
Bovine Intelectin 2 Expression as a Biomarker of Paratuberculosis Disease Progression

 , ,
, ,  ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animals and Samples
2.3. Serological, Bacteriological and Molecular Characterization of the Animals
2.4. Histopathological Classification of Animals
2.5. Immunohistochemistry (IHC) and Image Analysis
2.6. Statistical Analysis
3. Results
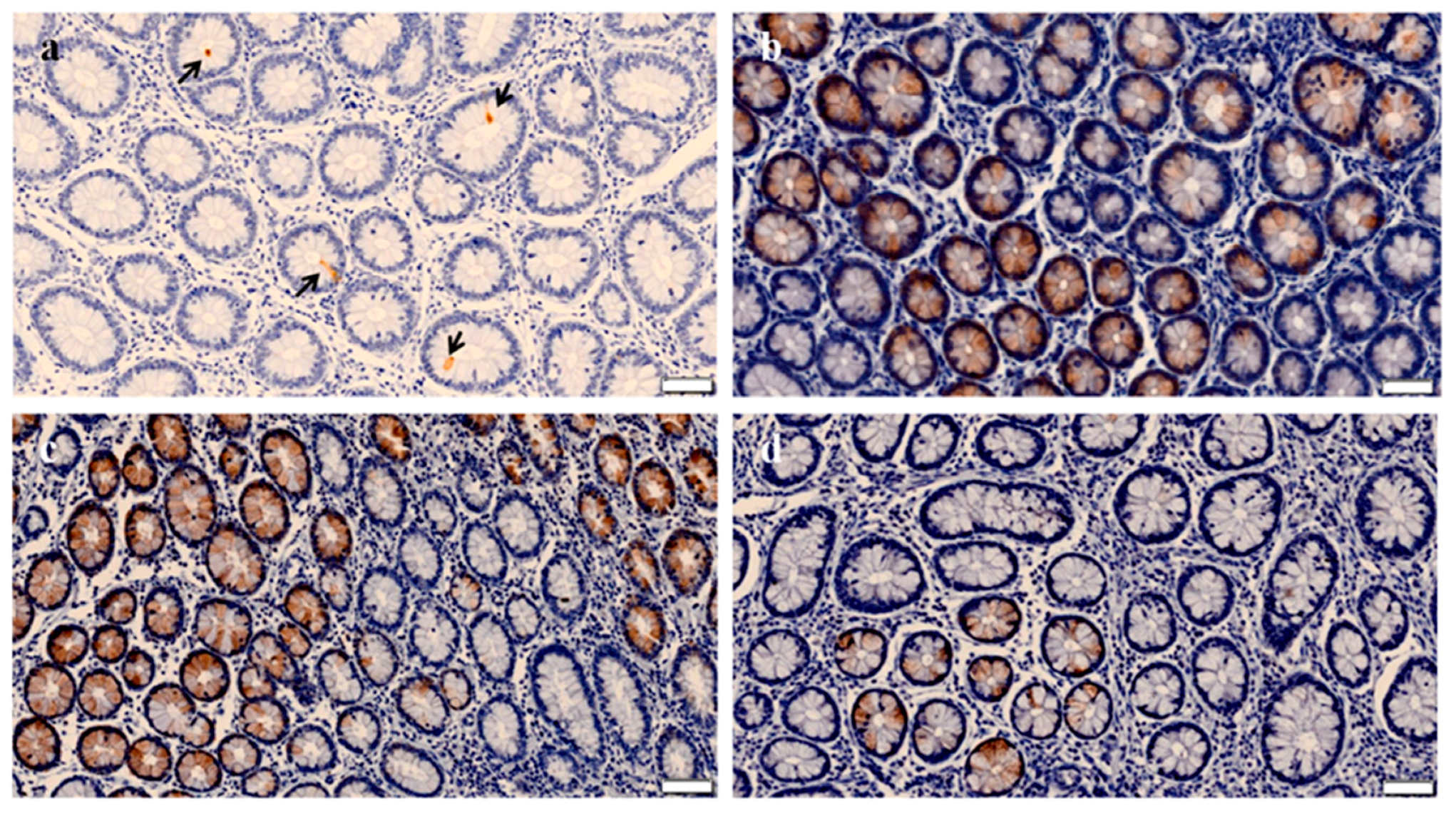
3.1. Quantitative Immunohistochemical Evaluation of ITLN2 Secreting Cells within the Different PTB Histopathological Types
3.2. ROC Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benedictus, G.; Dijkhuizen, A.A.; Stelwagen, J. Economic losses due to paratuberculosis in dairy cattle. Vet. Rec. 1987, 121, 142–146. [Google Scholar] [CrossRef]
- Kudahl, A.B.; Nielsen, S.S.; Sørensen, J.T. Relationship between antibodies against Mycobacterium avium subsp. paratuberculosis in milk and shape of lactation curves. Pre. Vet. Med. 2004, 62, 119–134. [Google Scholar] [CrossRef]
- Feller, M.; Huwiler, K.; Stephan, R.; Altpeter, E.; Shang, A.; Furrer, H.; Pfyffer, G.E.; Jemmi, T.; Baumgartner, A.; Egger, M. Mycobacterium avium subspecies paratuberculosis and Crohn’s disease: A systematic review and meta-analysis. Lancet Infect. Dis. 2007, 7, 607–613. [Google Scholar] [CrossRef]
- Kuenstner, J.T.; Naser, S.; Chamberlin, W.; Borody, T.; Graham, D.Y.; McNees, A.; Hermon-Taylor, J.; Hermon-Taylor, A.; Dow, C.T.; Thayer, W.; et al. The consensus from the Mycobacterium avium ssp. paratuberculosis (MAP) Conference 2017. Front Public Health 2017, 5, 208. [Google Scholar] [CrossRef] [PubMed]
- Bo, M.; Erre, G.L.; Niegowska, M.; Piras, M.; Taras, L.; Longu, M.G.; Passiu, G.; Sechi, L.A. Interferon regulatory factor is a potential target of autoimmune response triggered by Epstein-Barr virus and Mycobacterium avium subsp. paratuberculosis in rheumatoid arthritis: Investigating a mechanism of molecular mimicry. Clin. Exp. Rheumatol. 2018, 36, 376–381. [Google Scholar]
- Bo, M.; Niegowska, M.; Erre, G.L.; Piras, M.; Longu, M.G.; Manchia, P.; Manca, M.; Passiu, G.; Sechi, L.A. Rheumatoid arthritis patient antibodies highly recognize IL-2 in the immune response pathway involving IRF5 and EBV antigens. Sci. Rep. 2018, 8, 1789. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, K.; Cossu, D.; Hoshino, Y.; Tomizawa, Y.; Momotani, E.; Hattori, N. Anti-mycobacterial antibodies in paired cerebrospinal fluid and serum samples from Japanese patients with multiple sclerosis or neuromyelitis optica spectrum disorder. J. Clin. Med. 2018, 7, 522. [Google Scholar] [CrossRef] [PubMed]
- Mameli, G.; Cocco, E.; Frau, J.; Marrosu, M.G.; Sechi, L.A. Epstein-Barr virus and Mycobacterium avium subsp. paratuberculosis peptides are recognized in sera and cerebrospinal fluid of MS patients. Sci. Rep. 2016, 6, 22401. [Google Scholar] [CrossRef]
