
Selected Alternative Feed Additives Used to Manipulate the Rumen Microbiome

 , ,
, ,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
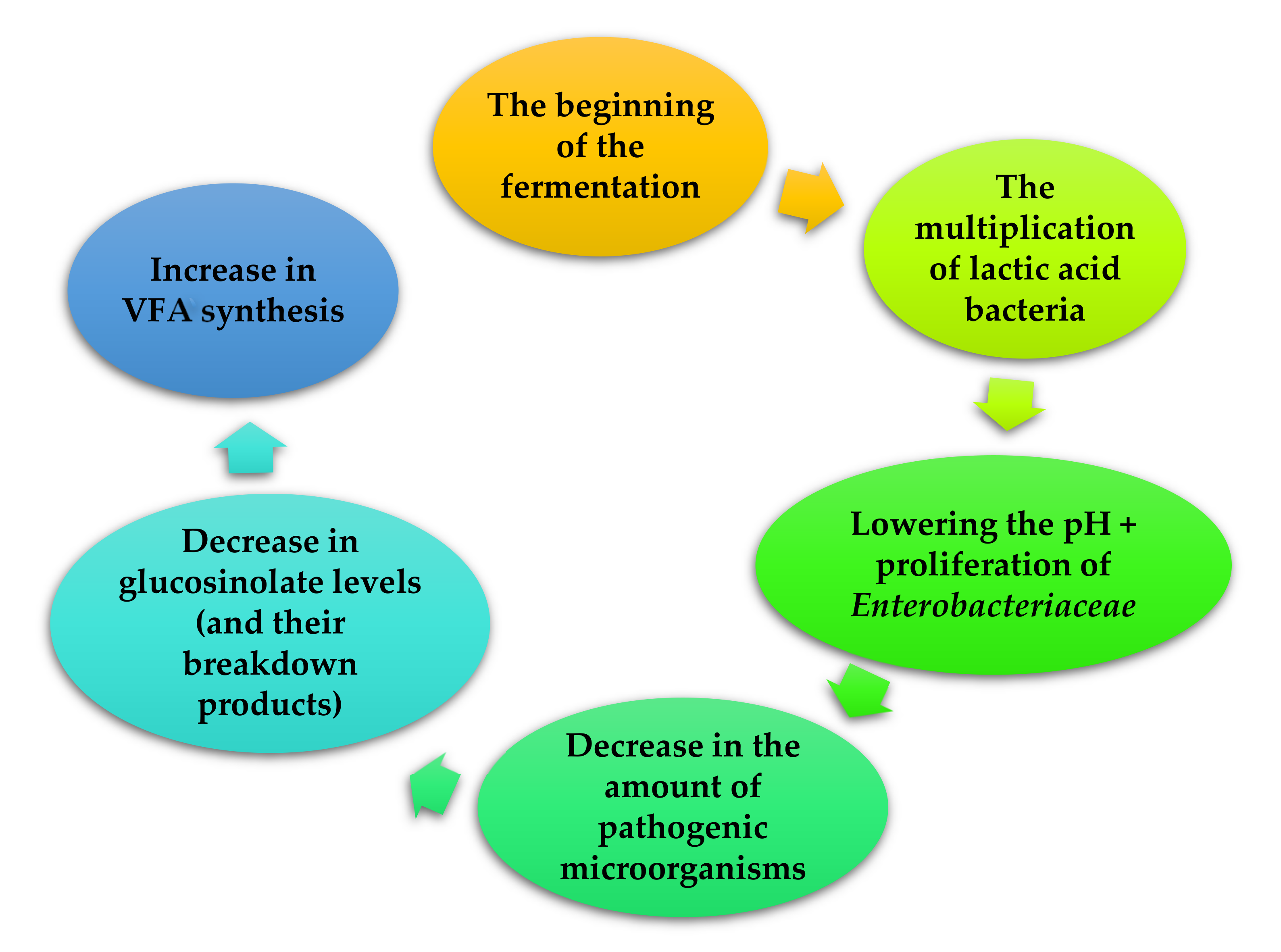
2. Fermented Feeds
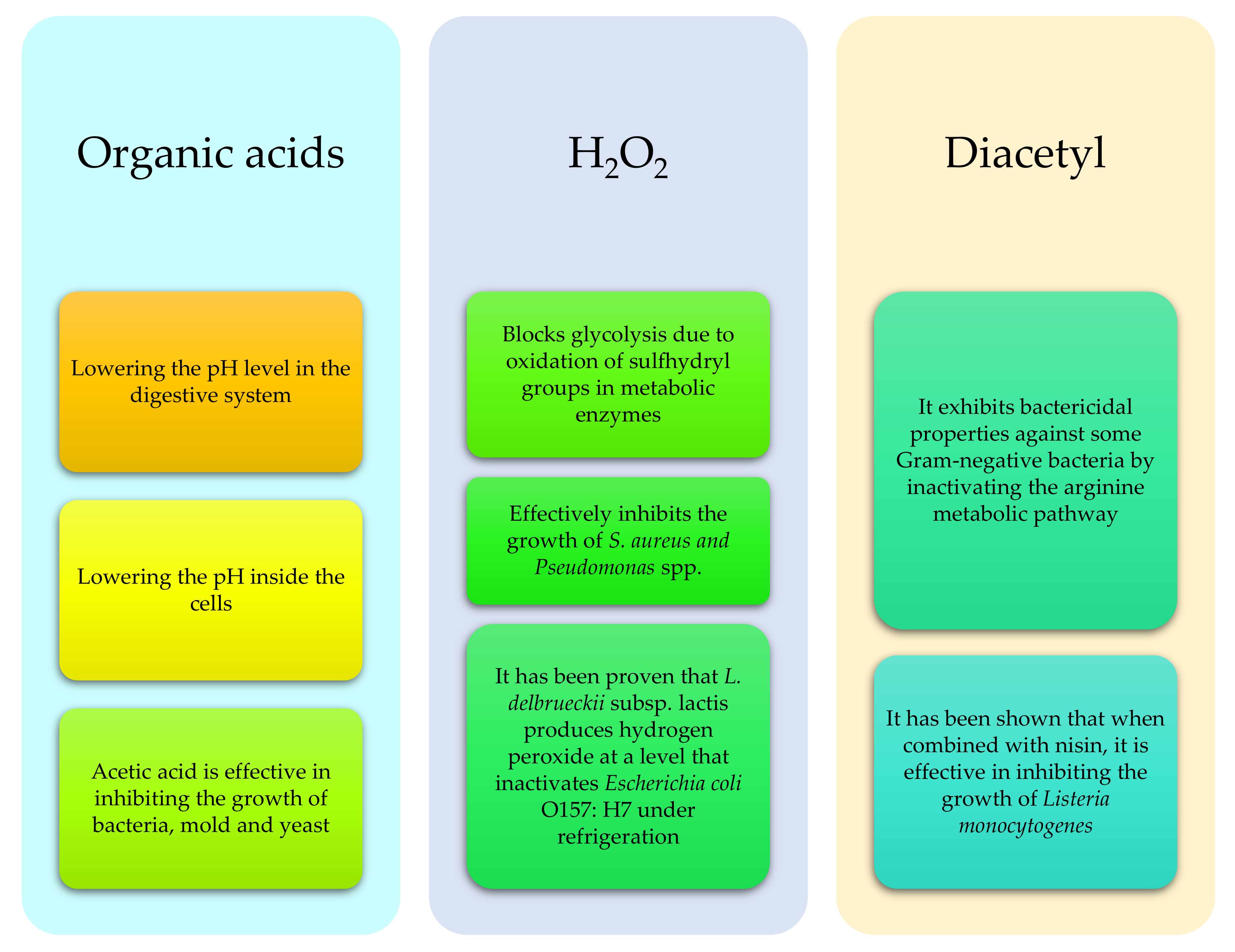
3. Probiotics and Prebiotics
- Preservation of the ability to survive in the digestive system,
- Non-pathogenic and non-toxic,
- Taxonomic affiliation established with modern genetic methods (genus, species and strain),
- Show clinically documented beneficial effects on health,
- Be safe, i.e., show no undesirable side effects,
- Demonstrate stability and the possibility of large-scale production of biomass.
4. Algae
5. Selected Additives of Plant Origin
6. Summary
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- FAO: Guidelines for the Evaluation of Probiotics in Food, Report of a Joint FAO/WHO Working Group on Drafting Guidelines for the Evaluation of Probiotics in Food. London, Ontario, Canada, April 30 and May 1. 2002. Available online: https://www.who.int/foodsafety/fs_management/en/probiotic_guidelines.pdf. (accessed on 15 April 2021).
