
Untargeted Metabolomics Reveals Intestinal Pathogenesis and Self-Repair in Rabbits Fed an Antibiotic-Free Diet

 , , and
, , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Farm Animal Sampling
2.2. Intestinal Tissue Sample Collection
2.3. Morphological Analysis of Intestinal Tissue
2.4. Metabolite Extraction
2.5. UHPLC–MS/MS Analysis
2.6. Data Processing and Metabolite Identification
2.7. Metabolite Analysis
3. Results
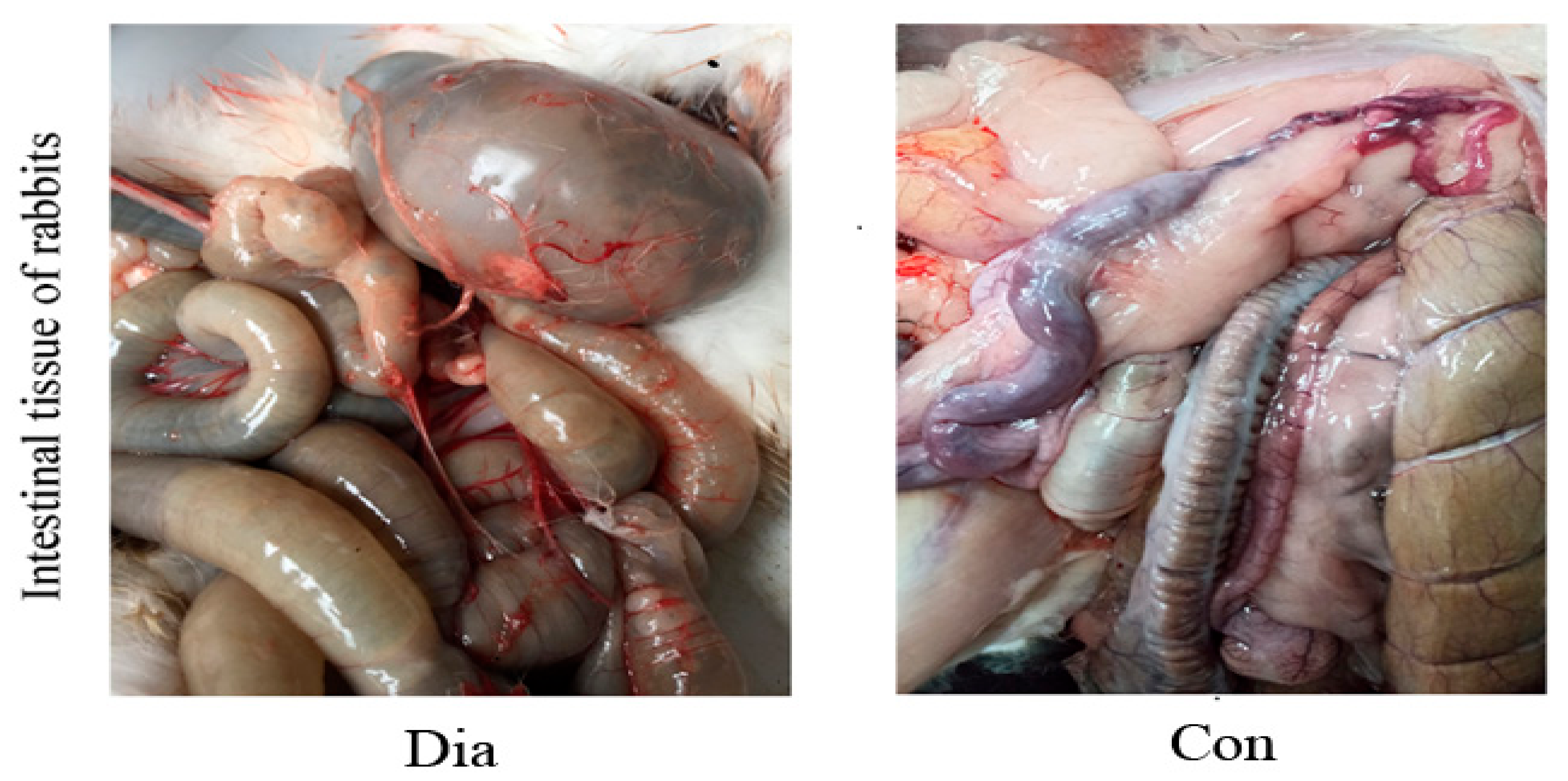
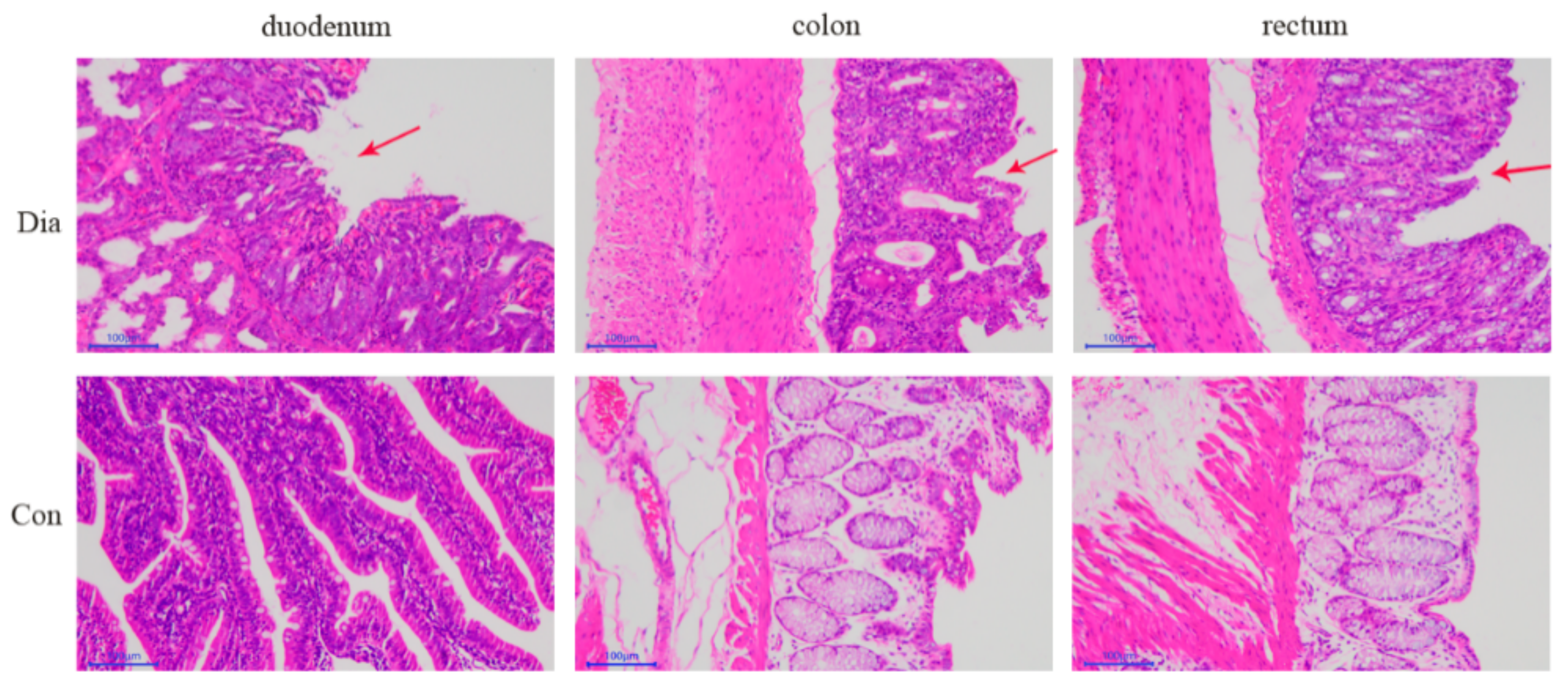
3.1. Intestinal Pathological Characteristics
3.2. Screening and Acquisition of Primitive Metabolites
3.3. Quality Control of Metabolite Data from Intestinal Samples
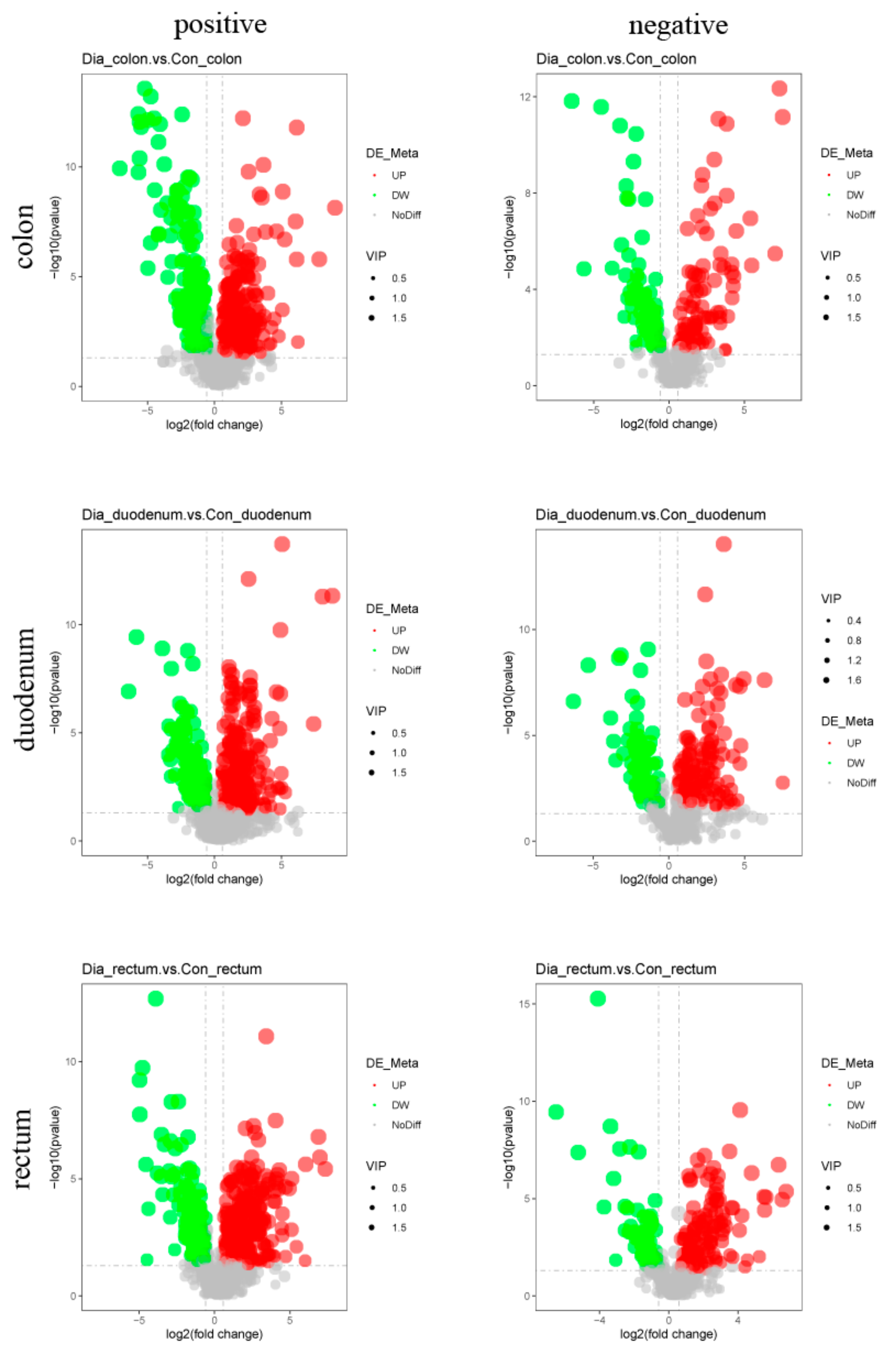
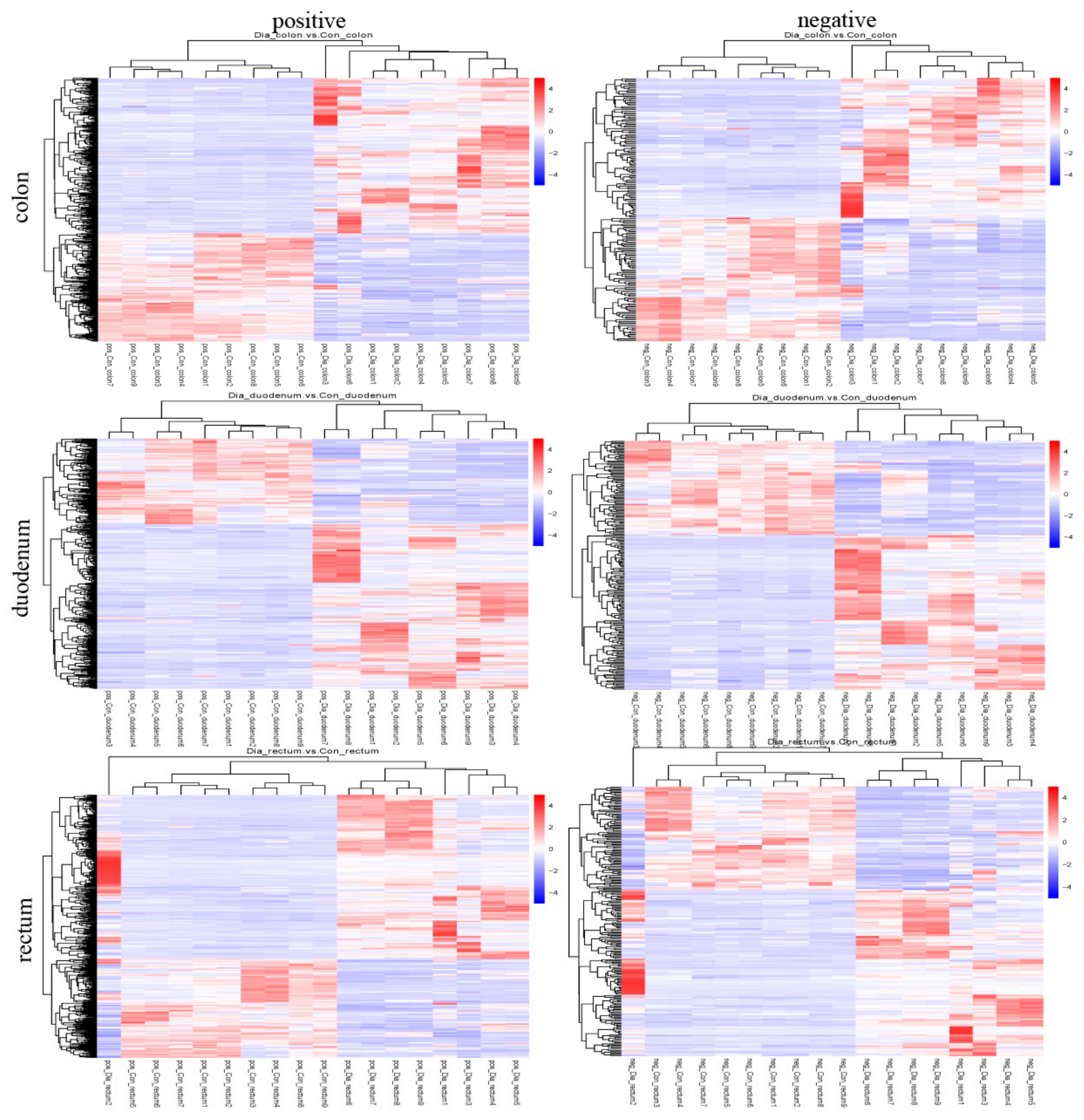
3.4. Differential Metabolite Analysis
3.5. KEGG Pathway Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hoop, R.K.; Ehrsam, H.; Keller, B. 10 years of rabbit autopsy—A review of frequent disease and mortality causes. Schweiz. Arch. Tierheilk. 1993, 135, 212–216. [Google Scholar]
- Rateau, J.G.; Blayac, M.; Parier, J.L. Infectious diarrhea: Weakening of mucosal protection caused by a pathogenic Escherichia coli in ileal loops in the rabbit. Bull. Assoc. Anat. 1982, 66, 519–529. [Google Scholar]
- Potter, M.E.; Kaufmann, A.F.; Thomason, B.M.; Blake, P.A.; Farmer, J.J. Diarrhelil Due to Escherichia coli O157:H7 in the Infant Rabbit. J. Infect. Dis. 1985, 152, 1341–1343. [Google Scholar] [CrossRef] [PubMed]
- Petric, M.; Middleton, P.J.; Grant, C.; Tam, J.S.; Hewitt, C.M. Lapine rotavirus: Preliminary studies on epizoology and transmission. Can. J. Comp. Med. Rev. Can. Med. Comp. 1978, 42, 143–147. [Google Scholar]
- Inman, L.R.; Takeuchi, A. Spontaneous Cryptosporidiosis in an Adult Female Rabbit. Vet. Pathol. 1979, 16, 89–95. [Google Scholar] [CrossRef]
- Osborne, D.J.; Huff, W.E.; Hamilton, P.B.; Burmeister, H.R. Comparison of Ochratoxin, Aflatoxin, and T-2 Toxin for Their Effects on Selected Parameters Related to Digestion and Evidence for Specific Metabolism of Carotenoids in Chickens. Poult. Sci. 1982, 61, 1646–1652. [Google Scholar] [CrossRef]
- Kelly, C. Clostridium diffeile colitis. N. Engl. J. Med. 1994, 27, 1754–1755. [Google Scholar]
- Smith, T.N.; Pesti, G.M.; Bakalli, R.I.; Kilburn, J.; Edwards, H.M. The Use of Near-Infrared Reflectance Spectroscopy to Predict the Moisture, Nitrogen, Calcium, Total Phosphorus, Gross Energy, and Phytate Phosphorus Contents of Broiler Excreta. Poult. Sci. 2001, 80, 314–319. [Google Scholar] [CrossRef]
- Black, R.E.; Merson, M.H.; Rowe, B.; Taylor, P.R.; Alim, A.R.M.A.; Gross, R.J.; Sack, D.A. En terotoxigenic Escherichia coli diarrhoea: Acquired immunity and transmission in an endemic area. Bull. World Health Organ. 1981, 59, 263–268. [Google Scholar]