- Niegowska, M.; Rapini, N.; Piccini, S.; Mameli, G.; Caggiu, E.; Manca Bitti, M.L.; Sechi, L.A. Type 1 diabetes at-risk children highly recognize Mycobacterium avium subspecies paratuberculosis epitopes homologous to human Znt8 and proinsulin. Sci. Rep. 2016, 6, 22266. [Google Scholar] [CrossRef] [PubMed]
- Bastida, F.; Juste, R.A. Paratuberculosis control: A review with a focus on vaccination. J. Immune Based Ther. Vaccines 2011, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Groenendaal, H.; Nielen, M.; Jalvingh, A.W.; Horst, S.H.; Galligan, D.T.; Hesselink, J.W. A simulation of Johne’s disease control. Prev. Vet. Med. 2002, 54, 225–245. [Google Scholar] [CrossRef]
- Garry, F. Control of paratuberculosis in dairy herds. Vet. Clin. North Am. Food Anim. Pract. 2011, 27, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Kudahl, A.B.; Nielsen, S.S.; Ostergaard, S. Strategies for time of culling in control of paratuberculosis in dairy herds. J. Dairy Sci. 2011, 94, 3824–3834. [Google Scholar] [CrossRef]
- Nielsen, S.S.; Toft, N. Age-specific characteristics of ELISA and fecal culture for purpose-specific testing for paratuberculosis. J. Dairy Sci. 2006, 89, 569–579. [Google Scholar] [CrossRef]
- Sevilla, I.A.; Garrido, J.M.; Molina, E.; Geijo, M.V.; Elguezabal, N.; Vázquez, P.; Juste, R.A. Development and evaluation of a novel multicopy-element-targeting triplex PCR for detection of Mycobacterium avium subsp. paratuberculosis in feces. Appl. Environ. Microbiol. 2014, 80, 3757–3768. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wells, S.J.; Collins, M.T.; Faaberg, K.S.; Wees, C.; Tavornpanich, S.; Petrini, K.R.; Collins, J.E.; Cernicchiaro, N.; Whitlock, R.H. Evaluation of a rapid fecal PCR test for detection of Mycobacterium avium subsp. paratuberculosis in dairy cattle. Clin. Vaccine Immunol. 2006, 13, 1125–1130. [Google Scholar] [CrossRef]
- Nielsen, S.S.; Toft, N. Ante mortem diagnosis of paratuberculosis: A review of accuracies of ELISA, interferon-γ assay and faecal culture techniques. Vet. Microbiol. 2008, 129, 217–235. [Google Scholar] [CrossRef]
- Capsel, R.T.; Thoen, C.O.; Reinhardt, T.A.; Lippolis, J.D.; Olsen, R.; Stabel, J.R.; Bannantine, J.P. Composition and potency characterization of Mycobacterium avium subsp. paratuberculosis purifed protein derivatives. PLoS ONE 2016, 11, 1–19. [Google Scholar] [CrossRef]
- Seth, M.; Lamont, E.A.; Janagama, H.K.; Widdel, A.; Vulchanova, L.; Stabel, J.R.; Waters, W.R.; Palmer, M.V.; Sreevatsan, S. Biomarker discovery in subclinical mycobacterial infections of cattle. PLoS ONE 2009, 4, e5478. [Google Scholar] [CrossRef]
- You, Q.; Verschoor, C.P.; Pant, S.D.; Macri, J.; Kirby, G.M.; Karrow, N.A. Proteomic analysis of plasma from Holstein cows testing positive for Mycobacterium avium subsp. paratuberculosis (MAP). Vet. Immunol. Immunopathol. 2012, 148, 243–251. [Google Scholar] [CrossRef]
