Good to Know: Baseline Data on Feed Intake, Fecal Pellet Output and Intestinal Transit Time in Guinea Pig as a Frequently Used Model in Gastrointestinal Research


{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Animals
2.2. Experiment 1: 24-h Feed Intake and Fecal Pellet Output in Single-Housed Guinea Pigs
2.3. Experiment 2: 24-h Feed Intake in Single-Housed and Pair-Housed Guinea Pigs
2.4. Experiment 3: Determination of Whole Intestinal Transit Time
2.5. Statistics
3. Results
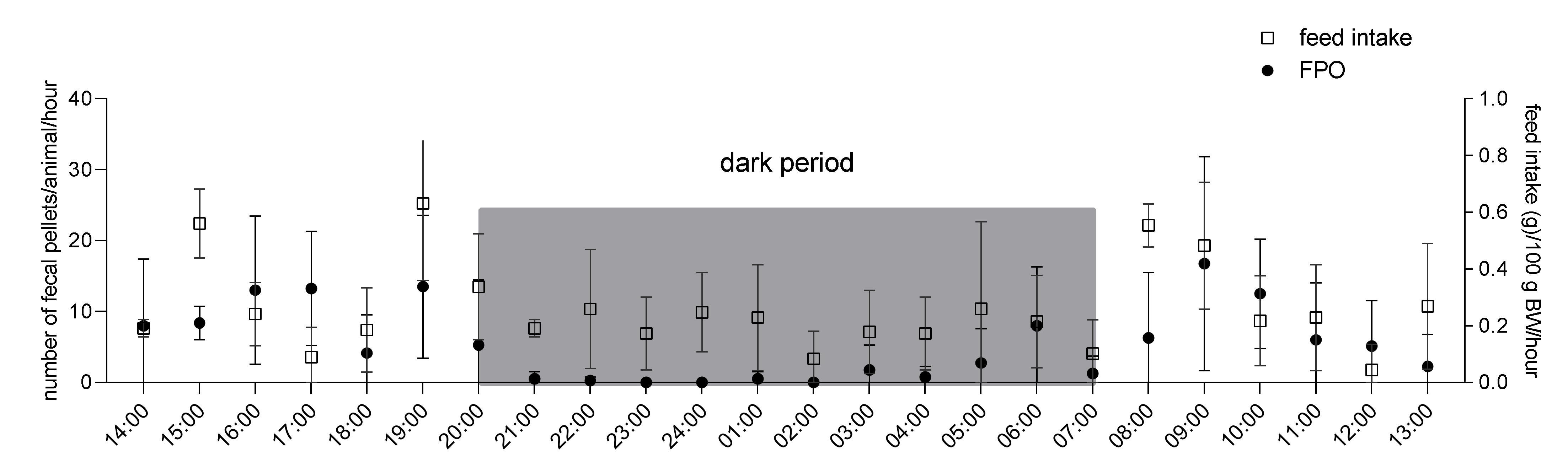
3.1. Experiment 1: 24-h Feed Intake and Fecal Pellet Output in Single-Housed Guinea Pigs
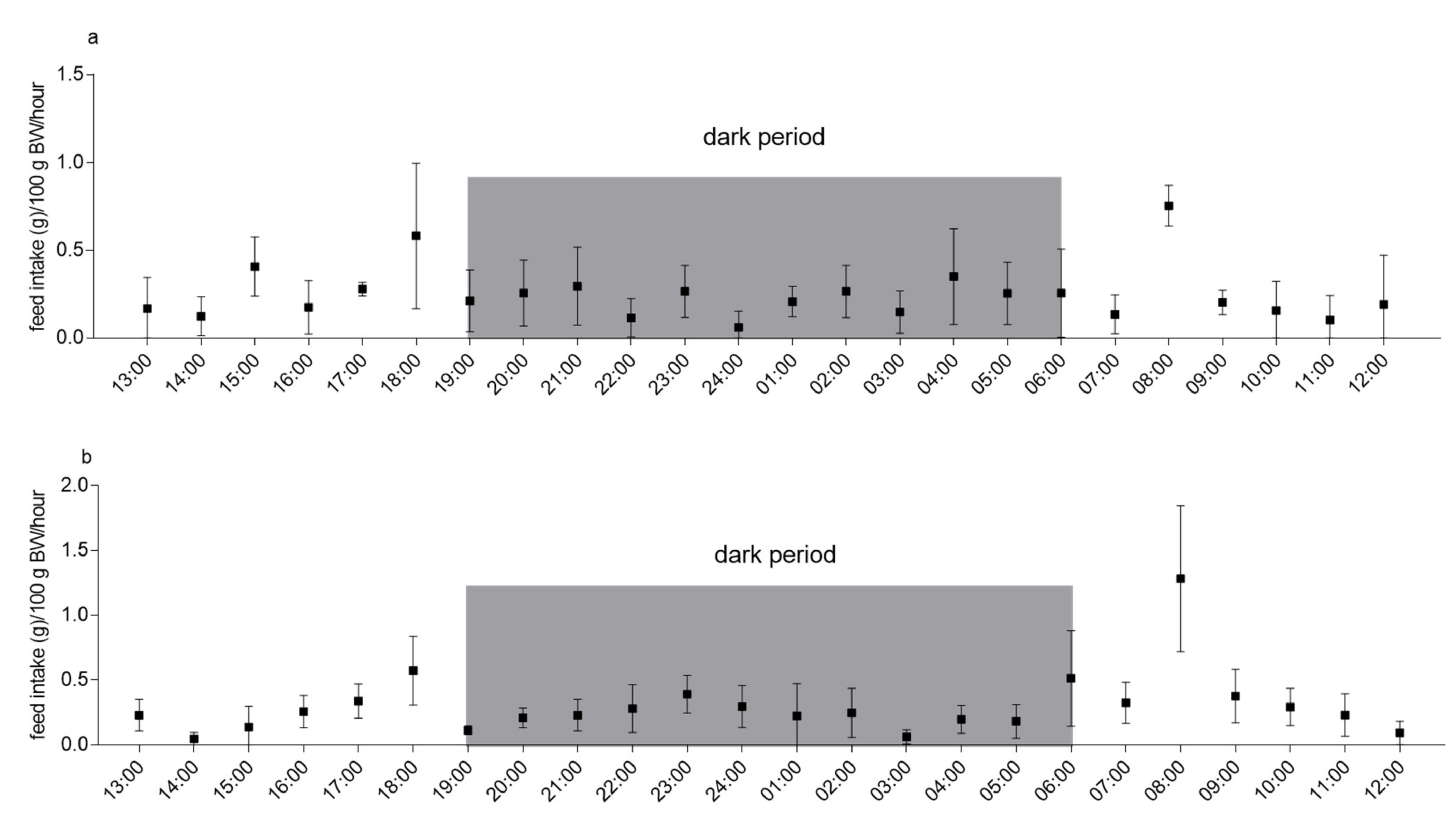
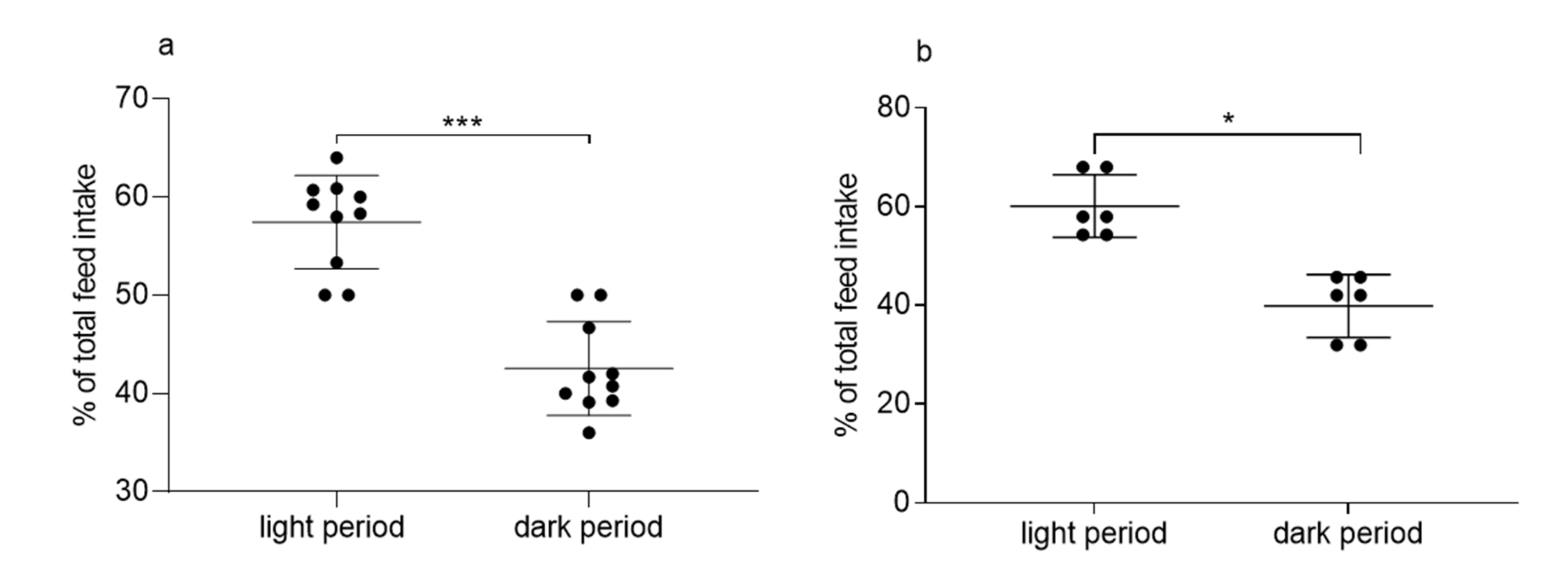
3.2. Experiment 2: 24-h Feed Intake in Single-Housed and Pair-Housed Guinea Pigs
3.3. Experiment 3: Whole Intestinal Transit Time
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harkness, J.E.; Murray, K.A.; Wagner, J.E. Biology and diseases of guinea pigs. In Laboratory Animal Medicine; 2007/09/02 ed.; Fox, J.G., Anderson, L.C., Loew, F.M., Quimby, F.W., Eds.; Elsevier: Amsterdam, The Netherlands, 2002; pp. 203–246. [Google Scholar]
- Brookes, S.J. Classes of enteric nerve cells in the guinea-pig small intestine. Anat. Rec. 2001, 262, 58–70. [Google Scholar] [CrossRef]
- Merchant, H.A.; McConnell, E.L.; Liu, F.; Ramaswamy, C.; Kulkarni, R.P.; Basit, A.W.; Murdan, S. Assessment of gastrointestinal pH, fluid and lymphoid tissue in the guinea pig, rabbit and pig, and implications for their use in drug development. Eur. J. Pharm. Sci. 2011, 42, 3–10. [Google Scholar] [CrossRef]
- Hildebrand, F.; Ebersbach, T.; Nielsen, H.B.; Li, X.; Sonne, S.B.; Bertalan, M.; Dimitrov, P.; Madsen, L.; Qin, J.; Wang, J.; et al. A comparative analysis of the intestinal metagenomes present in guinea pigs (Cavia porcellus) and humans (Homo sapiens). BMC Genom. 2012, 13, 514. [Google Scholar] [CrossRef]
- Al-Saffar, A.; Takemi, S.; Saaed, H.K.; Sakata, I.; Sakai, T. Utility of animal gastrointestinal motility and transit models in functional gastrointestinal disorders. Best Pract. Res. Clin. Gastroenterol. 2019, 40–41, 101633. [Google Scholar] [CrossRef]
- Kararli, T.T. Comparison of the gastrointestinal anatomy, physiology, and biochemistry of humans and commonly used laboratory animals. Biopharm. Drug Dispos. 1995, 16, 351–380. [Google Scholar] [CrossRef]