- Lord, B. Gastrointestinal disease in rabbits 2. Intestinal diseases. Practice 2012, 34, 156–162. [Google Scholar] [CrossRef]
- Thornton, P.K. Livestock production: Recent trends, future prospects. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2853–2867. [Google Scholar] [CrossRef]
- Mingmongkolchai, S.; Panbangred, W. Bacillus probiotics: An alternative to antibiotics for livestock production. J. Appl. Microbiol. 2018, 124, 1334–1346. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Functional groups of additives as described in Annex 1 of Regulation (EC) No 1831/2003 Self-task of the Panel on Additives and Products or Substances used in Animal Feed. EFSA J. 2008, 6, 1–19. [Google Scholar]
- Zhou, J.; Wang, Y.; Zeng, X.; Zhang, T.; Li, P.; Yao, B.; Wang, L.; Qiao, S.; Zeng, X. Effect of antibiotic-free, low-protein diets with specific amino acid compositions on growth and intestinal flora in weaned pigs. Food Funct. 2020, 11, 493–507. [Google Scholar] [CrossRef]
- Borrmann, E.; Berndt, A.; Hänel, I.; Köhler, H. Campylobacter-induced interleukin-8 responses in human intestinal epithelial cells and primary intestinal chick cells. Vet. Microbiol. 2007, 124, 115–124. [Google Scholar] [CrossRef]
- Brisbin, J.T.; Gong, J.; Sharif, S. Interactions between commensal bacteria and the gut-associated immune system of the chicken. Anim. Health Res. Rev. 2008, 9, 101–110. [Google Scholar] [CrossRef]
- McOrist, S.; Corona-Barrera, E.; Niewold, T. Chapter 3: Intestinal diseases of pigs. Intest. Health 2015, 22, 51–70. [Google Scholar] [CrossRef]
- Olkowski, A.; Wojnarowicz, C.; Chirino-Trejo, M.; Drew, M. Responses of broiler chickens orally challenged with Clostridium perfringens isolated from field cases of necrotic enteritis. Res. Vet. Sci. 2006, 81, 99–108. [Google Scholar] [CrossRef]
- Lu, H.; Adedokun, S.A.; Adeola, L.; Ajuwon, K. Anti-Inflammatory Effects of Non-Antibiotic Alternatives in Coccidia Challenged Broiler Chickens. Int. J. Poult. Sci. 2014, 51, 14–21. [Google Scholar] [CrossRef]
- Jansson, J.K.; Willing, B.; Lucio, M.; Fekete, A.; Dicksved, J.; Halfvarson, J.; Tysk, C.; Schmitt-Kopplin, P. Metabolomics Reveals Metabolic Biomarkers of Crohn’s Disease. PLoS ONE 2009, 4, e6386. [Google Scholar] [CrossRef]
- Want, E.J.; Masson, P.; Michopoulos, F.; Wilson, I.D.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Loftus, N.; Holmes, E.; Nicholson, J.K. Global metabolic profiling of animal and human tissues via UPLC-MS. Nat. Protoc. 2012, 8, 17–32. [Google Scholar] [CrossRef]
- Rao, G.; Sui, J.; Zhang, J. Metabolomics reveals significant variations in metabolites and correlations regarding the maturation of walnuts (Juglans regia L.). Biol. Open 2016, 5, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef] [PubMed]
