
Effect of Dietary Mineral Content and Phytase Dose on Nutrient Utilization, Performance, Egg Traits and Bone Mineralization in Laying Hens from 22 to 31 Weeks of Age


 ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Experimental Diets
2.3. Laying Performance and Egg Quality
2.4. Fecal and Ileal Digestibility
2.5. Bone Mineralization and Blood Analysis
2.6. Analytical Methods
2.7. Statistical Analyses
3. Results
4. Discussion
4.1. Nutrient Utilization
4.2. Laying Hens’ Performance and Egg Quality
4.3. Bone Mineralization
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lei, X.G.; Stahl, C.H. Biotechnological development of effective phytases for mineral nutrition and environmental protection. Appl. Microbiol. Biotechnol. 2001, 57, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Woyengo, T.A.; Nyachoti, C.M. Review: Anti-nutritional effects of phytic acid in diets for pigs and poultry—Current knowledge and directions for future research. Can. J. Anim. Sci. 2017, 93, 9–21. [Google Scholar] [CrossRef]
- Scott, M.L.; Hull, S.J.; Mullenhoff, P.A. The calcium requirements of laying hens and effects of dietary oyster shell upon egg shell quality. Poult. Sci. 1971, 50, 1055–1063. [Google Scholar] [CrossRef]
- Pelicia, K.; Garcia, E.A.; Faitarone, A.B.G.; Silva, A.P.; Berto, D.A.; Molino, A.B.; Vercese, F. Calcium and available phosphorus levels for laying hens in second production cycle. Braz. J. Poult. Sci. 2009, 11, 39–49. [Google Scholar] [CrossRef]
- Bougouin, A.; Appuhamy, J.A.; Kebreab, E.; Dijkstra, J.; Kwakkel, R.P.; France, J. Effects of phytase supplementation on phosphorus retention in broilers and layers: A meta-analysis. Poul. Sci. 2014, 93, 1981–1992. [Google Scholar] [CrossRef]
- Lim, H.S.; Namkung, H.; Paik, I.K. Effects of phytase supplementation on the performance, egg quality, and phosphorous excretion of laying hens fed different levels of dietary calcium and nonphytate phosphorous. Poult. Sci. 2003, 82, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Hughes, H.L.; Dahiya, J.P.; Wyatt, C.L.; Classen, H.L. Effect of Quantum phytase on nutrient digestibility and bone ash in white leghorn laying hens fed corn-soybean meal-based diets. Poult. Sci. 2009, 88, 1191–1198. [Google Scholar] [CrossRef]
- Selle, P.H.; Ravindran, V. Microbial phytase in poultry nutrition. Anim. Feed Sci. Technol. 2007, 135, 1–41. [Google Scholar] [CrossRef]
- Liebert, F.; Htoo, J.K.; Sunder, A. Performance and nutrient utilization of laying hens fed low-phosphorus corn–soybean and wheat–soybean diets supplemented with microbial phytase. Poult. Sci. 2005, 84, 1576–1583. [Google Scholar] [CrossRef]
- Boling, S.D.; Douglas, M.W.; Johnson, M.L.; Wang, X.; Parsons, C.M.; Koelkebeck, K.W.; Zimmerman, R.A. The effects of dietary available phosphorus levels and phytase on performance of young and older laying hens. Poult. Sci. 2000, 79, 224–230. [Google Scholar] [CrossRef]
- Francesch, M.; Broz, J.; Brufau, J. Effects of an experimental phytase on performance, egg quality, tibia ash content and phosphorus bioavailability in laying hens fed on maize- or barley-based diets. Br. Poult. Sci. 2005, 46, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Cowieson, A.J.; Wilcock, P.; Bedford, M.R. Super-dosing effects of phytase in poultry and other monogastrics. World’s Poult. Sci. J. 2011, 67, 225–235. [Google Scholar] [CrossRef]
- Cambra-López, M.; Cerisuelo, A.; Ferrer, P.; Ródenas, L.; Aligué, R.; Moset, V.; Pascual, J.J. Age influence on effectiveness of a novel 3-phytase in barley-wheat based diets for pigs from 12 to 108 kg under commercial conditions. Anim. Feed Sci. Technol. 2020, 267, 114549. [Google Scholar] [CrossRef]
- FEDNA (Fundación Española Desarrollo Nutrición Animal). Necesidades Nutricionales Para Avicutura, 2nd ed.; FEDNA: Madrid, Spain, 2010; ISBN 978-84-09-06529-5. Available online: http://www.fundacionfedna.org/node/75 (accessed on 8 April 2019).