- Abbate, J.M.; Macrì, F.; Capparucci, F.; Iaria, C.; Briguglio, G.; Cicero, L.; Salvo, A.; Arfuso, F.; Ieni, A.; Piccione, G.; et al. Administration of Protein Hydrolysates from Anchovy (Engraulis Encrasicolus) Waste for Twelve Weeks Decreases Metabolic Dysfunction-Associated Fatty Liver Disease Severity in ApoE–/–Mice. Animals 2020, 10, 2303. [Google Scholar] [CrossRef]
- Avondo, M.; Pagano, R.I.; Guastella, A.M.; Criscione, A.; Di Gloria, M.; Valenti, B.; Piccione, G.; Pennisi, D. Diet selection and milk production and composition in Girgentana goats with different as1-casein genotype. J. Dairy Res. 2009, 76, 202–209. [Google Scholar] [CrossRef]
- Armato, L.; Gianesella, M.; Morgante, M.; Fiore, E.; Rizzo, M.; Giudice, E.; Piccione, G. Rumen volatile fatty acids × dietary supplementation with live yeast and yeast cell wall in feedlot beef cattle. Acta Agric. Scand. A Anim. Sci. 2016, 66, 119–124. [Google Scholar] [CrossRef]
- Monteverde, V.; Congiu, F.; Vazzana, I.; Dara, S.; Di Pietro, S.; Piccione, G. Serum lipid profile modification related to polyunsaturated fatty acid supplementation in thoroughbred horses. J. Appl. Ani. Res. 2017, 45, 615–618. [Google Scholar] [CrossRef]
- Piccione, G.; Marafioti, S.; Giannetto, C.; Panzera, M.; Fazio, F. Effect of dietary supplementation with omega 3 on clotting time, fibrinogen concentration and platelet aggregation in the athletic horse. Livest. Sci. 2014, 161, 109–113. [Google Scholar] [CrossRef]
- Piccione, G.; Arfuso, F.; Fazio, F.; Bazzano, M.; Giannetto, C. Serum Lipid Modification Related to Exercise and Polyunsaturated Fatty Acid Supplementation in Jumpers and Thoroughbred Horses. J. Equine Vet. Sci. 2014, 34, 1181–1187. [Google Scholar] [CrossRef]
- Bazzano, M.; Rizzo, M.; Arfuso, F.; Giannetto, C.; Fazio, F.; Piccione, G. Increase in erythrocyte osmotic resistance following polyunsaturated fatty acids (PUFA) supplementation in show jumper horses. Livest. Sci. 2015, 181, 236–241. [Google Scholar] [CrossRef]
- Piccione, G.; Giannetto, C.; Bruschetta, D.; Congiu, F.; Arfuso, F.; Giudice, E. Influence of exercise and dietary omega-3 oil supplementation on interleukin 1-Ra serum concentrations in Standardbred horses. Ani. Product. Sci. 2019, 59, 232–235. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; McGinn, S.M. Methane emissions from feedlot cattle fed barley or corn diets. J. Ani. Sci. 2005, 83, 653–661. [Google Scholar] [CrossRef]
- Kassow, A.; Rahmann, G.; Blank, B.; Paulsen, H.M.; Aulrich, K. Studies on greenhouse gas emissions in organic and conventional dairy farms. In Ressortforschung für den Ökologischen Landbau 2009; Johann Heinrich von Thünen-Institut-Bundesforschungsinstitut für Ländliche Räume, Wald und Fischerei (vTI): Braunschweig, Germany, 2010; pp. 65–76. [Google Scholar]
- Moss, A.R.; Jouany, J.P.; Newbold, J. Methane production by ruminants: Its contribution to global warming. Ann. Zootech. 2000, 49, 231–253. [Google Scholar] [CrossRef]
- Malmuthuge, N.; Guan, L.L. Gut microbiome and omics: A new definition to ruminant production and health. Anim. Front. 2016, 6, 8–12. [Google Scholar] [CrossRef]
- Qian, X.; Chen, T.; Xu, Y.-P.; Chen, L.; Sun, F.-X.; Lu, M.-P.; Liu, Y.-X. A guide to human microbiome research: Study design, sample collection, and bioinformatics analysis. Chin. Med. J. 2020, 133, 1844–1855. [Google Scholar] [CrossRef] [PubMed]
- Honan, M.; Feng, A.; Tricarico, J.M.; Kebreab, E. Feed additives as a strategic approach to reduce enteric methane production in cattle: Modes of action, effectiveness and safety. Anim. Prod. Sci. 2021. [Google Scholar] [CrossRef]
- Faiz-ul, H.; Muhammad, A.A.; Hossam, M.E.; Muhammad, S.R.; Muhammad, S.K.; Shehryaar, S.; Yang, C. Phytogenic Additives Can Modulate Rumen Microbiome to Mediate Fermentation Kinetics and Methanogenesis Through Exploiting Diet–Microbe Interaction. Front. Vet. Sci. 2020, 7. [Google Scholar] [CrossRef]
- Lascano, C.E.; Cárdenas, E. Alternatives for methane emission mitigation in livestock systems. R. Bras. Zootec. 2010, 39, 175–182. [Google Scholar] [CrossRef]