- Want, E.J.; Wilson, I.D.; Gika, H.G.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.P.; Holmes, E.; Nicholson, J.K. Global metabolic profiling procedures for urine using UPLC–MS. Nat. Protoc. 2010, 5, 1005–1018. [Google Scholar] [CrossRef] [PubMed]
- Thilsted, J.P.; Newton, W.M.; Crandell, R.A.; Bevill, R.F. Fatal diarrhea in rabbits resulting from the feeding of antibi-otic-contaminated feed. J. Am. Vet. Med. Assoc. 1981, 179, 360–362. [Google Scholar] [PubMed]
- Gutiérrez, I.; Espinosa, A.; García, J.; Carabaño, R.; De Blas, J.C. Effect of levels of starch, fiber, and lactose on digestion and growth performance of early-weaned rabbits. J. Anim. Sci. 2002, 80, 1029–1037. [Google Scholar] [CrossRef]
- Marounek, M.; Vovk, S.J.; Skřivanová, V. Distribution of activity of hydrolytic enzymes in the digestive tract of rabbits. Br. J. Nutr. 1995, 73, 463–469. [Google Scholar] [CrossRef]
- Hu, C.; Xiao, K.; Luan, Z.S.; Song, J. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs. J. Anim. Sci. 2013, 91, 1094–1101. [Google Scholar] [CrossRef]
- Smith, F.; Clark, J.E.; Overman, B.L.; Tozel, C.C.; Huang, J.H.; Rivier, J.E.F.; Blisklager, A.T.; Moeser, A.J. Early weaning stress impairs development of mucosal barrier function in the porcine intestine. Am. J. Physiol. Liver Physiol. 2010, 298, G352–G363. [Google Scholar] [CrossRef]
- Zhang, H.; Yin, J.; Li, D.; Zhou, X.; Li, X. Tryptophan enhances ghrelin expression and secretion associated with increased food intake and weight gain in weanling pigs. Domest. Anim. Endocrinol. 2007, 33, 47–61. [Google Scholar] [CrossRef]
- Sun, X.Z.; Sun, D.Y.; Zhao, Y.C.; Zhou, Q.Q.; Wang, G.; Qin, S.K. Alteration of fecal tryptophan metabolism correlates with shifted microbiota and may be involved in pathogenesis of colorectal cancer. World J. Gastroenterol. 2020, 26, 7173–7190. [Google Scholar] [CrossRef]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease—Sci-enceDirect. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef]
- Liu, Y.; Hou, Y.; Wang, G.; Zheng, X.; Hao, H. Gut Microbial Metabolites of Aromatic Amino Acids as Signals in Host–Microbe Interplay. Trends Endocrinol. Metab. 2020, 31, 818–834. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Holmes, E.; Kinross, J.; Burcelin, R.; Gibson, G.; Jia, W.; Pettersson, S. Host-Gut Microbiota Metabolic Interactions. Science 2012, 336, 1262–1267. [Google Scholar] [CrossRef]
- Keszthelyi, D.; Troost, F.J.; Masclee, A.A.M. Understanding the role of tryptophan and serotonin metabolism in gastroin-testinal function. Neurogastroenterol. Motil. 2010, 21, 1239–1249. [Google Scholar] [CrossRef]
- Aragozzini, F.; Ferrari, A.; Pacini, N.; Gualandris, R. Indole-3-lactic acid as a tryptophan metabolite produced by Bifidobacterium spp. Appl. Environ. Microbiol. 1979, 38, 544–546. [Google Scholar] [CrossRef]