- McGinnis, G.R.; Young, M.E. Circadian regulation of metabolic homeostasis: Causes and consequences. Nat. Sci. Sleep 2016, 8, 163–180. [Google Scholar] [CrossRef] [PubMed]
- Voigt, R.M.; Forsyth, C.B.; Keshavarzian, A. Circadian rhythms: A regulator of gastrointestinal health and dysfunction. Expert Rev. Gastroenterol. Hepatol. 2019, 13, 411–424. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, M.; Linden, D.R. Measurement of gastrointestinal and colonic motor functions in humans and animals. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 412–428. [Google Scholar] [CrossRef]
- Horton, B.J.; West, C.E.; Turley, S.D. Diurnal variation in the feeding pattern of guinea pigs. Nutr. Metab. 1975, 18, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, E. Some determinants of intake and patterns of feeding in the guinea pig. Physiol. Behav. 1973, 11, 687–704. [Google Scholar] [CrossRef]
- Hussain, Z.; Kim, H.W.; Huh, C.W.; Lee, Y.J.; Park, H. The effect of peripheral CRF peptide and water avoidance stress on colonic and gastric transit in guinea pigs. Yonsei Med. J. 2017, 58, 872–877. [Google Scholar] [CrossRef]
- Hussain, Z.; Jung, D.H.; Lee, Y.J.; Park, H. The effect of trimebutine on the overlap syndrome model of guinea pigs. J. Neurogastroenterol. Motil. 2018, 24, 669–675. [Google Scholar] [CrossRef]
- Hussain, Z.; Rhee, K.W.; Lee, Y.J.; Park, H. The effect of DA-9701 in opioid-induced bowel dysfunction of guinea pig. J. Neurogastroenterol. Motil. 2016, 22, 529–538. [Google Scholar] [CrossRef]
- Jeong, E.J.; Chung, S.Y.; Hong, H.N.; Oh, S.W.; Sim, J.Y. The novel, potent and highly selective 5-HT(4) receptor agonist YH12852 significantly improves both upper and lower gastrointestinal motility. Br. J. Pharmacol. 2018, 175, 485–500. [Google Scholar] [CrossRef]
- Park, J.J.; Chon, N.R.; Lee, Y.J.; Park, H. The effects of an extract of Atractylodes Japonica rhizome, SKI3246 on gastrointestinal motility in guinea pigs. J. Neurogastroenterol. Motil. 2015, 21, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.M.; Lee, Y.J.; Lee, Y.H.; Kim, T.I.; Park, H. Effects of ramosetron on gastrointestinal transit of guinea pig. J. Neurogastroenterol. Motil. 2013, 19, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Jilge, B. The gastrointestinal transit time in the guinea-pig. Z. Versuchstierkd. 1980, 22, 204–210. [Google Scholar]