- Haugh, R.R. The Haugh unit for measuring egg quality. U.S. Egg Poult. Mag. 1937, 43, 552–555. [Google Scholar]
- Adedokun, S.A.; Jaynes, P.; Abd El-Hack, M.E.; Payne, R.L.; Applegate, T.J. Standardized ileal amino acid digestibility of meat and bone meal and soybean meal in laying hens and broilers. Poult. Sci. 2014, 93, 420–428. [Google Scholar] [CrossRef]
- Rutherfurd, S.M.; Chung, T.K.; Morel, P.C.H.; Moughan, P.J. Effect of microbial phytase on ileal digestibility of phytate phosphorus, total phosphorus, and amino acids in a low-phosphorus diet for broilers. Poult. Sci. 2004, 83, 61–68. [Google Scholar] [CrossRef]
- Gao, C.Q.; Ji, C.; Zhang, J.Y.; Zhao, L.H.; Ma, Q.G. Effect of a novel plant phytase on performance, egg quality, apparent ileal nutrient digestibility and bone mineralization of laying hens fed corn–soybean diets. Anim. Feed Sci. Technol. 2013, 186, 101–105. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official’s Analytical Chemists, 21st ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2019. [Google Scholar]
- Haug, W.; Lantzch, H.J. Sensitive method for the rapid determination of phytate in cereals and cereal products. J. Sci. Food Agric. 1983, 34, 1423–1426. [Google Scholar] [CrossRef]
- Short, F.J.; Gorton, P.; Wiseman, J.; Boorman, K.N. Determination of titanium dioxide added as an inert marker in chicken digestibility studies. Anim. Feed Sci. Technol. 1996, 59, 215–221. [Google Scholar] [CrossRef]
- Littell, R.C.; Henry, P.R.; Ammerman, C.B. Statistical analysis of repeated measures data using SAS procedures. J. Anim. Sci. 1998, 76, 1216–1231. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 2, 292–305. [Google Scholar] [CrossRef]
- Mallin, M.A. Impacts of industrial animal production on rivers and estuaries. Am. Sci. 2000, 88, 26–37. [Google Scholar] [CrossRef]
- Lei, X.G.; Porres, J.M. Phytase enzymology application and biotechnology. Biotechnol. Lett. 2003, 25, 1787–1794. [Google Scholar] [CrossRef]
- Lei, X.G.; Porres, J.M.; Mullaney, E.J.; Brinch-Pedersen, H. Phytase source, structure and applications. In Industrial Enzymes. Structure, Function and Applications; Polaina, J., MacCabe, A.P., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 505–529. [Google Scholar]
- Sari, M.; Onol, A.G.; Daskiran, M.; Cengiz, O. Egg production and calcium-phosphorus utilization of laying hens fed diets supplemented with phytase alone or in combination with organic acid. Int. J. Poult. Sci. 2012, 11, 181–189. [Google Scholar] [CrossRef][Green Version]
- Meyer, E.; Parsons, C. The efficacy of a phytase enzyme fed to Hy-Line W-36 laying hens from 32 to 62 weeks of age. J. Appl. Poult. Res. 2011, 20, 136–142. [Google Scholar] [CrossRef]