- Cha, S.B.; Yoo, A.; Park, H.T.; Sung, K.Y.; Shin, M.K.; Yoo, H.S. Analysis of transcriptional profiles to discover biomarker candidates in Mycobacterium avium subsp. paratuberculosis-infected macrophages, RAW 264.7. J. Microbiol. Biotechnol. 2013, 23, 1167–1175. [Google Scholar] [CrossRef]
- David, J.; Barkema, H.W.; Guan, L.L.; De Buck, J. Gene-expression profiling of calves 6 and 9 months after inoculation with Mycobacterium avium subsp. paratuberculosis. Vet. Res. 2014, 45, 96. [Google Scholar] [CrossRef]
- Shin, M.K.; Park, H.T.; Shin, S.W.; Jung, M.; Im, Y.B.; Park, H.E.; Cho, Y.I.; Yoo, H.S. Whole-blood gene-expression profiles of cows infected with Mycobacterium avium subsp. paratuberculosis reveal changes in immune response and lipid metabolism. J. Microbiol. Biotechnol. 2015, 25, 255–267. [Google Scholar] [CrossRef]
- Shin, M.K.; Park, H.; Shin, S.W.; Jung, M.; Lee, S.H.; Kim, D.Y.; Yoo, H.S. Host transcriptional profiles and immunopathologic response following Mycobacterium avium subsp. paratuberculosis infection in mice. PLoS ONE 2015, 10, e0138770. [Google Scholar] [CrossRef]
- Hempel, R.J.; Bannantine, J.P.; Stabel, J.R. Transcriptional profiling of ileocecal valve of Holstein dairy cows infected with Mycobacterium avium subsp. paratuberculosis. PLoS ONE 2016, 11, e0153932. [Google Scholar] [CrossRef]
- Malvisi, M.; Palazzo, F.; Morandi, N.; Lazzari, B.; Williams, J.L.; Pagnacco, G.; Minozzi, G. Responses of bovine innate immunity to Mycobacterium avium subsp. paratuberculosis infection revealed by changes in gene expression and levels of microRNA. PLoS ONE 2016, 11, e0164461. [Google Scholar] [CrossRef]
- Van den Esker, M.H.; Koets, A.P. Application of transcriptomics to enhance early diagnostics of mycobacterial infections, with an emphasis on Mycobacterium avium ssp. paratuberculosis. Vet. Sci. 2019, 6, 59. [Google Scholar] [CrossRef] [PubMed]
- Palacios, A.; Sampedro, L.; Sevilla, I.A.; Molina, E.; Gil, D.; Azkargorta, M.; Elortza, F.; Garrido, J.M.; Anguita, J.; Prados-Rosales, R. Mycobacterium tuberculosis extracelular vesicle-associated lipoprotein LpqH as a potential biomarker to distinguish paratuberculosis infection or vaccination from tuberculosis infection. BMC Vet. Res. 2019, 15, 188. [Google Scholar] [CrossRef]
- Purdie, A.C.; Plain, K.M.; Begg, D.J.; de Silva, K.; Whittington, R.J. Gene expression profiles during subclinical Mycobacterium avium subspecies paratuberculosis infection in sheep can predict disease outcome. Sci. Rep. 2019, 9, 8245. [Google Scholar] [CrossRef] [PubMed]
- Shaughnessy, R.G.; Farrell, D.; Stojkovic, B.; Browne, J.A.; Kenny, K.; Gordon, S.V. Identification of microRNAs in bovine faeces and their potential as biomarkers of Johne’s Disease. Sci. Rep. 2020, 10, 5908. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Hearn, M.; Canive, M.; Blanco-Vázquez, C.; Torremocha, R.; Balseiro, A.; Amado, J.; Varela-Martínez, E.; Ramos, R.; Jugo, B.M.; Casais, R. RNA-Seq analysis of ileocecal valve and peripheral blood from Holstein cattle infected with Mycobacterium avium subsp. paratuberculosis revealed dysregulation of the CXCL8/IL8 signaling pathway. Sci. Rep. 2019, 9, 14845. [Google Scholar] [CrossRef]