- Jiang, P.; Trimigno, A.; Stanstrup, J.; Khakimov, B.; Viereck, N.; Engelsen, S.B.; Sangild, P.T.; Dragsted, L.O. Antibiotic Treatment Preventing Necrotising Enterocolitis Alters Urinary and Plasma Metabolomes in Preterm Pigs. J. Proteome Res. 2017, 16, 3547–3557. [Google Scholar] [CrossRef]
- Wlodarska, M.; Luo, C.; Kolde, R.; D’Hennezel, E.; Annand, J.W.; Heim, C.E.; Krastel, P.; Schmitt, E.K.; Omar, A.S.; Creasey, E.A.; et al. Indoleacrylic Acid Produced by Commensal Peptostreptococcus Species Suppresses Inflammation. Cell Host Microbe 2017, 22, 25–37. [Google Scholar] [CrossRef]
- Scott, S.A.; Fu, J.; Chang, P.V. Microbial tryptophan metabolites regulate gut barrier function via the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. USA 2020, 117, 19376–19387. [Google Scholar] [CrossRef]
- Megna, B.W.; Carney, P.R.; Depke, M.G.; Nukaya, M.; McNally, J.; Larsen, L.; Rosengren, R.J.; Kennedy, G.D.; Larson, L. The aryl hydrocarbon receptor as an antitumor target of synthetic curcuminoids in colorectal cancer. J. Surg. Res. 2017, 213, 16–24. [Google Scholar] [CrossRef]
- Bryant, W.; Megna, B.S.; Patrick, R.; Carney, B.S.; Manabu, N.; Geiger, B.S.P.; Gregory, D.; Kennedy, M.D. Indole-3-carbinol induces tumor cell death: Function follows form. J. Surg. Res. 2016, 204, 47–54. [Google Scholar]
- Metidji, A.; Omenetti, S.; Crotta, S.; Li, Y.; Nye, E.; Ross, E.; Li, V.; Maradana, M.R.; Schiering, C.; Stockinger, B. The Environmental Sensor AHR Protects from Inflammatory Damage by Maintaining Intestinal Stem Cell Homeostasis and Barrier Integrity. Immunity 2018, 49, 353–362. [Google Scholar] [CrossRef]
- Bansal, T.; Alaniz, R.C.; Wood, T.K.; Jayaraman, A. The bacterial signal indole increases epithelial-cell tight-junction resistance and attenuates indicators of inflammation. Proc. Natl. Acad. Sci. USA 2010, 107, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, I.; Rizzo, A.; Sarra, M.; Sica, G.; Sileri, P.; Biancone, L.; Macdonald, T.T.; Pallone, F.; Monteleone, G. Aryl Hydrocarbon Receptor-Induced Signals Up-regulate IL-22 Production and Inhibit Inflammation in the Gastrointestinal Tract. Gastroenterology 2011, 141, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Popolo, A.; Pinto, A.; Daglia, M.; Nabavi, S.F.; Farooqi, A.A.; Rastrelli, L. Two likely targets for the anti-cancer effect of indole derivatives from cruciferous vegetables: PI3K/Akt/mTOR signalling pathway and the aryl hydrocarbon receptor. Semin. Cancer Biol. 2017, 46, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Gjaltema, R.A.F.; Bank, R.A. Molecular insights into prolyl and lysyl hydroxylation of fibrillar collagens in health and disease. Crit. Rev. Biochem. Mol. Biol. 2016, 52, 74–95. [Google Scholar] [CrossRef]
