based Model of Nucleotide Substitution for Protein-Coding DNA Sequences. Mol. Biol. Evol. 1994, 11, 725–736. [Google Scholar]
- Redon, R.; Ishikawa, S.; Fitch, K.R.; Feuk, L.; Perry, G.H.; Andrews, T.D.; Fiegler, H.; Shapero, M.H.; Carson, A.R.; Chen, W. Global Variation in Copy Number in the Human Genome. Nature 2006, 444, 444–454. [Google Scholar] [CrossRef] [PubMed]
- Gamazon, E.R.; Stranger, B.E. The Impact of Human Copy Number Variation on Gene Expression. Brief. Funct Genom. 2015, 14, 352–357. [Google Scholar] [CrossRef]
- Ba, H.; Cai, Z.; Gao, H.; Qin, T.; Liu, W.; Xie, L.; Zhang, Y.; Jing, B.; Wang, D.; Li, C. Chromosome-Level Genome Assembly of Tarim Red Deer, Cervus Elaphus Yarkandensis. Sci. Data 2020, 7, 187. [Google Scholar] [CrossRef]
- Bana, N.A.; Nyiri, A.; Nagy, J.; Frank, K.; Nagy, T.; Stéger, V.; Schiller, M.; Lakatos, P.; Sugár, L.; Horn, P.; et al. The Red Deer Cervus Elaphus Genome CerEla1.0: Sequencing, Annotating, Genes, and Chromosomes. Mol. Genet. Genom. 2018, 293, 665–684. [Google Scholar] [CrossRef] [PubMed]
- Bahbahani, H.; Musa, H.H.; Wragg, D.; Shuiep, E.S.; Almathen, F.; Hanotte, O. Genome Diversity and Signatures of Selection for Production and Performance Traits in Dromedary Camels. Front. Genet. 2019, 10, 893. [Google Scholar] [CrossRef] [PubMed]
- Hedayat-Evrigh, N.; Khalkhali-Evrigh, R.; Bakhtiarizadeh, M.R. Genome-Wide Identification and Analysis of Variants in Domestic and Wild Bactrian Camels Using Whole-Genome Sequencing Data. Int. J. Genom. 2020, 2020, 2430846. [Google Scholar] [CrossRef] [PubMed]
- Jirimutu, C.; Wang, Z.; Ding, G.; Chen, G.; Sun, Y.; Sun, Z.; Zhang, H.; Wang, L.; Hasi, S.; Zhang, Y.; et al. Genome Sequences of Wild and Domestic Bactrian Camels. Nat. Commun. 2012, 3, 1202. [Google Scholar] [CrossRef]
- Fitak, R.R.; Mohandesan, E.; Corander, J.; Burger, P.A. The de Novo Genome Assembly and Annotation of a Female Domestic Dromedary of North African Origin. Mol. Ecol. Resour. 2016, 16, 314–324. [Google Scholar] [CrossRef]
- Fitak, R.R.; Mohandesan, E.; Corander, J.; Yadamsuren, A.; Chuluunbat, B.; Abdelhadi, O.; Raziq, A.; Nagy, P.; Walzer, C.; Faye, B. Genomic Signatures of Domestication in Old World Camels. Commun. Biol. 2020, 3, 1–10. [Google Scholar] [CrossRef]
- Elbers, J.P.; Rogers, M.F.; Perelman, P.L.; Proskuryakova, A.A.; Serdyukova, N.A.; Johnson, W.E.; Horin, P.; Corander, J.; Murphy, D.; Burger, P.A. Improving Illumina Assemblies with Hi-C and Long Reads: An Example with the North African Dromedary. Mol. Ecol. Resour. 2019, 19, 1015–1026. [Google Scholar] [CrossRef]
- Canavez, F.C.; Luche, D.D.; Stothard, P.; Leite, K.R.M.; Sousa-Canavez, J.M.; Plastow, G.; Meidanis, J.; Souza, M.A.; Feijao, P.; Moore, S.S.; et al. Genome Sequence and Assembly of Bos Indicus. J. Hered. 2012, 103, 342–348. [Google Scholar] [CrossRef]
- Elsik, C.G.; Tellam, R.L.; Worley, K.C. The Genome Sequence of Taurine Cattle: A Window to Ruminant Biology and Evolution. Science 2009, 324, 522. [Google Scholar] [CrossRef]
- Weldenegodguad, M.; Popov, R.; Pokharel, K.; Ammosov, I.; Ming, Y.; Ivanova, Z.; Kantanen, J. Whole-Genome Sequencing of Three Native Cattle Breeds Originating From the Northernmost Cattle Farming Regions. Front. Genet. 2019, 9, 728. [Google Scholar] [CrossRef]
- Zimin, A.V.; Delcher, A.L.; Florea, L.; Kelley, D.R.; Schatz, M.C.; Puiu, D.; Hanrahan, F.; Pertea, G.; Van Tassell, C.P.; Sonstegard, T.S.; et al. A Whole-Genome Assembly of the Domestic Cow, Bos Taurus. Genome Biol. 2009, 10, R42. [Google Scholar] [CrossRef]
- Wade, C.M.; Giulotto, E.; Sigurdsson, S.; Zoli, M.; Gnerre, S.; Imsland, F.; Lear, T.L.; Adelson, D.L.; Bailey, E.; Bellone, R.R.; et al. Genome Sequence, Comparative Analysis, and Population Genetics of the Domestic Horse. Science 2009, 326, 865. [Google Scholar] [CrossRef]
- Berihulay, H.; Li, Y.; Gebrekidan, B.; Gebreselassie, G.; Liu, X.; Jiang, L.; Ma, Y. Whole Genome Resequencing Reveals Selection Signatures Associated With Important Traits in Ethiopian Indigenous Goat Populations. Front. Genet. 2019, 10, 1190. [Google Scholar] [CrossRef]
- Bickhart, D.M.; Rosen, B.D.; Koren, S.; Sayre, B.L.; Hastie, A.R.; Chan, S.; Lee, J.; Lam, E.T.; Liachko, I.; Sullivan, S.T.; et al. Single-Molecule Sequencing and Chromatin Conformation Capture Enable de Novo Reference Assembly of the Domestic Goat Genome. Nat. Genet. 2017, 49, 643–650. [Google Scholar] [CrossRef]