- Chiou, P.W.-S.; Yu, B.; Kuo, C.-Y. Comparison of digestive function among rabbits, guinea-pigs, rats and hamsters. I. Performance, digestibility and rate of digesta passage. Asian Australas J. Anim. Sci. 2000, 13, 1499–1507. [Google Scholar] [CrossRef]
- Sakaguchi, E.; Itoh, H.; Uchida, S.; Horigome, T. Comparison of fibre digestion and digesta retention time between rabbits, guinea-pigs, rats and hamsters. Br. J. Nutr. 1987, 58, 149–158. [Google Scholar] [CrossRef]
- Sakaguchi, E.; Ohmura, S. Fibre digestion and digesta retention time in guinea-pigs (Cavia porcellus), degus (Octodon degus) and leaf-eared mice (Phyllotis darwini). Comp. Biochem. Physiol. Comp. Physiol. 1992, 103, 787–791. [Google Scholar] [CrossRef]
- Cuomo, R.; Sarnelli, G. Food intake and gastrointestinal motility. A complex interplay. Nutr. Metab. Cardiovasc. Dis. 2004, 14, 173–179. [Google Scholar] [CrossRef]
- Ellacott, K.L.J.; Morton, G.J.; Woods, S.C.; Tso, P.; Schwartz, M.W. Assessment of feeding behavior in laboratory mice. Cell Metab. 2010, 12, 10–17. [Google Scholar] [CrossRef]
- Georgsson, L.; Barrett, J.; Gietzen, D. The effects of group-housing and relative weight on feeding behaviour in rats. Scand. J. Lab. Anim. Sci. 2001, 28, 201–209. [Google Scholar]
- Chvédoff, M.; Clarke, M.R.; Irisarri, E.; Faccini, J.M.; Monro, A.M. Effects of housing conditions on food intake, body weight and spontaneous lesions in mice. A review of the literature and results of an 18-month study. Food Cosmet. Toxicol. 1980, 18, 517–522. [Google Scholar] [CrossRef]
- Whary, M.; Peper, R.; Borkowski, G.; Lawrence, W.; Ferguson, F. The effects of group housing on the research use of the laboratory rabbit. Lab. Anim. 1993, 27, 330–341. [Google Scholar] [CrossRef]
- Horton, B.J.; Turley, S.D.; West, C.E. Diurnal variation in the feeding pattern of rabbits. Life Sci. 1974, 15, 1895–1907. [Google Scholar] [CrossRef]
- King, J.A. Social relations of the domestic guinea pig living under semi-natural conditions. Ecology 1956, 37, 221–228. [Google Scholar] [CrossRef]
- Rood, J.P. Ecological and behavioural comparisons of three genera of argentine cavies. Anim. Behav. Monogr. 1972, 5, 1–IN4. [Google Scholar] [CrossRef]