- Kellokumpu, S.; Sormunen, R.; Heikkinen, J.; Myllylä, R. Lysyl hydroxylase, a collagen processing enzyme, exemplifies a novel class of luminally-oriented peripheral membrane proteins in the endoplasmic reticulum. J. Biol. Chem. 1994, 269, 30524–30529. [Google Scholar] [CrossRef]
- Veiga-Da-Cunha, M.; Hadi, F.; Balligand, T.; Stroobant, V.; Van Schaftingen, E. Molecular Identification of Hydroxylysine Kinase and of Ammoniophospholyases Acting on 5-Phosphohydroxy-l-lysine and Phosphoethanolamine. J. Biol. Chem. 2012, 287, 7246–7255. [Google Scholar] [CrossRef]
- Cleveland, B.M.; Kiess, A.S.; Blemings, K.P. α-Aminoadipate δ-Semialdehyde Synthase mRNA Knockdown Reduces the Lysine Requirement of a Mouse Hepatic Cell Line. J. Nutr. 2008, 138, 2143–2147. [Google Scholar] [CrossRef]
- Villa, E.; Ali, E.S.; Sahu, U.; Ben-Sahra, I. Cancer Cells Tune the Signaling Pathways to Empower de Novo Synthesis of Nucleotides. Cancers 2019, 11, 688. [Google Scholar] [CrossRef]
- Raquel, B.; Aird, K.M. Deoxyribonucleotide Triphosphate Metabolism in Cancer and Metabolic Disease. Front. Endocrinol. 2018, 9, 177. [Google Scholar]
- Löffler, M.; Fairbanks, L.D.; Zameitat, E.; Marinaki, A.M.; Simmonds, H.A. Pyrimidine pathways in health and disease. Trends Mol. Med. 2005, 11, 430–437. [Google Scholar] [CrossRef]
- Blay, J.; White, T.D.; Hoskin, D.W. The Extracellular Fluid of Solid Carcinomas Contains Immunosuppressive Concentrations of Adenosine. Cancer Res. 1997, 57, 2602–2605. [Google Scholar]
- Burnstock, G. Purinergic Signalling and Neurological Diseases: An Update. CNS Neurol. Disord. Drug Targets 2017, 16, 257–265. [Google Scholar] [CrossRef]
- Schneider, G.; Glaser, T.; Lameu, C.; Abdelbaset-Ismail, A.; Sellers, Z.P. Extracellular nucleotides as novel, underappreciated prometastatic factors that stimulate purinergic signaling in human lung cancer cells. Mol. Cancer 2015, 14, 201. [Google Scholar] [CrossRef]
- Zhang, W.C.; Shyh-Chang, N.; Yang, H.; Rai, A.; Umashankar, S.; Ma, S.; Soh, B.S.; Sun, L.L.; Tai, B.C.; Nga, M.E.; et al. Glycine Decarboxylase Activity Drives Non-Small Cell Lung Cancer Tumor-Initiating Cells and Tumorigenesis. Cell 2012, 148, 259–272. [Google Scholar] [CrossRef]
- Feinberg, A.P.; Gehrke, C.W.; Kuo, K.C.; Ehrlich, M. Reduced genomic 5-methylcytosine content in human colonic neoplasia. Cancer Res. 1988, 48, 1159–1161. [Google Scholar]
- Barciszewska, A.-M.; Murawa, D.; Gawronska, I.; Murawa, P.; Nowak, S.; Barciszewska, M.Z. Analysis of 5-Methylcytosine in DNA of Breast and Colon Cancer Tissues. IUBMB Life 2007, 59, 765–770. [Google Scholar] [CrossRef]
- Li, J.; Huang, Q.; Zeng, F.; Li, W.; He, Z.; Chen, W.; Zhu, W.; Zhang, B. The Prognostic Value of Global DNA Hypomethylation in Cancer: A Meta-Analysis. PLoS ONE 2014, 9, e106290. [Google Scholar] [CrossRef]