- Newbold, C.J.; Wallace, R.J.; McIntosh. Mode of action of the yeast Saccharomyces cerevisiae as a feed additive for ruminants. Br. J. Nutr. 1996, 76, 249–261. [Google Scholar] [CrossRef]
- Parker, D.S.; Armstrong, D.G. Antibiotic feed additives and livestock production. Proc. Nutr. Soc. 1987, 46, 415–421. [Google Scholar] [CrossRef]
- Maeda, K.; Nguyen, V.T.; Suzuki, T.; Yamada, K.; Kudo, K.; Hikita, C.; Le, V.P.; Nguyen, M.C.; Yoshida, N. Network analysis and functional estimation of the microbiome reveal the effects of cashew nut shell liquid feeding on methanogen behaviour in the rumen. Microb. Biotechnol. 2020, 14, 277–290. [Google Scholar] [CrossRef]
- Roque, B.M.; Brooke, C.G.; Ladau, J.; Polley, T.; Marsh, L.J.; Najafi, N.; Pandey, P.; Singh, L.; Kinley, R.; Salwen, J.K.; et al. Effect of the macroalgae Asparagopsis taxiformis on methane production and rumen microbiome assemblage. Ani. Microbiome 2019, 1, 1–14. [Google Scholar]
- Zhou, M.; Hünerberg, M.; Chen, Y.; Reuter, T.; McAllister, T.A.; Evans, F.; Critchley, A.T.; Guan, L.L. Air-Dried Brown Seaweed, Ascophyllum nodosum, Alters the Rumen Microbiome in a Manner That Changes Rumen Fermentation Profiles and Lowers the Prevalence of Foodborne Pathogens. Appl. Environ. Sci. 2018, 3, 1–18. [Google Scholar] [CrossRef]
- Uritskiy, G.V.; DiRuggiero, J.; Taylor, J. MetaWRAP—a flexible pipeline for genome-resolved metagenomic data analysis. Microbiome 2018, 6, 158. [Google Scholar] [CrossRef] [PubMed]
- Herlinae, H.; Yemima, Y.; Rumiasih, R. Pengaruh aditif EM4 dan gula merah terhadap karakteristik silase rumput gajah (Pennisetum purpureum) [Effect of additives EM4 and palm sugar on the characteristics of elephant grass (Pennisetum purpureum) silage]. J. Ilmu Hewani Trop. 2015, 4, 27–30. (In Indonesian) [Google Scholar]
- Nega, T.; Woldes, Y. Review on nutritional limitations and opportunities of using rapeseed meal and other rape seed by-products in animal feeding. J. Nutr. Health Food Eng. 2018, 8, 43–48. [Google Scholar] [CrossRef]
- Wang, C.; Shi, C.; Zhang, Y.; Song, D.; Lu, Z.; Wang, Y. Microbiota in fermented feed and swine gut. Appl. Microbiol. BioTechnol. 2018, 102, 2941–2948. [Google Scholar] [CrossRef]
- Canibe, N.; Jensen, B.B. Fermented liquid feed–microbial and nutritional aspects and impact on enteric diseases in pigs. Anim. Feed Sci. Technol. 2012, 173, 17–40. [Google Scholar] [CrossRef]
- Dai, Z.; Cui, L.; Li, J.; Wang, B.; Guo, L.; Wu, Z.; Zhu, W.; Wu, G. Fermentation techniques in feed production. Anim. Agric. 2020, 407–429. [Google Scholar]
- Basu, S.; Gaur, R.; Gomes, J.; Sreekrishnan, T.R.; Bisaria, V.S. Effect of seed culture on solid state bioconversion of wheat straw by Phanerochaete chrysosporium for animal feed production. J. Biosci. Bioeng. 2002, 1, 25–30. [Google Scholar] [CrossRef]
- Shrivastava, B.; Thakur, S.; Khasa, Y.P.; Gupte, A.; Puniya, A.K.; Kuhad, R.C. White-rot fungal conversion of wheat straw to energy rich cattle feed. Biodegradation 2011, 22, 823–831. [Google Scholar] [CrossRef]
- Chi, C.H.; Cho, S.J. Improvement of bioactivity of soybean meal by solid-state fermentation with Bacillus amyloliquefaciens versus Lactobacillus spp. and Saccharomyces cerevisiae. LWT–Food Sci. Technol. 2016, 68, 619–625. [Google Scholar] [CrossRef]
- Shi, C.; He, J.; Yu, J.; Yu, B.; Mao, X.; Zheng, P.; Huang, Z.; Chen, D. Amino acid, phosphorus, and energy digestibility of Aspergillus niger fermented rapeseed meal fed to growing pigs. J. Anim. Sci. 2015, 93, 2916–2925. [Google Scholar] [CrossRef]
- Canibe, N.; Højberg, O.; Badsberg, J.H.; Jensen, B.B. Effect of feeding fermented liquid feed and fermented grain to piglets on gastrointestinal ecology and growth performance. J. Anim. Sci. 2007, 85, 2959–2971. [Google Scholar] [CrossRef] [PubMed]
- Brooks, P.H. Fermented liquid feed for pigs. CAB Reviews: Perspectives in Agriculture. Vet. Sci. Nutr. Nat. Res. 2008, 3, 1–18. [Google Scholar]
- Kiesz, M. Efektywność stosowania fermentowanych pasz białkowych na wyniki produkcyjne oraz status zdrowotny świń. Ph.D. Thesis, Uniwersytet Przyrodniczy w Lublinie, Lublin, Poland, 2018. [Google Scholar]