- Ren, Z.; Sun, W.; Cheng, X.; Han, D.; Yan, J.; Pan, C.; Duan, Y.; Yang, X. The adaptability of Hy-Line Brown laying hens to low-phosphorus diets supplemented with phytase. Poult. Sci. 2020, 99, 3525–3531. [Google Scholar] [CrossRef]
- Pongmanee, K.; Kuhn, I.; Korver, D.R. Effects of phytase supplementation on eggshell and bone quality, and phosphorus and calcium digestibility in laying hens from 25 to 37 wk of age. Poult. Sci. 2020, 99, 2595–2607. [Google Scholar] [CrossRef] [PubMed]
- Bello, A.; Korver, D.R. Long-term effects of Buttiauxella sp. phytase on performance, eggshell quality, apparent ileal Ca and P digestibility, and bone properties of white egg layers. Poult. Sci. 2019, 98, 4848–4859. [Google Scholar] [CrossRef]
- Boorman, K.N.; Gunaratne, S.P. Dietary phosphorus supply, egg-shell deposition and plasma inorganic phosphorus in laying hens. Br. Poult. Sci. 2001, 42, 81–91. [Google Scholar] [CrossRef]
- Imari, Z.K.; Hassanabadi, A.; Moghaddam, H.N. Response of broiler chickens to calcium and phosphorus restriction: Effects on growth performance, carcase traits, tibia characteristics and total tract retention of nutrients. Ital. J. Anim. Sci. 2020, 19, 929–939. [Google Scholar] [CrossRef]
- Viveros, A.; Brenes, A.; Arija, I.; Centeno, C. Effects of microbial phytase supplementation on mineral utilization and serum enzyme activities in broiler chicks fed different levels of phosphorus. Poult. Sci. 2002, 81, 1172–1183. [Google Scholar] [CrossRef]
- Ahmadi, H.; Rodehutscord, M. A meta-analysis of responses to dietary nonphytate phosphorus and phytase in laying hens. Poult. Sci. 2012, 91, 2072–2078. [Google Scholar] [CrossRef] [PubMed]
- van der Klis, J.D.; Versteegh, H.A.J.; Simons, P.C.M.; Keis, A.K. The efficacy of phytase in corn-soybean meal-based diets for laying hens. Poult. Sci. 1997, 76, 1535–1542. [Google Scholar] [CrossRef] [PubMed]
- Elaroussi, M.A.; Forte, L.R.; Eber, S.L.; Biellier, H.V. Calcium homeostasis in the laying hen. 1. Age and dietary calcium effects. Poult. Sci. 1994, 73, 1581–1589. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; Kwakernaak, C. Comparative effects of two phytases versus increasing the inorganic phosphorus content of the diet, on nutrient and amino acid digestibility in boilers. Anim. Feed Sci. Technol. 2018, 253, 166–180. [Google Scholar] [CrossRef]
- Lei, Q.B.; Shi, L.X.; Zhang, X.Y.; Ding, X.M.; Bai, S.P.; Liu, Y.G. Effect of reduced energy, protein and entire substitution of inorganic phosphorus by phytase on performance and bone mineralisation of laying hens. Br. Poult. Sci. 2011, 52, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Scott, T.A.; Kampen, R.; Silvesides, F.G. The effect of phosphorus, phytase enzyme and calcium on the performance of layers fed corn-based diets. Poult. Sci. 1999, 78, 1742–1749. [Google Scholar] [CrossRef] [PubMed]