- Jordi, F.; Xavier, S.; Paz, M.F.; Victor, M.; Manel, E.; Gabriel, C.; Peinado, M.A. Differential DNA hypermethylation and hypomethylation signatures in colorectal cancer. Hum. Mol. Genet. 2005, 14, 319–326. [Google Scholar]
- Mirna, S.; Nives, L.; Dubravka, Ž.K.; Leila, P.; Dunja, I.; Maja, M.; Solomun, G.F. Immunoexpression of 5-methylcytosine (5mc) in Bone Marrow Haematopoietic Cells in Patients with Myelodysplastic Syndromes. Ann. Clin. Lab. Sci. 2019, 49, 810–817. [Google Scholar]
- Hongchang, G.; Qi, S.; Jiuxia, C.; Kai, F.; Pengtao, X.; Qi, Z.; Chen, L.; Hong, Z. Antibiotic Exposure Has Sex-Dependent Effects on the Gut Microbiota and Metabolism of Short-Chain Fatty Acids and Amino Acids in Mice. Msystems 2019, 4. [Google Scholar] [CrossRef]
- Bloxam, D.L. Nutritional aspects of amino acid metabolism. 1. A rat liver perfusion method for the study of amino acid metabolism. Br. J. Nutr. 1972, 27, 233–247. [Google Scholar] [CrossRef]
- Dong, S.; Zhan, Z.Y.; Cao, H.Y.; Wu, C.; Bian, Y.Q.; Li, J.Y.; Cheng, G.H.; Liu, P.; Sun, M.Y. Urinary metabolomics analysis identifes key biomarkers of diferent stages of nonalcoholic fatty liver disease. World J. Gastroenterol. 2017, 23, 2771–2784. [Google Scholar] [CrossRef] [PubMed]
- Rechner, A.R.; Smith, M.A.; Kuhnle, G.; Gibson, G.R.; Debnam, E.S.; Srai, S.S.; Moore, K.P.; Rice-Evans, C.A. Colonic Metabolism of Dietary Polyphenols: Influence of Structure on Microbial Fermentation Products. Free. Radic. Biol. Med. 2003, 36, 212–225. [Google Scholar] [CrossRef]
- Fuchs, D.; Sperner-Unterweger, B.; Leblhuber, F.; Kurz, K.; Geisler, S.; Capuron, L. Activated Immune System and Inflammation in Healthy Ageing: Relevance for Tryptophan and Neopterin Metabolism. Curr. Pharm. Des. 2014, 20, 6048–6057. [Google Scholar]
- Fujita, T.; Amuro, Y.; Hada, T.; Higashino, K. Plasma levels of pipecolic acid, both L- and D-enantiomers, in patients with chronic liver diseases, especially hepatic encephalopathy. Clin. Chim. Acta 1999, 287, 99–109. [Google Scholar]
- Singla, P.; Dalal, P.; Kaur, M.; Arya, G.; Nimesh, S.; Singh, R.; Salunke, D.B. Bile Acid Oligomers and Their Combination with Antibiotics to Combat Bacterial Infections. J. Med. Chem. 2018, 61, 10265–10275. [Google Scholar] [CrossRef]
- Hofmann, A.F. Bile Acids: The Good, the Bad, and the Ugly. Physiology 1999, 14, 24–29. [Google Scholar] [CrossRef]
- Ehrlich, D.; Pavlidis, P.; Bjarnason, I.; Vincent, R.; Hayee, P. Systematic review: Bile acids and intestinal inflammation-luminal aggressors or regulators of mucosal defence? Aliment. Pharmacol. Ther. 2015, 42, 802–817. [Google Scholar]
- Hofmann, A.F.; Hagey, L.R. Bile Acid Chemistry, Biology, and Therapeutics during the Last 80 Years: Historical Aspects. J. Lipid Res. 2014, 55, 1553. [Google Scholar] [CrossRef]