- Kurumiya, S.; Kawamura, H. Circadian oscillation of the multiple unit activity in the guinea pig suprachiasmatic nucleus. J. Comp. Physiol. A 1988, 162, 301–308. [Google Scholar] [CrossRef]
- White, W.; Lang, C. Use of cage space by guineapigs. Lab. Anim. 1989, 23, 208–214. [Google Scholar] [CrossRef]
- Nicholls, E.E. A study of the spontaneous activity of the guinea pig. J. Comp. Psychol. 1922, 2, 303–330. [Google Scholar] [CrossRef][Green Version]
- Lee, K.-N.; Pellom, S.T.; Oliver, E.; Chirwa, S. Characterization of the guinea pig animal model and subsequent comparison of the behavioral effects of selective dopaminergic drugs and methamphetamine. Synapse 2014, 68, 221–233. [Google Scholar] [CrossRef]
- Malone, J.C.; Thavamani, A. Physiology, Gastrocolic Reflex (Gastrocolic Response); StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Bove, G.M. A non-invasive method to evaluate gastrointestinal transit behavior in rat. J. Pharmacol. Toxicol. Methods 2015, 74, 1–6. [Google Scholar] [CrossRef][Green Version]
- Kenagy, G.J.; Veloso, C.; Bozinovic, F. Daily rhythms of food intake and feces reingestion in the degu, an herbivorous Chilean rodent: Optimizing digestion through coprophagy. Physiol. Biochem. Zool. 1999, 72, 78–86. [Google Scholar] [CrossRef]
- Ebino, K. Studies on coprophagy in experimental animals. Jikken Dobutsu 1993, 42, 1–9. [Google Scholar] [PubMed]
- Bass, P.; Kennedy, J.A.; Wiley, J.N. Measurement of fecal output in rats. Am. J. Dig. Dis. 1972, 17, 925–928. [Google Scholar] [CrossRef]
- Huang, H.H.; Ting, C.H.; Syu, Y.F.; Chang, S.C.; Chen, C.Y. Correlation between colonic secretion and colonic motility in rats: Role of ghrelin. World J. Gastroenterol. 2016, 22, 10140–10147. [Google Scholar] [CrossRef] [PubMed]
- Mönnikes, H.; Schmidt, B.G.; Taché, Y. Psychological stress-induced accelerated colonic transit in rats involves hypothalamic corticotropin-releasing factor. Gastroenterology 1993, 104, 716–723. [Google Scholar] [CrossRef]
- Kadowaki, M.; Nagakura, Y.; Tomoi, M.; Mori, J.; Kohsaka, M. Effect of FK1052, a potent 5-hydroxytryptamine3 and 5-hydroxytryptamine4 receptor dual antagonist, on colonic function in vivo. J. Pharmacol. Exp. Ther. 1993, 266, 74–80. [Google Scholar] [PubMed]
- Maillot, C.; Million, M.; Wei, J.Y.; Gauthier, A.; Taché, Y. Peripheral corticotropin-releasing factor and stress-stimulated colonic motor activity involve type 1 receptor in rats. Gastroenterology 2000, 119, 1569–1579. [Google Scholar] [CrossRef]