- Dong, Y.; Xie, M.; Jiang, Y.; Xiao, N.; Du, X.; Zhang, W.; Tosser-Klopp, G.; Wang, J.; Yang, S.; Liang, J.; et al. Sequencing and Automated Whole-Genome Optical Mapping of the Genome of a Domestic Goat (Capra Hircus). Nat. Biotechnol. 2013, 31, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Tao, H.; Li, P.; Li, L.; Zhong, T.; Wang, L.; Ma, J.; Chen, X.; Song, T.; Zhang, H. Whole-Genome Sequencing Reveals Selection Signatures Associated with Important Traits in Six Goat Breeds. Sci. Rep. 2018, 8, 10405. [Google Scholar] [CrossRef]
- Zheng, Z.; Wang, X.; Li, M.; Li, Y.; Yang, Z.; Wang, X.; Pan, X.; Gong, M.; Zhang, Y.; Guo, Y.; et al. The Origin of Domestication Genes in Goats. Sci. Adv. 2020, 6, eaaz5216. [Google Scholar] [CrossRef] [PubMed]
- El-Khishin, D.A.; Ageez, A.; Saad, M.E.; Ibrahim, A.; Shokrof, M.; Hassan, L.R.; Abouelhoda, M.I. Sequencing and Assembly of the Egyptian Buffalo Genome. PLoS ONE 2020, 15, e0237087. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.L.; Iamartino, D.; Pruitt, K.D.; Sonstegard, T.; Smith, T.P.L.; Low, W.Y.; Biagini, T.; Bomba, L.; Capomaccio, S.; Castiglioni, B.; et al. Genome Assembly and Transcriptome Resource for River Buffalo, Bubalus Bubalis (2n = 50). GigaScience 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Tian, S.; Jin, L.; Zhou, G.; Li, Y.; Zhang, Y.; Wang, T.; Yeung, C.K.L.; Chen, L.; Ma, J.; et al. Genomic Analyses Identify Distinct Patterns of Selection in Domesticated Pigs and Tibetan Wild Boars. Nat. Genet. 2013, 45, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Xie, M.; Chen, W.; Talbot, R.; Maddox, J.F.; Faraut, T.; Wu, C.; Muzny, D.M.; Li, Y.; Zhang, W.; et al. The Sheep Genome Illuminates Biology of the Rumen and Lipid Metabolism. Science 2014, 344, 1168. [Google Scholar] [CrossRef]
- Miller, J.M.; Moore, S.S.; Stothard, P.; Liao, X.; Coltman, D.W. Harnessing Cross-Species Alignment to Discover SNPs and Generate a Draft Genome Sequence of a Bighorn Sheep (Ovis Canadensis). BMC Genom. 2015, 16, 397. [Google Scholar] [CrossRef]
- Yang, J.; Li, W.R.; Lv, F.H.; He, S.G.; Tian, S.L.; Peng, W.F.; Sun, Y.W.; Zhao, Y.X.; Tu, X.L.; Zhang, M.; et al. Whole-Genome Sequencing of Native Sheep Provides Insights into Rapid Adaptations to Extreme Environments. Mol. Biol. Evol. 2016, 33, 2576–2592. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhao, Y.; Bai, D.; Shiraigol, W.; Li, B.; Yang, L.; Wu, J.; Bao, W.; Ren, X.; Jin, B.; et al. Donkey Genome and Insight into the Imprinting of Fast Karyotype Evolution. Sci. Rep. 2015, 5, 14106. [Google Scholar] [CrossRef][Green Version]
- Taylor, R.S.; Horn, R.L.; Zhang, X.; Golding, G.B.; Manseau, M.; Wilson, P.J. The Caribou (Rangifer Tarandus) Genome. Genes 2019, 10, 540. [Google Scholar] [CrossRef]
- Weldenegodguad, M.; Pokharel, K.; Ming, Y.; Honkatukia, M.; Peippo, J.; Reilas, T.; Røed, K.H.; Kantanen, J. Genome Sequence and Comparative Analysis of Reindeer (Rangifer Tarandus) in Northern Eurasia. Sci. Rep. 2020, 10, 8980. [Google Scholar] [CrossRef]
- Mizzi, J.E.; Lounsberry, Z.T.; Brown, C.T.; Sacks, B.N. Draft Genome of Tule Elk Cervus Canadensis Nannodes. F1000Res 2017, 6, 1691. [Google Scholar] [CrossRef] [PubMed]
- Dobson, L.K. Sequencing the Genome of the North American Bison. Ph.D. Thesis, A & M University, College Station, TX, USA, 2015. [Google Scholar]
- Liu, Y.; Luo, J.; Dou, J.; Yan, B.; Ren, Q.; Tang, B.; Wang, K.; Qiu, Q. The Sequence and de Novo Assembly of the Wild Yak Genome. Sci. Data 2020, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Li, W.; Jin, J.; Cui, K.; Yan, C.; Peng, C.; Jian, Z.; Bu, P.; Price, M.; Zhang, X.; et al. The Draft Genome Sequence of Forest Musk Deer (Moschus Berezovskii). GigaScience 2018, 7. [Google Scholar] [CrossRef]
- Yi, L.; Dalai, M.; Su, R.; Lin, W.; Erdenedalai, M.; Luvsantseren, B.; Chimedtseren, C.; Wang, Z.; Hasi, S. Whole-Genome Sequencing of Wild Siberian Musk Deer (Moschus Moschiferus) Provides Insights into Its Genetic Features. BMC Genom. 2020, 21, 108. [Google Scholar] [CrossRef] [PubMed]
- Ming, Y.; Jian, J.; Yu, F.; Yu, X.; Wang, J.; Liu, W. Molecular Footprints of Inshore Aquatic Adaptation in Indo-Pacific Humpback Dolphin (Sousa Chinensis). Genomics 2019, 111, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Moskalev, A.A.; Kudryavtseva, A.V.; Graphodatsky, A.S.; Beklemisheva, V.R.; Serdyukova, N.A.; Krutovsky, K.V.; Sharov, V.V.; Kulakovskiy, I.V.; Lando, A.S.; Kasianov, A.S.; et al. De Novo Assembling and Primary Analysis of Genome and Transcriptome of Gray Whale Eschrichtius Robustus. BMC Evol. Biol. 2017, 17, 258. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; An, Y.R.; Kanda, N.; An, C.M.; An, H.S.; Kang, J.H.; Kim, E.M.; An, D.H.; Jung, H.; Joung, M.; et al. Cetaceans Evolution: Insights from the Genome Sequences of Common Minke Whales. BMC Genom. 2015, 16, 13. [Google Scholar] [CrossRef] [PubMed]