- Vancamelbeke, M.; Vermeire, S. The intestinal barrier: A fundamental role in health and disease. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef]
- Münch, A.; Strm, M.; Sderholm, J.D. Dihydroxy bile acids increase mucosal permeability and bacterial uptake in human colon biopsies. Scand. J. Gastroenterol. 2007, 42, 1167–1174. [Google Scholar] [CrossRef]
- Tiratterra, E.; Franco, P.; Porru, E.; Katsanos, K.H.; Christodoulou, D.K.; Roda, G. Role of bile acids in inflammatory bowel disease. Ann. Gastroenterol. 2018, 31, 266–272. [Google Scholar] [CrossRef]
- Ward, J.B.J.; Lajczak, N.K.; Kelly, O.B.; O’Dwyer, A.M.; Giddam, A.K.; Gabhann, J.N.; Franco, P.; Tambuwala, M.M.; Jefferies, C.A.; Keely, S.; et al. Ursodeoxycholic acid and lithocholic acid exert anti-inflammatory actions in the colon. Am. J. Physiol. Liver Physiol. 2017, 312, G550–G558. [Google Scholar] [CrossRef]
- Lajczak-McGinley, N.K.; Porru, E.; Fallon, C.M.; Smyth, J.; Curley, C.; McCarron, P.A.; Tambuwala, M.M.; Roda, A.; Keely, S.J. The secondary bile acids, ursodeoxycholic acid and lithocholic acid, protect against intestinal inflammation by inhibition of epithelial apoptosis. Physiol. Rep. 2020, 8, e14456. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
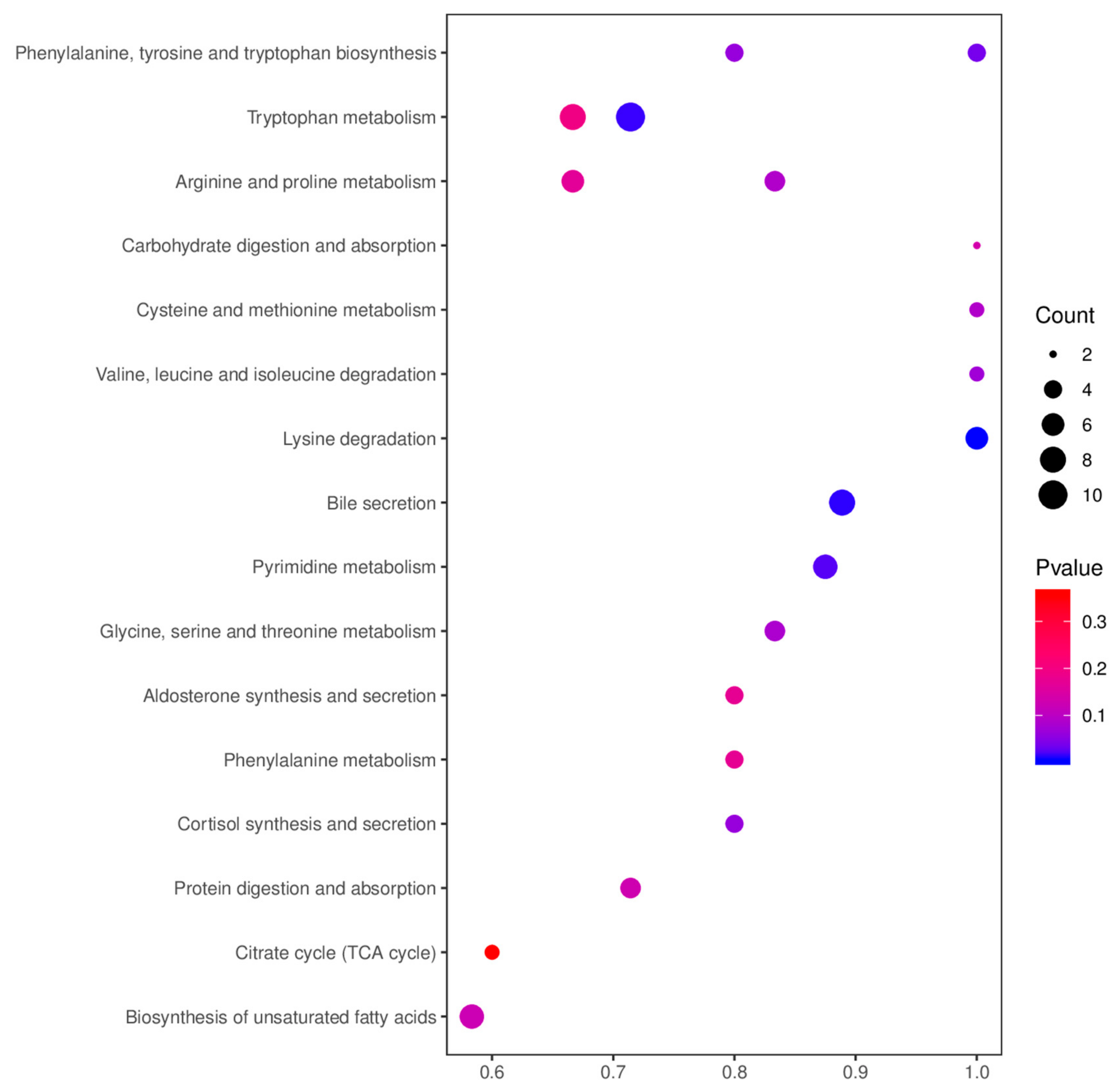{kind=link}
{kind=link}
| Intestinal Tissue Comparison | Ion Mode | N Total Identified 1 | N Signif Different 2 | N Signif Up 3 | N Signif Down 4 |
|---|---|---|---|---|---|
| Dia_colon vs. Con_colon | Positive | 1191 | 472 | 278 | 194 |
| Negative | 472 | 179 | 95 | 84 | |
| Sum | 1663 | 651 | 373 | 278 | |
| Dia_duodenum vs. Con_duodenum | Positive | 1096 | 432 | 284 | 148 |
| Negative | 493 | 204 | 127 | 77 | |
| Sum | 1589 | 636 | 411 | 225 | |
| Dia_rectum vs. Con_rectum | Positive | 1224 | 471 | 294 | 177 |
| Negative | 522 | 211 | 130 | 81 | |
| Sum | 1746 | 682 | 424 | 258 | |
| All intestinal tissue comparisons | Total | 4998 | 1969 | 1208 | 761 |
| Tissue | Map ID | Map Title | p-Value | N | Meta IDs |
|---|---|---|---|---|---|
| colon | map04976 | Bile secretion | 0.204218 | 94 | Salicylic acid, Deoxycholic acid, Lithocholic Acid, and Chenodeoxycholic Acid. |
| map00400 | Phenylalanine, tyrosine and tryptophan biosynthesis | 0.063043 | 184 | Anthranilic acid, Phenylpyruvic acid, L-Tryptophan, and Indole. | |
| map04927 | Cortisol synthesis and secretion | 0.063043 | 184 | Cortisol, Pregnenolone, Progesterone, and Cortodoxone. | |
| map04934 | Cushing’s syndrome | 0.063043 | 184 | Cortisol, Pregnenolone, Progesterone, and Cortodoxone. | |
| duodenum | map00330 | Arginine and proline metabolism | 0.089757 | 159 | Creatine, Spermine, S-Adenosylmethionine, Creatinine, and L-Glutamic acid. |
| map00360 | Phenylalanine metabolism | 0.173262 | 159 | 2-Phenylacetamide, Phenylacetylglycine, L-Phenylalanine, and D-Phenylalanine. | |