- Ran, T.; Gomaa, W.M.S.; Shen, Y.Z.; Saleem, A.M.; Yang, W.Z.; McAllister, T.A. Use of naturally sourced feed additives (lactobacillus fermentation products and enzymes) in growing and finishing steers: Effects on performance, carcass characteristics and blood metabolites. Ani. Feed Sci. Technol. 2019, 254, 114190. [Google Scholar] [CrossRef]
- Azlan, P.M.; Jahromi, M.F.; Ariff, M.O.; Ebrahimi, M.; Candyrine, S.C.L.; Liang, J.B. Aspergillus terreus treated rice straw suppresses methane production and enhances feed digestibility in goats. Trop. Ani. Health. Prod. 2018, 50, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Lesmeister, K.E.; Heinrichs, A.J.; Gabler, M.T. Effects of supplemental yeast (Saccharomyces cerevisiae) culture on rumen development, growth characteristics, and blood parameters in neonatal dairy calves. J. Dairy Sci. 2004, 87, 1832–1839. [Google Scholar] [CrossRef]
- Shrivastava, B.; Nandal, P.; Sharma, A.; Jain, K.K.; Khasa, Y.P.; Das, T.K.; Mani, V.; Kewalramani, N.J.; Kundu, S.S.; Kuhad, R.C. Solid state bioconversion of wheat straw into digestible and nutritive ruminant feed by Ganoderma sp. rckk02. Bioresour. Technol. 2012, 107, 347–351. [Google Scholar] [CrossRef]
- Stefańska, B.; Sroka, J.; Katzer, F.; Goliński, P.; Nowak, W. The effect of probiotics, phytobiotics and their combination as feed additives in the diet of dairy calves on performance, rumen fermentation and blood metabolites during the preweaning period. Ani. Feed Sci. Technol. 2021, 272, 114738. [Google Scholar] [CrossRef]
- Retta, K.S. Role of probiotics in rumen fermentation and animal performance: A review. Int. J. Livest. Produc. 2016, 7, 24–32. [Google Scholar]
- Cholewińska, P.; Czyż, K.; Nowakowski, P.; Wyrostek, A. The microbiome of the digestive system of ruminants–a review. Ani. Heal. Res. Rev. 2020, 21, 3–14. [Google Scholar] [CrossRef]
- Livermore, D.M.; Williams, J.D. β-Lactams: Mode of Action and Mechanism of Bacterial Resistance. In Antibiotics in Laboratory Medicine; Lorian, V., Ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1996; pp. 502–578. [Google Scholar]
- The International Scientific Association for Probiotics and Prebiotics (ISAPP). Available online: https://isappscience.org/ (accessed on 2 May 2021).
- Gaggìa, F.; Mattarelli, P.; Biavati, B. Probiotics and prebiotics in animal feeding for safe food production. Inter. J. Food Microb. 2010, 141, 15–28. [Google Scholar] [CrossRef]
- Rolfe, R.D. The role of probiotic cultures in the control of gastrointestinal health. J. Nut. 2000, 130, 396S–402S. [Google Scholar] [CrossRef]
- Chen, J.; Harstad, O.M.; McAllister, T.; Dörsch, P.; Holo, H. Propionic acid bacteria enhance ruminal feed degradation and reduce methane production in vitro. Acta Agric. Scand. Sect. A Ani. Sci. 2020, 69, 169–175. [Google Scholar] [CrossRef]
- Schrezenmeir, J.; De Vrese, M. Probiotics, prebiotics and synbiotics-approaching a definition. Am. J. Clin. Nutr. 2001, 73, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Saulnier, D.M.; Spinler, J.K.; Gibson, G.R.; Versalovic, J. Mechanisms of probiosis and prebiosis: Considerations for enhanced functional foods. Curr. Opin. Biotechnol. 2009, 20, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Chaucheyras-Durand, F.; Durand, H. Probiotics in animal nutrition and health. Benef. Microbes 2010, 1, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Salas-Jara, M.J.; Ilabaca, A.; Vega, M.; García, A. Biofilm Forming Lactobacillus: New Challenges for the Development of Probiotics. Microorganisms 2016, 4, 35. [Google Scholar] [CrossRef] [PubMed]
- Knecht, D.; Cholewińska, P.; Jankowska-Mąkosa, A.; Czyż, K. Development of swine’s digestive tract microbiota and its relation to production indices—a review. Animals 2020, 10, 527. [Google Scholar] [CrossRef]
- Gupta, V.; Garg, R. Probiotics. Indian J. Med. Microbiol. 2009, 27, 202–209. [Google Scholar] [CrossRef]
- Gibson, G.R.; Probert, H.M.; Van Loo, J.; Rastall, R.A.; Robertfroid, M. Dietary modulation of the human colonic microbiota: Updating the concept of prebiotics. Nutr. Res. Rev. 2004, 17, 259–275. [Google Scholar] [CrossRef]