- Keshavarz, K. Nonphytate phosphorus requirement of laying hens with and without phytase on a phase feeding program. Poult Sci. 2000, 79, 748–763. [Google Scholar] [CrossRef]
- Hughes, A.L.; Dahiya, J.P.; Wyatt, C.L.; Classen, H.L. The efficacy of Quantum phytase in a forty-week production trial using white Leghorn laying hens fed corn-soybean meal-based diets. Poult. Sci. 2008, 87, 1156–1161. [Google Scholar] [CrossRef]
- Kozlowski, K.; Jeroch, H. Efficacy of different levels of Escherichia coli phytase in hens fed maize-soyabean meal-based diets with a decreased non-phytate phosphorus content. J. Anim. Feed Sci. 2011, 20, 224–235. [Google Scholar] [CrossRef]
- Hartel, H. Evaluation of the dietary interaction of calcium and phosphorus in the high producing laying hen. Br. Poult. Sci. 1990, 31, 473–494. [Google Scholar] [CrossRef] [PubMed]
- Troesch, B.; Jing, H.; Laillou, A.; Fowler, A. Absorption studies show that phytase from Aspergillus Niger significantly increases iron and zinc bioavailability from phytate-rich foods. Food Nutr. Bull. 2013, 34, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Gordon, R.W.; Roland, D.A. Performance of commercial laying hens fed various phosphorus levels with and without supplemental phytase. Poult. Sci. 1997, 76, 1172–1177. [Google Scholar] [CrossRef] [PubMed]
- Boling, S.D.; Douglas, M.W.; Shirley, R.B.; Parsons, C.M.; Koelkebeck, K.W. The effects of various dietary levels of phytase and available phosphorus on performance of laying hens. Poult. Sci. 2000, 79, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Keshavarz, K. The effect of different levels of nonphytate phosphorus with and without phytase on the performance of four strains of laying hens. Poult. Sci. 2003, 82, 71–91. [Google Scholar] [CrossRef]
- Keshavarz, K. A comparison between cholecalciferol and 25-OH-cholecalciferol on performance and eggshell quality of hens fed different levels of calcium and phosphorus. Poult. Sci. 2003, 82, 1415–1422. [Google Scholar] [CrossRef]
- Jalal, M.A.; Scheideler, S.E. Effect of supplementation of two different sources of phytase on egg production parameters in laying hens and nutrient digestibility. Poult. Sci. 2001, 80, 1463–1471. [Google Scholar] [CrossRef]
- Scott, T.A.; Kampen, R.; Silversides, F.G. The effect of adding exogenous phytase to nutrient reduced corn and wheat-based diets on performance and egg quality of two strains of laying hens. Can. J. Anim. Sci. 2001, 81, 393–401. [Google Scholar] [CrossRef]