- Yim, H.S.; Cho, Y.S.; Guang, X.; Kang, S.G.; Jeong, J.Y.; Cha, S.-S.; Oh, H.-M.; Lee, J.-H.; Yang, E.C.; Kwon, K.K.; et al. Minke Whale Genome and Aquatic Adaptation in Cetaceans. Nat. Genet. 2014, 46, 88–92. [Google Scholar] [CrossRef]
- Whitehead, A.; Pilcher, W.; Champlin, D.; Nacci, D. Common Mechanism Underlies Repeated Evolution of Extreme Pollution Tolerance. Proc. R. Soc. B Biol. Sci. 2012, 279, 427–433. [Google Scholar] [CrossRef]
- Kim, E.S.; Elbeltagy, A.R.; Aboul-Naga, A.M.; Rischkowsky, B.; Sayre, B.; Mwacharo, J.M.; Rothschild, M.F. Multiple Genomic Signatures of Selection in Goats and Sheep Indigenous to a Hot Arid Environment. Heredity 2016, 116, 255–264. [Google Scholar] [CrossRef]
- Watson, M.; Holman, D.M.; Maguire-Eisen, M. Ultraviolet Radiation Exposure and Its Impact on Skin Cancer Risk. Semin. Oncol. Nurs. 2016, 32, 241–254. [Google Scholar] [CrossRef]
- Mwacharo, J.M.; Kim, E.S.; Elbeltagy, A.R.; Aboul-Naga, A.M.; Rischkowsky, B.A.; Rothschild, M.F. Genomic Footprints of Dryland Stress Adaptation in Egyptian Fat-Tail Sheep and Their Divergence from East African and Western Asia Cohorts. Sci. Rep. 2017, 7, 17647. [Google Scholar] [CrossRef]
- Chebii, V.J.; Oyola, S.O.; Kotze, A.; Domelevo Entfellner, J.-B.; Musembi Mutuku, J.; Agaba, M. Genome-Wide Analysis of Nubian Ibex Reveals Candidate Positively Selected Genes That Contribute to Its Adaptation to the Desert Environment. Animals 2020, 10, 2181. [Google Scholar] [CrossRef]
- Tunaru, S.; Chennupati, R.; Nüsing, R.M.; Offermanns, S. Arachidonic Acid Metabolite 19(S)-HETE Induces Vasorelaxation and Platelet Inhibition by Activating Prostacyclin (IP) Receptor. PLoS ONE 2016, 11, e0163633. [Google Scholar] [CrossRef] [PubMed]
- Blumthaler, M.; Ambach, W.; Ellinger, R. Increase in Solar UV Radiation with Altitude. J. Photochem. Photobiol. B Biol. 1997, 39, 130–134. [Google Scholar] [CrossRef]
- Gou, W.; Peng, J.; Wu, Q.; Zhang, Q.; Zhang, H.; Wu, C. Expression Pattern of Heme Oxygenase 1 Gene and Hypoxic Adaptation in Chicken Embryos. Comp. Biochem. Physiol. Part. B Biochem. Mol. Biol. 2014, 174, 23–28. [Google Scholar] [CrossRef]
- Li, L.L.; Ma, S.K.; Peng, W.; Fang, Y.G.; Fu, H.Y.; Jia, G.X. Whole Genome Sequencing of 5 Tibetan Sheep Breeds Identifies Selective Signatures to Adaptability at Different High-Altitude Areas in Qinghai-Tibetan Plateau. bioRxiv 2020. [Google Scholar] [CrossRef]
- Ma, Y.F.; Han, X.M.; Huang, C.P.; Zhong, L.; Adeola, A.C.; Irwin, D.M.; Xie, H.B.; Zhang, Y.P. Population Genomics Analysis Revealed Origin and High-Altitude Adaptation of Tibetan Pigs. Sci. Rep. 2019, 9, 11463. [Google Scholar] [CrossRef]
- Wei, C.; Wang, H.; Liu, G.; Zhao, F.; Kijas, J.W.; Ma, Y.; Lu, J.; Zhang, L.; Cao, J.; Wu, M.; et al. Genome-Wide Analysis Reveals Adaptation to High Altitudes in Tibetan Sheep. Sci. Rep. 2016, 6, 26770. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Li, Y.; Pan, J.; Wang, D.; Chen, W.; Zheng, Z.; He, X.; Zhao, Q.; Pu, Y.; et al. EPAS1 Gain-of-Function Mutation Contributes to High-Altitude Adaptation in Tibetan Horses. Mol. Biol. Evol. 2019, 36, 2591–2603. [Google Scholar] [CrossRef]
- Xiao, W.Y.; Ding, X.Z.; Chu, M.; Guo, X.; Bao, P.J.; Liang, C.N.; Yan, P. Novel SNP of EPAS1 Gene Associated with Higher Hemoglobin Concentration Revealed the Hypoxia Adaptation of Yak (Bos Grunniens). J. Integr. Agric. 2015, 14, 741–748. [Google Scholar] [CrossRef]
- Wu, D.-D.; Yang, C.-P.; Wang, M.-S.; Dong, K.-Z.; Yan, D.-W.; Hao, Z.-Q.; Fan, S.-Q.; Chu, S.-Z.; Shen, Q.-S.; Jiang, L.-P. Convergent Genomic Signatures of High-Altitude Adaptation among Domestic Mammals. Natl. Sci. Rev. 2020, 7, 952–963. [Google Scholar] [CrossRef]
- Iqbal, N.; Liu, X.; Yang, T.; Huang, Z.; Hanif, Q.; Asif, M.; Khan, Q.M.; Mansoor, S. Genomic Variants Identified from Whole-Genome Resequencing of Indicine Cattle Breeds from Pakistan. PLoS ONE 2019, 14, e0215065. [Google Scholar] [CrossRef]