- Million, M.; Wang, L.; Martinez, V.; Taché, Y. Differential Fos expression in the paraventricular nucleus of the hypothalamus, sacral parasympathetic nucleus and colonic motor response to water avoidance stress in Fischer and Lewis rats. Brain Res. 2000, 877, 345–353. [Google Scholar] [CrossRef]
- Barone, F.C.; Deegan, J.F.; Price, W.J.; Fowler, P.J.; Fondacaro, J.D.; Ormsbee, H.S., 3rd. Cold-restraint stress increases rat fecal pellet output and colonic transit. Am. J. Physiol. 1990, 258, G329–G337. [Google Scholar] [CrossRef] [PubMed]
- Meredith, A.; Prebble, J. Impact of diet on faecal output and caecotroph consumption in rabbits: Rabbit faecal output and caecotroph consumption. J. Small Anim. Pract. 2017, 58, 139–145. [Google Scholar] [CrossRef]
- Rota, L.; Pellegrini, C.; Benvenuti, L.; Antonioli, L.; Fornai, M.; Blandizzi, C.; Cattaneo, A.; Colla, E. Constipation, deficit in colon contractions and alpha-synuclein inclusions within the colon precede motor abnormalities and neurodegeneration in the central nervous system in a mouse model of alpha-synucleinopathy. Transl. Neurodegener. 2019, 8, 5. [Google Scholar] [CrossRef]
- Nagakura, Y.; Naitoh, Y.; Kamato, T.; Yamano, M.; Miyata, K. Compounds possessing 5-HT3 receptor antagonistic activity inhibit intestinal propulsion in mice. Eur. J. Pharmacol. 1996, 311, 67–72. [Google Scholar] [CrossRef]
- Toure, A.M.; Charrier, B.; Pilon, N. Male-specific colon motility dysfunction in the TashT mouse line. Neurogastroenterol. Motil. 2016, 28, 1494–1507. [Google Scholar] [CrossRef]
- Spear, E.T.; Holt, E.A.; Joyce, E.J.; Haag, M.M.; Mawe, S.M.; Hennig, G.W.; Lavoie, B.; Applebee, A.M.; Teuscher, C.; Mawe, G.M. Altered gastrointestinal motility involving autoantibodies in the experimental autoimmune encephalomyelitis model of multiple sclerosis. Neurogastroenterol. Motil. 2018, 30, e13349. [Google Scholar] [CrossRef] [PubMed]
- Hoibian, E.; Florens, N.; Koppe, L.; Vidal, H.; Soulage, C.O. Distal Colon Motor Dysfunction in Mice with Chronic Kidney Disease: Putative Role of Uremic Toxins. Toxins 2018, 10, 204. [Google Scholar] [CrossRef] [PubMed]