- Uyeno, Y.; Shigemori, S.; Shimosato, T. Effect of probiotics/prebiotics on cattle health and productivity. Microbes Environ. 2015, 30, 126–132. [Google Scholar] [CrossRef]
- Kido, K.; Tejima, S.; Haramiishi, M.; Uyeno, Y.; Ide, Y.; Kurosu, K.; Kushibiki, S. Provision of beta-glucan prebiotics (cellooligosaccharides and kraft pulp) to calves from pre-to post-weaning period on pasture. Anim. Sci. J. 2019, 90, 1537–1543. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, K.; Piotrowska-Cyplik, A. Metabolity bakterii kwasu mlekowego i ich zastosowanie w przemyśle. Postępy Mikrobiol. 2017, 56, 4. (In Polish) [Google Scholar]
- Cangiano, L.R.; Yohe, T.T.; Steele, M.A.; Renaud, D.L. Invited Review: Strategic use of microbial-based probiotics and prebiotics in dairy calf rearing. Appl. Anim. Sci. 2020, 36, 630–651. [Google Scholar] [CrossRef]
- Madeira, M.S.; Cardoso, C.; Lopes, P.A.; Coelho, D.; Afonso, C.; Bandarra, N.M.; Prates, J.A.M. Microalgae as feed ingredients for livestock production and meat quality: A review. Livest. Sci. 2017, 205, 111–121. [Google Scholar] [CrossRef]
- Choi, Y.Y.; Lee, S.J.; Kim, H.S.; Eom, J.S.; Kim, D.H.; Lee, S.S. The potential nutritive value of Sargassum fulvellum as a feed ingredient for ruminants. Algal Res. 2020, 45, 101761. [Google Scholar] [CrossRef]
- McCauley, J.I.; Labeeuw, L.; Jaramillo-Madrid, A.C.; Nguyen, L.N.; Nghiem, L.D.; Chaves, A.V.; Ralph, P.J. Management of enteric methanogenesis in ruminants by algal-derived feed additives. Curr. Pollut. Rep. 2020, 6, 188–205. [Google Scholar] [CrossRef]
- Choi, Y.Y.; Shin, N.H.; Lee, S.J.; Lee, Y.J.; Kim, H.S.; Eom, J.S.; Lee, S.S.; Kim, E.T.; Lee, S.S. In vitro five brown algae extracts for efficiency of ruminal fermentation and methane yield. J. Appl. Phycol. 2021, 33, 1253–1262. [Google Scholar] [CrossRef]
- Bulgariu, L. Efficient use of algae biomass loaded with essential metal ions in the manufacture of feed additives. J. Appl. Phycol. 2020, 32, 1779–1788. [Google Scholar] [CrossRef]
- Burchacka, E.; Łukaszewicz, M.; Kułażyński, M. Determination of mechanisms of action of active carbons as a feed additive. Bioorg. Chem. 2019, 93, 102804. [Google Scholar] [CrossRef]
- Villagómez-Estrada, S.; Pérez, J.F.; Darwich, L.; Vidal, A.; van Kuijk, S.; Melo-Durán, D.; Solà-Oriol, D. Effects of copper and zinc sources and inclusion levels of copper on weanling pig performance and intestinal microbiota. J. Ani. Sci. 2020, 98, 117. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K.; Saeid, A. Plant growth biostimulants, dietary feed supplements and cosmetics formulated with supercritical CO2 algal extracts. Molecules 2017, 22, 66. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Latif, H.H.; Shams El-Din, N.G.; Ibrahim, H.A.H. Antimicrobial activity of the newly recorded red alga Grateloupia doryphora collected from the Eastern Harbor, Alexandria, Egypt. J. Appl. Microbiol. 2018, 125, 1321–1332. [Google Scholar] [CrossRef] [PubMed]
- Martelli, F.; Cirlini, M.; Lazzi, C.; Neviani, E.; Bernini, V. Edible seaweeds and spirulina extracts for food application: In vitro and in situ evaluation of antimicrobial activity towards foodborne pathogenic bacteria. Foods 2020, 9, 1442. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Liu, X.; Zhao, G.; Hu, T.; Wang, Y. Potential and challenges of tannins as an alternative to in-feed antibiotics for farm animal production. Ani. Nutr. 2018, 4, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Dell’Anno, M.; Sotira, S.; Rebucci, R.; Reggi, S.; Castiglioni, B.; Rossi, R. In vitro evaluation of antimicrobial and antioxidant activities of algal extracts. Ital. J. Ani. Sci. 2020, 19, 103–113. [Google Scholar] [CrossRef]
- Koch, B.J.; Hungate, B.A.; Price, L.B. Food-animal production and the spread of antibiotic resistance: The role of ecology. Front. Ecol. Environ. 2017, 15, 309–318. [Google Scholar] [CrossRef]
- Dweba, C.C.; Zishiri, O.T.; El Zowalaty, M.E. Methicillin-resistant Staphylococcus aureus: Livestock-associated, antimicrobial, and heavy metal resistance. Infect. Drug. Resist. 2018, 11, 2497. [Google Scholar] [CrossRef]