- Punna, S.; Roland, D.A., Sr. Influence of supplemental microbial phytase on first cycle laying hens fed phosphorus-deficient diets from day one of age. Poult. Sci. 1999, 78, 1407–1411. [Google Scholar] [CrossRef]
- Murakami, F.S.; Rodrigues, P.O.; de Campos, C.M.T.; Silva, M.A.S. Physicochemical study of CaCO3 from eggshells. Food Sci. Technol. 2007, 27, 658–662. [Google Scholar] [CrossRef]
- Rodríguez-Navarro, C.; Kudłacz, K.; Cizer, Ö.; Ruiz-Agudo, E. Formation of amorphous calcium carbonate and its transformation into mesostructured calcite. CrystEngComm 2015, 17, 58–72. [Google Scholar] [CrossRef]
- Bar, A.; Razaphkovsky, V.; Vax, E. Re-evaluation of calcium and phosphorus requirements in aged laying hens. Br. Poult. Sci. 2002, 43, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Yan, J.K.; Sun, W.Q.; Chen, Z.Y.; Wu, S.R.; Ren, Z.Z.; Yang, X.J. Effect of inorganic phosphate supplementation on egg production in Hy-Line Brown layers fed 2000 FTU/kg phytase. Animal 2020, 14, 2246–2252. [Google Scholar] [CrossRef]
- Jang, H.D.; Hyun, Y.; Kim, H.S.; Hwang, I.W.; Yoo, J.S.; Kim, H.J.; Shin, S.O.; Hwang, Y.; Zhou, T.X.; Chen, Y.J.; et al. Effect of dietary microbial phytase on laying performance, egg quality, phosphorus utilization and nutrient metabolizability in laying hens. Korean J. Poult. Sci. 2008, 35, 115–121. [Google Scholar] [CrossRef][Green Version]
- Englmaierová, M.; Dlouhá, G.; Marounek, M.; Skřivan, M. Efficacy of contrast levels of non-phytate phosphorus and Aspergillus niger phytase in hens fed wheat-maize-based diets. Czech J. Anim. Sci. 2012, 57, 499–505. [Google Scholar] [CrossRef]
- Brunelli, S.R.; Pinheiro, J.W. Effect of different inclusion levels of defatted corn germ meal in diets supplemented with phytase for laying hens. Semin. Cienc. Agrar. 2012, 33, 1991–2000. [Google Scholar] [CrossRef]
- Millán-Calleja, C.; Gracia, M.I.; Dersjant-Li, Y.; Debicki-Garnier, A.M.; Messager, B. Efficacy of a phytase issued from Buttiauxella sp. on performance of laying hens from 21 to 57 weeks of age. In Proceedings of the 12ème Journees de la Recherche Avicole et Palmipèdes à Foie Gras, Tours, France, 5–6 April 2017; pp. 622–626. [Google Scholar]
- Gardoni, B.L.K.; Sato, M.E.O.; Pontarolo, R.; Noronha, L.; Reichert, A.; Serafini, S.Z. Avaliação clínica e morfológica da ação da hidroquinona e do ácido fítico como agentes despigmentantes. Acta Farm. Banaerense 2004, 23, 297–303. [Google Scholar]


{kind=link}
| Ingredients and Chemical Composition | Positive Control | Negative Control |
|---|---|---|
| Ingredients | ||
| Corn grain | 592.5 | 610 |
| Soybean meal 44% CP | 276 | 276 |
| Soybean meal oil | 25.4 | 25.4 |
| DL-Methionine | 1.6 | 1.6 |