- Asuzu, D.T.; Hayashi, Y.; Izbeki, F.; Popko, L.N.; Young, D.L.; Bardsley, M.R.; Lorincz, A.; Kuro, O.M.; Linden, D.R.; Farrugia, G.; et al. Generalized neuromuscular hypoplasia, reduced smooth muscle myosin and altered gut motility in the klotho model of premature aging. Neurogastroenterol. Motil. 2011, 23, e309–e323. [Google Scholar] [CrossRef][Green Version]
- Beattie, D.T.; Armstrong, S.R.; Shaw, J.P.; Marquess, D.; Sandlund, C.; Smith, J.A.M.; Taylor, J.A.; Humphrey, P.P.A. The in vivo gastrointestinal activity of TD-5108, a selective 5-HT4 receptor agonist with high intrinsic activity. Naunyn Schmiedeberg’s Arch. Pharmacol. 2008, 378, 139–147. [Google Scholar] [CrossRef]
- Beattie, D.T.; Armstrong, S.R.; Vickery, R.G.; Tsuruda, P.R.; Campbell, C.B.; Richardson, C.; McCullough, J.L.; Daniels, O.; Kersey, K.; Li, Y.P.; et al. The pharmacology of TD-8954, a potent and selective 5-HT(4) receptor agonist with gastrointestinal prokinetic properties. Front. Pharmacol. 2011, 2, 25. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, K.; Kitsukawa, Y.; Suzuki, Y.; Saito, Y. Effects of several endothelin receptor antagonists on gastrointestinal transit of guinea pigs. Dig. Dis. Sci. 2000, 45, 960–966. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elfers, K.; Armbrecht, Y.; Mazzuoli-Weber, G. Good to Know: Baseline Data on Feed Intake, Fecal Pellet Output and Intestinal Transit Time in Guinea Pig as a Frequently Used Model in Gastrointestinal Research. Animals 2021, 11, 1593. https://doi.org/10.3390/ani11061593
Elfers K, Armbrecht Y, Mazzuoli-Weber G. Good to Know: Baseline Data on Feed Intake, Fecal Pellet Output and Intestinal Transit Time in Guinea Pig as a Frequently Used Model in Gastrointestinal Research. Animals. 2021; 11(6):1593. https://doi.org/10.3390/ani11061593
Chicago/Turabian StyleElfers, Kristin, Yvonne Armbrecht, and Gemma Mazzuoli-Weber. 2021. "Good to Know: Baseline Data on Feed Intake, Fecal Pellet Output and Intestinal Transit Time in Guinea Pig as a Frequently Used Model in Gastrointestinal Research" Animals 11, no. 6: 1593. https://doi.org/10.3390/ani11061593
APA StyleElfers, K., Armbrecht, Y., & Mazzuoli-Weber, G. (2021). Good to Know: Baseline Data on Feed Intake, Fecal Pellet Output and Intestinal Transit Time in Guinea Pig as a Frequently Used Model in Gastrointestinal Research. Animals, 11(6), 1593. https://doi.org/10.3390/ani11061593