- Palomera, S.Z.; Bucio-Mendez, A.; Valadez-Graham, V.; Reynaud, E.; Zurita, M. Drosophila P53 Is Required to Increase the Levels of the DKDM4B Demethylase after UV-Induced DNA Damage to Demethylate Histone H3 Lysine 9. J. Biol. Chem. 2010, 285, 31370–31379. [Google Scholar] [CrossRef]
- Shao, B.; Long, R.; Ding, Y.; Wang, J.; Ding, L.; Wang, H. Morphological Adaptations of Yak (Bos Grunniens) Tongue to the Foraging Environment of the Qinghai-Tibetan Plateau1. J. Anim. Sci. 2010, 88, 2594–2603. [Google Scholar] [CrossRef]
- Gorkhali, N.A.; Dong, K.; Yang, M.; Song, S.; Kader, A.; Shrestha, B.S.; He, X.; Zhao, Q.; Pu, Y.; Li, X.; et al. Genomic Analysis Identified a Potential Novel Molecular Mechanism for High-Altitude Adaptation in Sheep at the Himalayas. Sci. Rep. 2016, 6, 29963. [Google Scholar] [CrossRef]
- Ishengoma, E.; Agaba, M. Evolution of Toll-like Receptors in the Context of Terrestrial Ungulates and Cetaceans Diversification. BMC Evol. Biol. 2017, 17, 54. [Google Scholar] [CrossRef]
- Ndeereh, D.; Obanda, V.; Mijele, D.; Gakuya, F. Medicine in the Wild: Strategies towards Healthy and Breeding Wildlife Populations in Kenya. JSTOR 2012, 29, 100–108. [Google Scholar]
- Dimitrova, M.; Merilaita, S. Prey Concealment: Visual Background Complexity and Prey Contrast Distribution. Behav. Ecol. 2010, 21, 176–181. [Google Scholar] [CrossRef]
- Mitchell, G.; Roberts, D.G.; van Sittert, S.J.; Skinner, J.D. Orbit Orientation and Eye Morphometrics in Giraffes (Giraffa Camelopardalis). Afr. Zool. 2013, 48, 333–339. [Google Scholar] [CrossRef][Green Version]
- Shen, T.; Xu, S.; Wang, X.; Yu, W.; Zhou, K.; Yang, G. Adaptive Evolution and Functional Constraint at TLR4 during the Secondary Aquatic Adaptation and Diversification of Cetaceans. BMC Evol. Biol. 2012, 12, 39. [Google Scholar] [CrossRef] [PubMed]
- Huelsmann, M.; Hecker, N.; Springer, M.S.; Gatesy, J.; Sharma, V.; Hiller, M. Genes Lost during the Transition from Land to Water in Cetaceans Highlight Genomic Changes Associated with Aquatic Adaptations. Sci. Adv. 2019, 5, eaaw6671. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Wei, H.; Bi, J.; Ding, X.; Li, L.; Xu, S.; Yang, G.; Ren, W. Insights into Dietary Switch in Cetaceans: Evidence from Molecular Evolution of Proteinases and Lipases. J. Mol. Evol. 2020, 88, 521–535. [Google Scholar] [CrossRef] [PubMed]
- Croll, D.A.; Acevedo-Gutiérrez, A.; Tershy, B.R.; Urbán-Ramírez, J. The Diving Behavior of Blue and Fin Whales: Is Dive Duration Shorter than Expected Based on Oxygen Stores? Comp. Biochem. Physiol. Part. A: Mol. Integr. Physiol. 2001, 129, 797–809. [Google Scholar] [CrossRef]
- Söhle, J.; Machuy, N.; Smailbegovic, E.; Holtzmann, U.; Grönniger, E.; Wenck, H.; Stäb, F.; Winnefeld, M. Identification of New Genes Involved in Human Adipogenesis and Fat Storage. PLoS ONE 2012, 7, e31193. [Google Scholar] [CrossRef] [PubMed]
- Gonchar, O.; Mankovska, I. Antioxidant System in Adaptation to Intermittent Hypoxia. J. Biol. Sci. 2010, 10, 545–554. [Google Scholar] [CrossRef][Green Version]
- Tian, R.; Seim, I.; Ren, W.; Xu, S.; Yang, G. Comparative Genomics Reveals Contraction in Cytosolic Glutathione Transferase Genes in Cetaceans: Implications for Oxidative Stress Adaptation. bioRxiv 2018, 485615. [Google Scholar] [CrossRef]
- Tian, R.; Wang, Z.; Niu, X.; Zhou, K.; Xu, S.; Yang, G. Evolutionary Genetics of Hypoxia Tolerance in Cetaceans during Diving. Genome Biol. Evol. 2016, 8, 827–839. [Google Scholar] [CrossRef]
- Albalat, R.; Canestro, C. Evolution by Gene Loss. Nat. Rev. Genet. 2016, 17, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Hecker, N.; Roscito, J.G.; Foerster, L.; Langer, B.E.; Hiller, M. A Genomics Approach Reveals Insights into the Importance of Gene Losses for Mammalian Adaptations. Nat. Commun. 2018, 9, 1215. [Google Scholar] [CrossRef]
- Lyamin, O.I.; Manger, P.R.; Ridgway, S.H.; Mukhametov, L.M.; Siegel, J.M. Cetacean Sleep: An Unusual Form of Mammalian Sleep. Spec. Sect. Eur. Workshop Imag. Cogn. Neurocognition Vis. Imag. 2008, 32, 1451–1484. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Yang, Y.; Zhou, X.; Xu, J.; Zhou, K.; Yang, G. Adaptive Evolution of the Osmoregulation-Related Genes in Cetaceans during Secondary Aquatic Adaptation. BMC Evol. Biol. 2013, 13, 189. [Google Scholar] [CrossRef]