| Calcium carbonate | 80.0 | 75.0 |
| Dicalcium phosphate | 16.0 | 3.5 |
| Salt | 3.45 | 3.45 |
| Red coloring (canthaxanthin 10%) | 0.05 | 0.05 |
| Vit-min premix 1 | 5 | 5 |
| Chemical composition | ||
| Dry matter | 899 | 900 |
| Ash | 118 | 111 |
| Crude protein | 170 | 172 |
| Ether extract | 44.1 | 44.4 |
| Gross energy (kcal/kg) | 3652 | 3815 |
| Apparent metabolizable energy (kcal/kg) | 2819 | 2913 |
| Metabolizable protein | 90 | 87 |
| Calcium | 40.7 | 29.7 |
| Total phosphorus | 6.1 | 3.7 |
| Phytate phosphorus | 1.7 | 1.7 |
| Non-phytate phosphorus 2 | 4.4 | 2.0 |
| Ca | P | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| DM | OM | CP | GE | CTTAD | Ca Ret | Ca Exc | CTTAD | P Ret | P Exc | |
| 25 weeks of age | ||||||||||
| PC | 71.3 b | 74.9 a | 50.4 b | 76.8 a | 57.3 b | 2.54 a | 42.7 a | 20.8 b | 0.134 a | 79.2 a |
| NC | 71.8 b | 73.4 b | 46.0 c | 75.9 b | 63.6 a | 2.05 b | 36.4 b | 24.2 b | 0.089 b | 75.8 a |
| P500 | 72.3 ab | 74.7 a | 51.9 ab | 76.7 ab | 63.5 a | 2.44 a | 36.5 b | 30.1 a | 0.128 a | 69.9 b |
| P1000 | 72.8 a | 75.1 a | 53.0 a | 76.7 ab | 65.2 a | 2.05 b | 34.8 b | 24.9 b | 0.101 b | 75.1 a |
| SEM | 0.36 | 0.331 | 0.761 | 0.341 | 1.52 | 0.082 | 1.52 | 1.65 | 0.011 | 1.65 |
| 31 weeks of age | ||||||||||
| PC | 71.8 b | 75.9 a | 55.2 a | 77.6 a | 58.3 c | 2.68 a | 41.7 a | 10.1 c | 0.058 c | 90.9 a |
| NC | 72.7 b | 74.6 b | 55.0 a | 76.8 ab | 67.3 b | 2.62 ab | 32.7 bc | 31.7 b | 0.129 b | 68.3 b |
| P500 | 72.2 b | 74.6 b | 54.1 ab | 75.3 c | 64.6 b | 2.50 ab | 35.4 b | 29.9 b | 0.120 b | 70.1 b |
| P1000 | 73.9 a | 76.2 a | 52.2 b | 76.5 b | 70.6 a | 2.47 b | 29.4 c | 38.9 a | 0.179 a | 61.1 c |
| SEM | 0.445 | 0.348 | 0.994 | 0.389 | 1.66 | 0.086 | 1.66 | 2.15 | 0.011 | 2.19 |
| p-value | ||||||||||
| Diet | <0.001 | <0.001 | 0.02 | 0.002 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Age | 0.014 | <0.001 | <0.001 | 0.856 | 0.008 | <0.001 | 0.008 | 0.052 | 0.208 | 0.077 |
| Diet x Age | 0.344 | 0.117 | <0.001 | 0.002 | 0.366 | 0.006 | 0.366 | <0.001 | <0.001 | <0.001 |
| Traits | PC | NC | P500 | P1000 | SEM | p-Value |
|---|---|---|---|---|---|---|
| AID, % | ||||||
| Dry matter 1,2 | 68.24 b | 68.82 b | 66.58 b | 76.50 a | 1.37 | <0.001 |
| Ca | 42.70 c | 59.89 ab | 53.14 bc | 65.60 a | 4.27 | 0.003 |
| P 1 | 19.89 b | 22.91 b | 29.67 b | 52.96 a | 3.52 | <0.001 |
| Blood concentration, mg/dL | ||||||
| Ca | 30.47 a | 28.35 b | 28.01 b | 27.87 b | 0.721 | 0.043 |
| P | 7.35 a | 5.38 b | 5.56 b | 5.98 b | 0.368 | 0.002 |
| Traits | PC | NC | P500 | P1000 | SEM | p-Value |
|---|---|---|---|---|---|---|
| Initial body weight, g | 1749 | 1779 | 1753 | 1747 | 0.024 | 0.759 |