- Van den Honert, M.S.; Gouws, P.A.; Hoffman, L.C. Importance and implications of antibiotic resistance development in livestock and wildlife farming in South Africa: A Review. South Afr. J. Anim. Sci. 2018, 48, 401–412. [Google Scholar] [CrossRef]
- Brooke, C.G.; Roque, B.M.; Najafi, N.; Gonzalez, M.; Pfefferlen, A.; DeAnda, V.; Ginsburg, D.W.; Harden, M.C.; Nuzhdin, S.V.; Salwen, J.K. Evaluation of the potential of two common Pacific coast macroalgae for mitigating methane emissions from ruminants. Front. Mar. Sci. 2020, 7, 561. [Google Scholar] [CrossRef]
- Choi, Y.Y.; Lee, S.J.; Lee, Y.J.; Kim, H.S.; Eom, J.S.; Jo, S.U.; Lee, S.S. In vitro and in situ evaluation of Undaria pinnatifida as a feed ingredient for ruminants. J. Appl. Phycol. 2019, 1–11. [Google Scholar] [CrossRef]
- Black, J.L.; Davison, T.M.; Box, I. Methane Emissions from Ruminants in Australia: Mitigation Potential and Applicability of Mitigation Strategies. Animals 2021, 11, 951. [Google Scholar] [CrossRef]
- Abecia, L.; Martín-García, A.I.; Martínez, G.; Newbold, C.J.; Yáñez-Ruiz, D.R. Nutritional intervention in early life to manipulate rumen microbial colonization and methane output by kid goats postweaning. J. Ani. Sci. 2013, 91, 4832–4840. [Google Scholar]
- Lee, S.J.; Shin, N.H.; Jeong, J.S.; Kim, E.T.; Lee, S.K.; Lee, I.D.; Lee, S.S. Effects of Gelidium amansii extracts on in vitro ruminal fermentation characteristics, methanogenesis, and microbial populations. Asian-Australas. J. Anim. Sci. 2018, 31, 71. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.Y.; Lee, S.J.; Lee, Y.J.; Kim, H.S.; Eom, J.S.; Kim, S.C.; Kim, E.T.; Lee, S.S. New challenges for efficient usage of Sargassum fusiforme for ruminant production. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Altomonte, I.; Salari, F.; Licitra, R.; Martini, M. Use of microalgae in ruminant nutrition and implications on milk quality–A review. Livest. Sci. 2018, 214, 25–35. [Google Scholar] [CrossRef]
- Ku-Vera, J.C.; Jiménez-Ocampo, R.; Valencia-Salazar, S.S.; Montoya-Flores, M.D.; Molina-Botero, I.C.; Arango, J.; Gómez-Bravo, C.A.; Aguilar-Pérez, C.F.; Solorio-Sánchez, F.J. Role of Secondary Plant Metabolites on Enteric Methane Mitigation in Ruminants. Front. Vet. Sci. 2020, 7, 584. [Google Scholar] [CrossRef] [PubMed]
- Kholif, A.E.; Matloup, O.H.; Morsy, T.A.; Abdo, M.M.; Abu Elella, A.A.; Anele, U.Y.; Swanson, K.C. Rosemary and lemongrass herbs as phytogenic feed additives to improve efficient feed utilization, manipulate rumen fermentation and elevate milk production of Damascus goats. Livest. Sci. 2017, 204, 39–46. [Google Scholar] [CrossRef]
- Kolling, G.J.; Stivanin, S.C.B.; Gabbi, A.M.; Machado, F.S.; Ferreira, A.L.; Campos, M.M.; Tomich, T.R.; Cunha, C.S.; Dill, S.W.; Pereira, L.G.R.; et al. Performance and methane emissions in dairy cows fed oregano and green tea extracts as feed additives. J. Dairy Sci. 2018, 101, 4221–4234. [Google Scholar] [CrossRef]
- Velasco, V.; Williams, P. Improving meat quality through natural antioxidants. Chil. J. Agric. Res. 2011, 71, 313–322. [Google Scholar] [CrossRef]
- Okoh, O.O.; Sadimenko, A.P.; Afolayan, A.J. Comoarative evaluation of the antibacterial activities of the essential oils of Rosmarinus officinalis L. obtained by hydrodistillation and solvent free microwave extraction methods. Food Chem. 2010, 120, 308–312. [Google Scholar] [CrossRef]
- Wanapat, M.; Cherdthong, A.; Pakdee, P.; Wanapat, S. Manipulation of rumen ecology by dietary lemongrass (Stapf.) powder supplementation. J. Anim. Sci. 2008, 86, 3497–3503. [Google Scholar] [CrossRef]
- Crespy, V.; Williamson, G. A review of the health effects of green tea catechins in in vivo animal models. J. Nutr. 2004, 134, 3431–3440. [Google Scholar] [CrossRef] [PubMed]
- Heleno, S.A.; Martins, A.; Queiroz, M.J.R.; Ferreira, I.C. Bioactivity of phenolic acids: Metabolites versus parent compounds: A review. Food Chem. 2015, 173, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Jouany, J.P.; Morgavi, D.P. Use of ‘natural’products as alternatives to antibiotic feed additives in ruminant production. Animals 2007, 1, 1443–1466. [Google Scholar]