- São Pedro, S.L.; Alves, J.M.P.; Barreto, A.S.; de Lima, A.O.S. Evidence of Positive Selection of Aquaporins Genes from Pontoporia Blainvillei during the Evolutionary Process of Cetaceans. PLoS ONE 2015, 10, e0134516. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, X.; Hu, B.; Zheng, J.; Xiao, W.; Hao, Y.; Liu, W.; Wang, D. Physicochemical Evolution and Molecular Adaptation of the Cetacean Osmoregulation-Related Gene UT-A2 and Implications for Functional Studies. Sci. Rep. 2015, 5, 8795. [Google Scholar] [CrossRef] [PubMed]
- Blix, A.S. Arctic Animals and Their Adaptations to Life on the Edge; Tapir Academic Press: Cambridge, MA, USA, 2005. [Google Scholar]
- Alberto, F.J.; Boyer, F.; Orozco-terWengel, P.; Streeter, I.; Servin, B.; de Villemereuil, P.; Benjelloun, B.; Librado, P.; Biscarini, F.; Colli, L.; et al. Convergent Genomic Signatures of Domestication in Sheep and Goats. Nat. Commun. 2018, 9, 813. [Google Scholar] [CrossRef]
- Ghoreishifar, S.M.; Eriksson, S.; Johansson, A.M.; Khansefid, M.; Moghaddaszadeh-Ahrabi, S.; Parna, N.; Davoudi, P.; Javanmard, A. Signatures of Selection Reveal Candidate Genes Involved in Economic Traits and Cold Acclimation in Five Swedish Cattle Breeds. Genet. Sel. Evol. 2020, 52, 52. [Google Scholar] [CrossRef]
- Li, X.; Yang, J.; Shen, M.; Xie, X.-L.; Liu, G.-J.; Xu, Y.-X.; Lv, F.-H.; Yang, H.; Yang, Y.-L.; Liu, C.-B. Whole-Genome Resequencing of Wild and Domestic Sheep Identifies Genes Associated with Morphological and Agronomic Traits. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- McRae, K.M.; McEwan, J.C.; Dodds, K.G.; Gemmell, N.J. Signatures of Selection in Sheep Bred for Resistance or Susceptibility to Gastrointestinal Nematodes. BMC Genom. 2014, 15, 637. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Li, S.; Liu, Q.; Wang, Z.; Zhou, Z.; Di, R.; Miao, B.; Hu, W.; Wang, X.; Hu, X. Whole-Genome Sequences of 89 Chinese Sheep Suggest Role of RXFP2 in the Development of Unique Horn Phenotype as Response to Semi-Feralization. GigaScience 2018, 7, giy019. [Google Scholar] [CrossRef]
- Wiener, P.; Robert, C.; Ahbara, A.; Salavati, M.; Abebe, A.; Kebede, A.; Wragg, D.; Friedrich, J.; Vasoya, D.; Hume, D.A.; et al. Whole-Genome Sequence Data Suggest Environmental Adaptation of Ethiopian Sheep Populations. Genome Biol. Evol. 2021, 13. [Google Scholar] [CrossRef]
- Yang, L.; Xu, L.; Zhou, Y.; Liu, M.; Wang, L.; Kijas, J.W.; Zhang, H.; Li, L.; Liu, G.E. Diversity of Copy Number Variation in a Worldwide Population of Sheep. Genomics 2018, 110, 143–148. [Google Scholar] [CrossRef]
- Bertolini, F.; Servin, B.; Talenti, A.; Rochat, E.; Kim, E.S.; Oget, C.; Palhière, I.; Crisà, A.; Catillo, G.; Steri, R.; et al. Signatures of Selection and Environmental Adaptation across the Goat Genome Post-Domestication. Genet. Sel. Evol. 2018, 50, 57. [Google Scholar] [CrossRef]
- Liu, M.; Zhou, Y.; Rosen, B.D.; Van Tassell, C.P.; Stella, A.; Tosser-Klopp, G.; Rupp, R.; Palhière, I.; Colli, L.; Sayre, B.; et al. Diversity of Copy Number Variation in the Worldwide Goat Population. Heredity 2019, 122, 636–646. [Google Scholar] [CrossRef] [PubMed]
- Onzima, R.B.; Upadhyay, M.R.; Doekes, H.P.; Brito, L.F.; Bosse, M.; Kanis, E.; Groenen, M.A.M.; Crooijmans, R.P.M.A. Genome-Wide Characterization of Selection Signatures and Runs of Homozygosity in Ugandan Goat Breeds. Front. Genet. 2018, 9, 318. [Google Scholar] [CrossRef]
- Ben-Jemaa, S.; Mastrangelo, S.; Lee, S.-H.; Lee, J.H.; Boussaha, M. Genome-Wide Scan for Selection Signatures Reveals Novel Insights into the Adaptive Capacity in Local North African Cattle. Sci. Rep. 2020, 10, 19466. [Google Scholar] [CrossRef]
- Bickhart, D.M.; Hou, Y.; Schroeder, S.G.; Alkan, C.; Cardone, M.F.; Matukumalli, L.K.; Song, J.; Schnabel, R.D.; Ventura, M.; Taylor, J.F. Copy Number Variation of Individual Cattle Genomes Using Next-Generation Sequencing. Genome Res. 2012, 22, 778–790. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.E.; Hou, Y.; Zhu, B.; Cardone, M.F.; Jiang, L.; Cellamare, A.; Mitra, A.; Alexander, L.J.; Coutinho, L.L.; Dell’Aquila, M.E.; et al. Analysis of Copy Number Variations among Diverse Cattle Breeds. Genome Res. 2010, 20, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Pitt, D.; Bruford, M.W.; Barbato, M.; Orozco-terWengel, P.; Martínez, R.; Sevane, N. Demography and Rapid Local Adaptation Shape Creole Cattle Genome Diversity in the Tropics. Evol. Appl. 2019, 12, 105–122. [Google Scholar] [CrossRef]
- Diao, S.; Luo, Y.; Xi, D.; He, Y.; Ning, G.; Zhang, H.; Li, J.; Chen, Z.; Zhang, Z. Genome-Wide Detection of Selective Signatures in a Duroc Pig Population. J. Integr. Agric. 2018, 17, 2528–2535. [Google Scholar] [CrossRef]