| Final body weight, g | 1872 ab | 1851 b | 1901 a | 1840 b | 0.017 | 0.074 |
| ADFI, g/day | 109.2 | 108 | 110.5 | 105.2 | 1.77 | 0.176 |
| FCR, g feed/g egg | 1.849 a | 1.799 ab | 1.830 ab | 1.758 b | 0.031 | 0.187 |
| Average laying index | 0.973 | 0.975 | 0.978 | 0.98 | 0.044 | 0.528 |
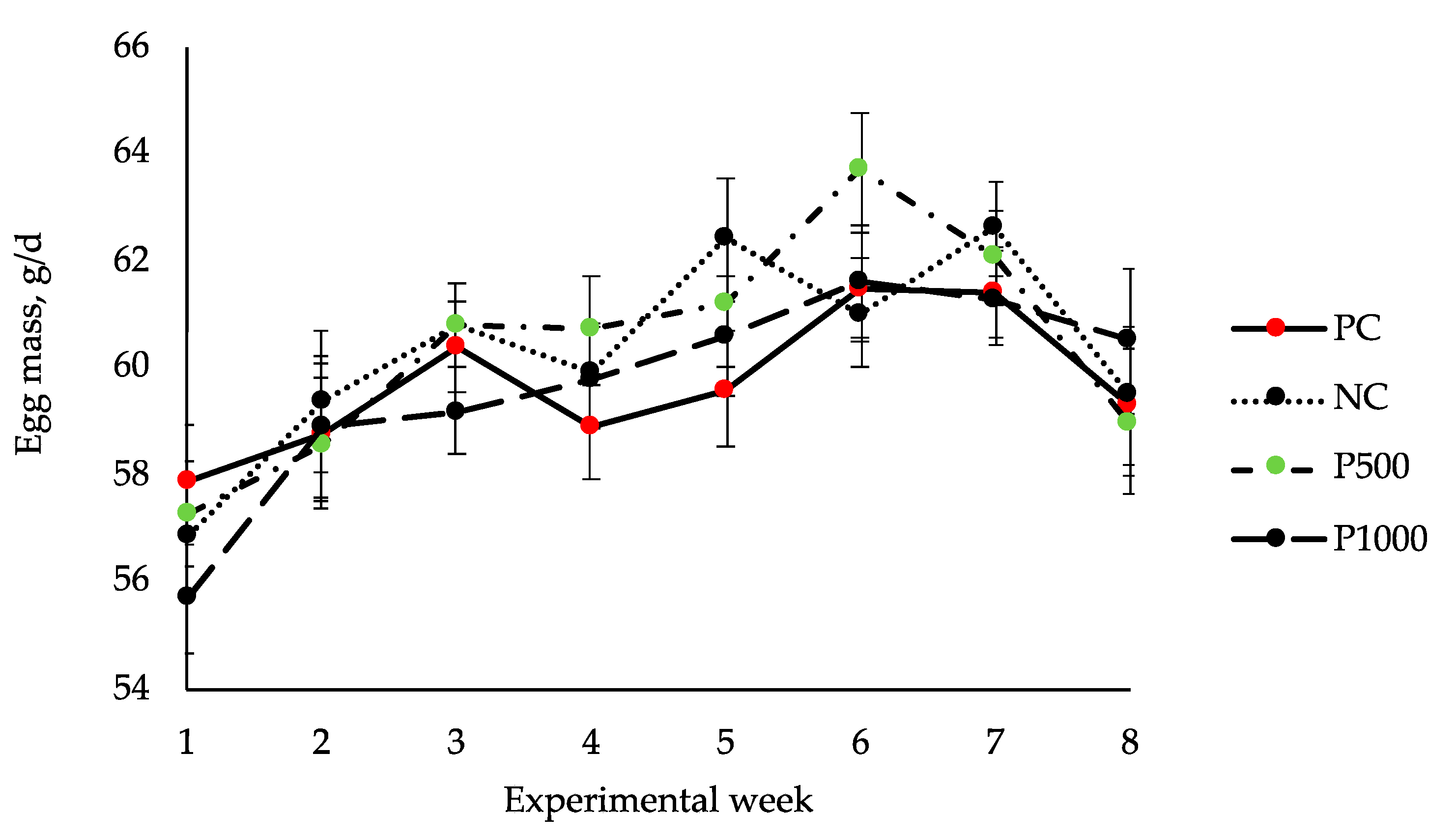
| Average egg mass, g/day | 59.28 | 60.56 | 59.66 | 59.84 | 0.696 | 0.594 |
| Egg traits at 31 weeks of age: | ||||||
| Shell thickness, mm 1,2 | 0.382 a | 0.371 b | 0.375 ab | 0.359 c | 0.003 | <0.001 |
| Albumen height, mm | 11.46 | 11.24 | 11.44 | 11.37 | 0.152 | 0.725 |
| Haugh units | 104.3 | 103.4 | 104.2 | 104.2 | 0.595 | 0.696 |
| Yolk color 1,3 | 13.81 a | 13.57 b | 13.80 a | 13.95 a | 0.059 | <0.001 |
| Traits | PC | NC | P500 | P1000 | SEM | p-Value |
|---|---|---|---|---|---|---|
| Tibia weight, g | 6 | 5.96 | 5.81 | 5.81 | 0.197 | 0.845 |
| Tibia weight, % BW | 0.322 a | 0.308 ab | 0.301 b | 0.299 b | 0.007 | 0.113 |
| Ash in tibia (% DM) | 52.0 a | 49.8 b | 49.6 b | 50.8 ab | 0.463 | 0.001 |
| Ca in tibia (% DM) | 18.72 a | 18.09 b | 17.92 b | 18.29 ab | 0.235 | 0.064 |
| P in tibia (% DM) | 8.78 a | 8.39 b | 8.38 b | 8.51 b | 0.085 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Javadi, M.; Pascual, J.J.; Cambra-López, M.; Macías-Vidal, J.; Donadeu, A.; Dupuy, J.; Carpintero, L.; Ferrer, P.; Cerisuelo, A. Effect of Dietary Mineral Content and Phytase Dose on Nutrient Utilization, Performance, Egg Traits and Bone Mineralization in Laying Hens from 22 to 31 Weeks of Age. Animals 2021, 11, 1495. https://doi.org/10.3390/ani11061495
Javadi M, Pascual JJ, Cambra-López M, Macías-Vidal J, Donadeu A, Dupuy J, Carpintero L, Ferrer P, Cerisuelo A. Effect of Dietary Mineral Content and Phytase Dose on Nutrient Utilization, Performance, Egg Traits and Bone Mineralization in Laying Hens from 22 to 31 Weeks of Age. Animals. 2021; 11(6):1495. https://doi.org/10.3390/ani11061495
Chicago/Turabian StyleJavadi, Mehran, Juan José Pascual, María Cambra-López, Judit Macías-Vidal, Andrés Donadeu, Javier Dupuy, Laura Carpintero, Pablo Ferrer, and Alba Cerisuelo. 2021. "Effect of Dietary Mineral Content and Phytase Dose on Nutrient Utilization, Performance, Egg Traits and Bone Mineralization in Laying Hens from 22 to 31 Weeks of Age" Animals 11, no. 6: 1495. https://doi.org/10.3390/ani11061495
APA StyleJavadi, M., Pascual, J. J., Cambra-López, M., Macías-Vidal, J., Donadeu, A., Dupuy, J., Carpintero, L., Ferrer, P., & Cerisuelo, A. (2021). Effect of Dietary Mineral Content and Phytase Dose on Nutrient Utilization, Performance, Egg Traits and Bone Mineralization in Laying Hens from 22 to 31 Weeks of Age. Animals, 11(6), 1495. https://doi.org/10.3390/ani11061495