- Belanche, A.; Newbold, C.J.; Morgavi, D.P.; Bach, A.; Zweifel, B.; Yáñez-Ruiz, D.R. A Meta-analysis Describing the Effects of the Essential oils Blend Agolin Ruminant on Performance, Rumen Fermentation and Methane Emissions in Dairy Cows. Animals 2020, 10, 620. [Google Scholar] [CrossRef] [PubMed]
- Kholif, A.E.; Olafadehan, O.A. Essential oils and phytogenic feed additives in ruminant diet: Chemistry, ruminal microbiota and fermentation, feed utilization and productive performance. Pharm. Rev. 2021, 1–22. [Google Scholar] [CrossRef]
- Busquet, M.; Calsamiglia, S.; Ferret, A.; Cardozo, P.W.; Kamel, C. Effects of cinnamaldehyde and garlic oil on rumen microbial fermentation in a dual flow continuous culture. J. Dairy Sci. 2005, 88, 2508–2516. [Google Scholar] [CrossRef]
- Busquet, M.; Calsamiglia, S.; Ferret, A.; Kamel, C. Plant extracts affect in vitro rumen microbial fermentation. J. Dairy Sci. 2006, 89, 761–771. [Google Scholar] [CrossRef]
- Garcia-Gonzales, R.; Dehority, B.A.; Lopez, S. Ruminal bacteria counts from in vitro cultures upon the addition of medicinal plants that modify the fermentation. In XI Jornadas sobre Produccion Animal; Gobierno de Aragón, Servicio de Investigación Agroalimentaria: Zaragoza, Spain, 2005; Volume I and II, pp. 614–616. [Google Scholar]
- Kobayashi, Y.; Oh, S.; Myint, H.; Koike, S. Use of Asian selected agricultural byproducts to modulate rumen microbes and fermentation. J. Ani. Sci. Biotechnol. 2016, 7, 1–10. [Google Scholar] [CrossRef]
- Konda, S.; Onodera, R.; Kanchanasatit, E.; Boonsaen, P.; Sawanon, S.; Nagashima, K.; Suzuki, Y.; Koike, S.; Kobayashi, Y. Effect of cashew nut shell liquid feeding on fermentation and microbiota in the rumen of Thai native cattle and swamp buffaloes. Livest. Sci. 2019, 226, 99–106. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Classification | Example |
|---|---|
| LAB | Lactobacilli |
| Enterococci | |
| LUB | Megasphaera elsdenii |
| Yeats | S. cerevisiae |
| Y. lipolytica | |
| Other | A. oryzae |
| A. niger |
| Prebiotic | Effect on Absorption Nutrients | Impact on Health |
|---|---|---|
| Fructooligosaccharides (FOS) | Improving feed efficiency, increasing yields | Positive effect on the immune system of calves and reduction of mortality rates. |
| Galactooligosaccharides (GOS) | Not fully known—divergent research results | Probably beneficial effects on LAB in the digestive system—however, further research is required |
| Mannanoligosaccharides (MOS) | There was an increase in the length of the rumen papillae and the height of the jejunum villi, which was probably related to the increase in substrate availability by MOS-utilizing bacteria. | Positive effect on health in calves—reduction of the intensity of diarrhea |
| Cellooligosaccharides (CO) | Acceleration of intestinal development, improves feed efficiency, increased calf growth. | Health and microbiological measurements do not change |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michalak, M.; Wojnarowski, K.; Cholewińska, P.; Szeligowska, N.; Bawej, M.; Pacoń, J. Selected Alternative Feed Additives Used to Manipulate the Rumen Microbiome. Animals 2021, 11, 1542. https://doi.org/10.3390/ani11061542
Michalak M, Wojnarowski K, Cholewińska P, Szeligowska N, Bawej M, Pacoń J. Selected Alternative Feed Additives Used to Manipulate the Rumen Microbiome. Animals. 2021; 11(6):1542. https://doi.org/10.3390/ani11061542
Chicago/Turabian StyleMichalak, Marta, Konrad Wojnarowski, Paulina Cholewińska, Natalia Szeligowska, Marcel Bawej, and Jakub Pacoń. 2021. "Selected Alternative Feed Additives Used to Manipulate the Rumen Microbiome" Animals 11, no. 6: 1542. https://doi.org/10.3390/ani11061542
APA StyleMichalak, M., Wojnarowski, K., Cholewińska, P., Szeligowska, N., Bawej, M., & Pacoń, J. (2021). Selected Alternative Feed Additives Used to Manipulate the Rumen Microbiome. Animals, 11(6), 1542. https://doi.org/10.3390/ani11061542