- Groenen, M.A.M. A Decade of Pig Genome Sequencing: A Window on Pig Domestication and Evolution. Genet. Sel. Evol. 2016, 48, 23. [Google Scholar] [CrossRef]
- Groenen, M.A.M.; Archibald, A.L.; Uenishi, H.; Tuggle, C.K.; Takeuchi, Y.; Rothschild, M.F.; Rogel-Gaillard, C.; Park, C.; Milan, D.; Megens, H.J.; et al. Analyses of Pig Genomes Provide Insight into Porcine Demography and Evolution. Nature 2012, 491, 393–398. [Google Scholar] [CrossRef]
- Paudel, Y.; Madsen, O.; Megens, H.-J.; Frantz, L.A.; Bosse, M.; Crooijmans, R.P.; Groenen, M.A. Copy Number Variation in the Speciation of Pigs: A Possible Prominent Role for Olfactory Receptors. BMC Genom. 2015, 16, 330. [Google Scholar] [CrossRef]
- Rubin, C.J.; Megens, H.J.; Barrio, A.M.; Maqbool, K.; Sayyab, S.; Schwochow, D.; Wang, C.; Carlborg, O.; Jern, P.; Jørgensen, C.B.; et al. Strong Signatures of Selection in the Domestic Pig Genome. Proc. Natl. Acad. Sci. USA 2012, 109, 19529. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, C.; Chen, J.; Bai, Y.; Wang, K.; Wang, Y.; Fang, M. Genome-Wide Analysis Reveals Human-Mediated Introgression from Western Pigs to Indigenous Chinese Breeds. Genes 2020, 11, 275. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Fan, Y.; Wang, G.; Lai, Z.; Gao, Y.; Wu, F.; Lei, C.; Dang, R. Detection of Selection Signatures Underlying Production and Adaptive Traits Based on Whole-Genome Sequencing of Six Donkey Populations. Animals 2020, 10, 1823. [Google Scholar] [CrossRef]
- Petersen, J.L.; Mickelson, J.R.; Rendahl, A.K.; Valberg, S.J.; Andersson, L.S.; Axelsson, J.; Bailey, E.; Bannasch, D.; Binns, M.M.; Borges, A.S.; et al. Genome-Wide Analysis Reveals Selection for Important Traits in Domestic Horse Breeds. PLoS Genet. 2013, 9, e1003211. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.; Jónsson, H.; Chang, D.; Der Sarkissian, C.; Ermini, L.; Ginolhac, A.; Albrechtsen, A.; Dupanloup, I.; Foucal, A.; Petersen, B.; et al. Prehistoric Genomes Reveal the Genetic Foundation and Cost of Horse Domestication. Proc. Natl. Acad. Sci. USA 2014, 111, E5661. [Google Scholar] [CrossRef]
- Goshu, H.A.; Chu, M.; Yan, P. Applications of Genomic Copy Number Variations on Livestock: A Review. Afr. J. Biotechnol. 2018, 17, 1313–1323. [Google Scholar]
- Bickhart, D.M.; Xu, L.; Hutchison, J.L.; Cole, J.B.; Null, D.J.; Schroeder, S.G.; Song, J.; Garcia, J.F.; Sonstegard, T.S.; Van Tassell, C.P.; et al. Diversity and Population-Genetic Properties of Copy Number Variations and Multicopy Genes in Cattle. DNA Res. 2016, 23, 253–262. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, K.; Wang, L.; Yang, Y.; Ni, Z.; Xie, X.; Shao, X.; Han, J.; Wan, D.; Qiu, Q. Genome-Wide Patterns of Copy Number Variation in the Chinese Yak Genome. BMC Genom. 2016, 17, 379. [Google Scholar] [CrossRef]
- Paudel, Y.; Madsen, O.; Megens, H.-J.; Frantz, L.A.; Bosse, M.; Bastiaansen, J.W.; Crooijmans, R.P.; Groenen, M.A. Evolutionary Dynamics of Copy Number Variation in Pig Genomes in the Context of Adaptation and Domestication. BMC Genom. 2013, 14, 449. [Google Scholar] [CrossRef]
- Molnár, J.; Nagy, T.; Stéger, S.; Tóth, G.; Marincs, F.; Barta, E. Genome Sequencing and Analysis of Mangalica, a Fatty Local Pig of Hungary. BMC Genom. 2014, 15, 761. [Google Scholar] [CrossRef]
- Cohen-Zinder, M.; Seroussi, E.; Larkin, D.M.; Loor, J.J.; Everts-van der Wind, A.; Lee, J.-H.; Drackley, J.K.; Band, M.R.; Hernandez, A.G.; Shani, M.; et al. Identification of a Missense Mutation in the Bovine ABCG2 Gene with a Major Effect on the QTL on Chromosome 6 Affecting Milk Yield and Composition in Holstein Cattle. Genome Res. 2005, 15, 936–944. [Google Scholar] [CrossRef]
- Árnyasi, M.; Komlósi, I.; Kent, M.P.; Czeglédi, L.; Gulyás, G.; Jávor, A. Investigation of Polymorphisms and Association of the ABCG2 Gene with Milk Production Traits in Sheep. Livest. Sci. 2013, 154, 64–68. [Google Scholar] [CrossRef]
- Cieslak, M.; Reissmann, M.; Hofreiter, M.; Ludwig, A. Colours of Domestication. Biol. Rev. 2011, 86, 885–899. [Google Scholar] [CrossRef] [PubMed]
- Fontanesi, L.; Martelli, P.L.; Beretti, F.; Riggio, V.; Dall’Olio, S.; Colombo, M.; Casadio, R.; Russo, V.; Portolano, B. An Initial Comparative Map of Copy Number Variations in the Goat (Capra Hircus) Genome. BMC Genom. 2010, 11, 639. [Google Scholar] [CrossRef] [PubMed]
- Bickhart, D.M.; Liu, G. The Challenges and Importance of Structural Variation Detection in Livestock. Front. Genet. 2014, 5, 37. [Google Scholar] [CrossRef] [PubMed]