| map00380 | Tryptophan metabolism | 0.195911 | 159 | Anthranilic acid, L-Tryptophan, N-Acetylserotonin, Kynurenic acid, Xanthurenic acid, 5-Hydroxyindoleacetate, N-Formylkynurenine, and Indole. | |
| duodenum | map00053 | Ascorbate and aldarate metabolism | 0.197835 | 159 | L-Ascorbate and UDP-D-glucuronate. |
| map00270 | Cysteine and methionine metabolism | 0.089654 | 106 | L-Aspartic acid, Glutathione, and Reduced Glutathione. | |
| map04925 | Aldosterone synthesis and secretion | 0.173475 | 106 | cGMP, Aldosterone, Pregnenolone, and NAD+. | |
| rectum | map00220 | Arginine biosynthesis | 0.070142 | 178 | L-Glutamic acid, N-Acetyl-L-glutamic acid, and L-Ornithine. |
| map00280 | Valine, leucine and isoleucine degradation | 0.070142 | 178 | 3-Methyl-2-oxobutanoic acid, TPP, and Acetoacetate. | |
| map00260 | Glycine, serine and threonine metabolism | 0.083262 | 178 | O-Phospho-L-serine, Creatine, Betaine, L-Tryptophan, and L-Cystathionine. | |
| map04974 | Protein digestion and absorption | 0.128992 | 178 | Indole, L-Asparagine, Histamine, L-Tryptophan, and Tyramine. | |
| map00330 | Arginine and proline metabolism | 0.166117 | 178 | L-Glutamic acid, Spermine, Creatine, Creatinine, N-Methylhydantoin, and L-Ornithine. | |
| map01040 | Biosynthesis of unsaturated fatty acids | 0.125115 | 110 | Nervonic acid, Docosanoic acid, Adrenic acid, Stearic acid, Erucic acid, Arachidic acid, and Docosahexaenoic acid. | |
| map04973 | Carbohydrate digestion and absorption | 0.136781 | 110 | Maltotriose and D-Glucose 6-phosphate. | |
| map00020 | Citrate cycle (TCA cycle) | 0.35937 | 110 | Cis-Aconitic acid, L-Malate, and Fumaric acid. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, T.; Li, Y.; Wang, J.; Elzo, M.A.; Shao, J.; Li, Y.; Xia, S.; Fan, H.; Jia, X.; Lai, S. Untargeted Metabolomics Reveals Intestinal Pathogenesis and Self-Repair in Rabbits Fed an Antibiotic-Free Diet. Animals 2021, 11, 1560. https://doi.org/10.3390/ani11061560
Tang T, Li Y, Wang J, Elzo MA, Shao J, Li Y, Xia S, Fan H, Jia X, Lai S. Untargeted Metabolomics Reveals Intestinal Pathogenesis and Self-Repair in Rabbits Fed an Antibiotic-Free Diet. Animals. 2021; 11(6):1560. https://doi.org/10.3390/ani11061560
Chicago/Turabian StyleTang, Tao, Ya Li, Jie Wang, Mauricio A. Elzo, Jiahao Shao, Yanhong Li, Siqi Xia, Huimei Fan, Xianbo Jia, and Songjia Lai. 2021. "Untargeted Metabolomics Reveals Intestinal Pathogenesis and Self-Repair in Rabbits Fed an Antibiotic-Free Diet" Animals 11, no. 6: 1560. https://doi.org/10.3390/ani11061560
APA StyleTang, T., Li, Y., Wang, J., Elzo, M. A., Shao, J., Li, Y., Xia, S., Fan, H., Jia, X., & Lai, S. (2021). Untargeted Metabolomics Reveals Intestinal Pathogenesis and Self-Repair in Rabbits Fed an Antibiotic-Free Diet. Animals, 11(6), 1560. https://doi.org/10.3390/ani11061560