- Han, J.L.; Yang, M.; Yue, Y.J.; Guo, T.T.; Liu, J.B.; Niu, C.E.; Yang, B.H. Analysis of Agouti Signaling Protein (ASIP) Gene Polymorphisms and Association with Coat Color in Tibetan Sheep (Ovis Aries). Genet. Mol. Res. 2015, 14, 1200–1209. [Google Scholar] [CrossRef]
- Xu, L.; Bickhart, D.M.; Cole, J.B.; Schroeder, S.G.; Song, J.; Tassell, C.P.V.; Sonstegard, T.S.; Liu, G.E. Genomic Signatures Reveal New Evidences for Selection of Important Traits in Domestic Cattle. Mol. Biol. Evol. 2015, 32, 711–725. [Google Scholar] [CrossRef]
- Wang, C.; Wang, H.; Zhang, Y.; Tang, Z.; Li, K.; Liu, B. Genome-Wide Analysis Reveals Artificial Selection on Coat Colour and Reproductive Traits in Chinese Domestic Pigs. Mol. Ecol. Resour. 2015, 15, 414–424. [Google Scholar] [CrossRef]
- Gering, E.; Incorvaia, D.; Henriksen, R.; Wright, D.; Getty, T. Maladaptation in Feral and Domesticated Animals. Evol. Appl. 2019, 12, 1274–1286. [Google Scholar] [CrossRef]
- Qiu, Q.; Wang, L.; Wang, K.; Yang, Y.; Ma, T.; Wang, Z.; Zhang, X.; Ni, Z.; Hou, F.; Long, R.; et al. Yak Whole-Genome Resequencing Reveals Domestication Signatures and Prehistoric Population Expansions. Nat. Commun. 2015, 6, 10283. [Google Scholar] [CrossRef] [PubMed]
- Kawahara-Miki, R.; Tsuda, K.; Shiwa, Y.; Arai-Kichise, Y.; Matsumoto, T.; Kanesaki, Y.; Oda, S.; Ebihara, S.; Yajima, S.; Yoshikawa, H. Whole-Genome Resequencing Shows Numerous Genes with Nonsynonymous SNPs in the Japanese Native Cattle Kuchinoshima-Ushi. BMC Genom. 2011, 12, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mei, C.; Wang, H.; Liao, Q.; Khan, R.; Raza, S.H.A.; Zhao, C.; Wang, H.; Cheng, G.; Tian, W.; Li, Y. Genome-Wide Analysis Reveals the Effects of Artificial Selection on Production and Meat Quality Traits in Qinchuan Cattle. Genomics 2019, 111, 1201–1208. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, S.; Zhang, K.; Fang, C.; Xie, S.; Du, X.; Li, X.; Ni, D.; Zhao, S. Genomic Analysis to Identify Signatures of Artificial Selection and Loci Associated with Important Economic Traits in Duroc Pigs. G3: Genesgenomesgenetics 2018, 8, 3617–3625. [Google Scholar] [CrossRef]
- Suárez-Vega, A.; Gutiérrez-Gil, B.; Klopp, C.; Tosser-Klopp, G.; Arranz, J.J. Variant Discovery in the Sheep Milk Transcriptome Using RNA Sequencing. BMC Genom. 2017, 18, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.-X.; Yang, J.; Lv, F.-H.; Hu, X.-J.; Xie, X.-L.; Zhang, M.; Li, W.-R.; Liu, M.-J.; Wang, Y.-T.; Li, J.-Q. Genomic Reconstruction of the History of Native Sheep Reveals the Peopling Patterns of Nomads and the Expansion of Early Pastoralism in East Asia. Mol. Biol. Evol. 2017, 34, 2380–2395. [Google Scholar] [CrossRef] [PubMed]
- Frischknecht, M.; Bapst, B.; Seefried, F.R.; Signer-Hasler, H.; Garrick, D.; Stricker, C.; Fries, R.; Russ, I.; Sölkner, J.; Bieber, A. Genome-Wide Association Studies of Fertility and Calving Traits in Brown Swiss Cattle Using Imputed Whole-Genome Sequences. BMC Genom. 2017, 18, 1–13. [Google Scholar] [CrossRef]
- Rothammer, S.; Seichter, D.; Förster, M.; Medugorac, I. A Genome-Wide Scan for Signatures of Differential Artificial Selection in Ten Cattle Breeds. BMC Genom. 2013, 14, 1–17. [Google Scholar] [CrossRef]
- Cao, Y.-H.; Xu, S.-S.; Shen, M.; Chen, Z.-H.; Gao, L.; Lv, F.-H.; Xie, X.-L.; Wang, X.-H.; Yang, H.; Liu, C.-B. Historical Introgression from Wild Relatives Enhanced Climatic Adaptation and Resistance to Pneumonia in Sheep. Mol. Biol. Evol. 2021, 38, 838–855. [Google Scholar] [CrossRef]
- Hu, X.-J.; Yang, J.; Xie, X.-L.; Lv, F.-H.; Cao, Y.-H.; Li, W.-R.; Liu, M.-J.; Wang, Y.-T.; Li, J.-Q.; Liu, Y.-G. The Genome Landscape of Tibetan Sheep Reveals Adaptive Introgression from Argali and the History of Early Human Settlements on the Qinghai–Tibetan Plateau. Mol. Biol. Evol. 2019, 36, 283–303. [Google Scholar] [CrossRef] [PubMed]
- Barbato, M.; Hailer, F.; Upadhyay, M.; Del Corvo, M.; Colli, L.; Negrini, R.; Kim, E.-S.; Crooijmans, R.P.; Sonstegard, T.; Ajmone-Marsan, P. Adaptive Introgression from Indicine Cattle into White Cattle Breeds from Central Italy. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Liu, X.; Li, Z.; Yan, Y.; Li, Y.; Wu, H.; Pei, J.; Yan, P.; Yang, R.; Guo, X.; Lan, X. Selection and Introgression Facilitated the Adaptation of Chinese Native Endangered Cattle in Extreme Environments. Evol. Appl. 2021, 14, 860–873. [Google Scholar] [CrossRef]
- Fan, R.; Gu, Z.; Guang, X.; Marín, J.C.; Varas, V.; González, B.A.; Wheeler, J.C.; Hu, Y.; Li, E.; Sun, X. Genomic Analysis of the Domestication and Post-Spanish Conquest Evolution of the Llama and Alpaca. Genome Biol. 2020, 21, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Grossen, C.; Keller, L.; Biebach, I.; Croll, D. International Goat Genome Consortium Introgression from Domestic Goat Generated Variation at the Major Histocompatibility Complex of Alpine Ibex. PLoS Genet. 2014, 10, e1004438. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, S.L. A Genome Wide Study of Genetic Adaptation to High Altitude in Feral Andean Horses of the Páramo. BMC Evol. Biol. 2013, 13, 1–13. [Google Scholar] [CrossRef]
- Wang, X.; Liu, J.; Zhou, G.; Guo, J.; Yan, H.; Niu, Y.; Li, Y.; Yuan, C.; Geng, R.; Lan, X.; et al. Whole-Genome Sequencing of Eight Goat Populations for the Detection of Selection Signatures Underlying Production and Adaptive Traits. Sci. Rep. 2016, 6, 38932. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.; Yin, D.; Liu, Y.; Seim, I.; Xu, S.; Yang, G. Adaptive Evolution of Energy Metabolism-Related Genes in Hypoxia-Tolerant Mammals. Front. Gen. 2017, 8, 205. [Google Scholar] [CrossRef]
- Yang, S.; Li, X.; Li, K.; Fan, B.; Tang, Z. A Genome-Wide Scan for Signatures of Selection in Chinese Indigenous and Commercial Pig Breeds. BMC Genet. 2014, 15, 7. [Google Scholar] [CrossRef]
- Bosse, M.; Megens, H.J.; Madsen, O.; Frantz, L.A.F.; Paudel, Y.; Crooijmans, R.P.M.A.; Groenen, M.A.M. Untangling the Hybrid Nature of Modern Pig Genomes: A Mosaic Derived from Biogeographically Distinct and Highly Divergent Sus Scrofa Populations. Mol. Ecol. 2014, 23, 4089–4102. [Google Scholar] [CrossRef]
- Frantz, L.A.F.; Schraiber, J.G.; Madsen, O.; Megens, H.J.; Cagan, A.; Bosse, M.; Paudel, Y.; Crooijmans, R.P.M.A.; Larson, G.; Groenen, M.A.M. Evidence of Long-Term Gene Flow and Selection during Domestication from Analyses of Eurasian Wild and Domestic Pig Genomes. Nat. Genet. 2015, 47, 1141–1148. [Google Scholar] [CrossRef]

| Type of Signature | Detectable Pattern | Methodologies |
|---|---|---|
| Change allele frequency spectrum | Increased frequency of derived alleles | Tajima’s D [24] |
| Extended haplotype homozygosity | Linkage disequilibrium (LD) persistency and unusual long-range haplotypes | Cross-population extended haplotype homozygosity (XP-EHH) [23] |
| Integrated haplotype score (iHS) [23] | ||
| Population differentiation | Different allele frequencies between populations | FST [23] |
| Composite methods | Detects increased frequency of derived alleles, difference in allele frequencies and unusual long-range haplotypes | CMS [26] |
| Ungulate | Species | Genome Size (Gbp) | Number of Annotated Genes | GenBank Assembly Accession |
|---|---|---|---|---|
| Desert ungulates | Bactrian camel | 2.4 | 20,251 | AGVR01000000, JARL00000000 |
| Dromedary camel | 2.5 | 20,714 | JDVD00000000 | |
| Red deer | 3.3 | 22,138 | MKHE00000000 | |
| High-altitude ungulates | Wild yak | 2.6 | 22,282 | AGSK01000000 |
| Siberian musk deer | 3.1 | 19,363 | GCA_011751665.1 | |
| Artic ungulates | Rein deer | 2.9 | 27,332 | GCA_014898785.1 |
| Savanna | Giraffe | 2.9 | 17,210 | LVKQ00000000 |
| African buffalo | 2.7 | 19,765 | SAMN05717674 | |
| Marine | Blue whale | 2.4 | 19,518 | GCA_009873245.2 |
| Dolphin | 2.5 | 16,550 | GCA_011057625.1 | |
| Domesticated ungulates | Cow | 2.7 | 21,880 | GCA_002263795.2 |
| Sheep | 2.9 | 20,506 | GCA_002742125.1 | |
| Goat | 2.9 | 21,361 | GCA_001704415.1 | |
| Pig | 2.5 | 21,303 | GCA_000003025.6 | |
| Donkey | 2.3 | 19,963 | GCA_003033725.1 | |
| Horse | 2.5 | 20,955 | GCA_002863925.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chebii, V.J.; Mpolya, E.A.; Muchadeyi, F.C.; Domelevo Entfellner, J.-B. Genomics of Adaptations in Ungulates. Animals 2021, 11, 1617. https://doi.org/10.3390/ani11061617
Chebii VJ, Mpolya EA, Muchadeyi FC, Domelevo Entfellner J-B. Genomics of Adaptations in Ungulates. Animals. 2021; 11(6):1617. https://doi.org/10.3390/ani11061617
Chicago/Turabian StyleChebii, Vivien J., Emmanuel A. Mpolya, Farai C. Muchadeyi, and Jean-Baka Domelevo Entfellner. 2021. "Genomics of Adaptations in Ungulates" Animals 11, no. 6: 1617. https://doi.org/10.3390/ani11061617
APA StyleChebii, V. J., Mpolya, E. A., Muchadeyi, F. C., & Domelevo Entfellner, J.-B. (2021). Genomics of Adaptations in Ungulates. Animals, 11(6), 1617. https://doi.org/10.3390/ani11061617