- Blanco Vázquez, C.; Alonso-Hearn, M.; Juste, R.A.; Canive, M.; Iglesias, T.; Iglesias, N.; Amado, J.; Vicente, F.; Balseiro, A.; Casais, R. Detection of latent forms of Mycobacterium avium subsp. paratuberculosis infection using host biomarker-based ELISAs greatly improves paratuberculosis diagnostic sensitivity. PLoS ONE 2020, 15, e0236336. [Google Scholar] [CrossRef] [PubMed]
- Arranz-Plaza, E.; Tracy, A.S.; Siriwardena, A.; Pierce, J.M.; Boons, G.J. High-avidity, low-affinity multivalent interactions and the block to polyspermy in Xenopus laevis. J. Am. Chem. Soc. 2002, 124, 13035–13046. [Google Scholar] [CrossRef]
- Tsuji, S.; Uehori, J.; Matsumoto, M.; Suzuki, Y.; Matsuhisa, A.; Toyoshima, K.; Seya, T. Human intelectin is a novel soluble lectin that recognizes galactofuranose in carbohydrate chains of bacterial cell wall. J. Biol. Chem. 2001, 276, 23456–23463. [Google Scholar] [CrossRef]
- Dann, S.M.; Eckmann, L. Innate immune defenses in the intestinal tract. Curr. Opin. Gastroenterol. 2007, 32, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Abe, Y.; Tokuda, M.; Ishimoto, R.; Azumi, K.; Yokosawa, H. A unique primary structure, cDNA cloning and function of a galactose-specific lectin from ascidian plasma. Eur. J. Biochem. 1999, 261, 33–39. [Google Scholar] [CrossRef]
- Pemberton, A.D.; Knight, P.A.; Gamble, J.; Colledge, W.H.; Lee, J.K.; Pierce, M.; Miller, H.R. Innate BALB/c enteric epithelial responses to Trichinella spiralis: Inducible expression of a novel goblet cell lectin, intelectin-2, and its natural deletion in C57BL/10 mice. J. Immunol. 2004, 173, 1894–1901. [Google Scholar] [CrossRef] [PubMed]
- Datta, R.; deSchoolmeester, M.L.; Hedeler, C.; Paton, N.W.; Brass, A.M.; Else, K.J. Identification of novel genes in intestinal tissue that are regulated after infection with an intestinal nematode parasite. Infect. Immun. 2005, 73, 4025–4033. [Google Scholar] [CrossRef]
- Chang, M.X.; Nie, P. Intelectin gene from the grass carp Ctenopharyngodon idella: cDNA cloning, tissue expression, and immunohistochemical localization. Fish Shellfish Immunol. 2007, 23, 128–140. [Google Scholar] [CrossRef]
- French, A.T.; Knight, P.A.; Smith, W.D.; Brown, J.K.; Craig, N.M.; Pate, J.A.; Miller, H.R.; Pemberton, A.D. Up-regulation of intelectin in sheep after infection with Teladorsagia circumcincta. Int. J. Parasitol. 2008, 38, 467–475. [Google Scholar] [CrossRef]
- Takano, T.; Sha, Z.; Peatman, E.; Terhune, J.; Liu, H.; Kucuktas, H.; Li, P.; Edholm, E.S.; Wilson, M.; Liu, Z. The two channel catfish intelectin genes exhibit highly differential patterns of tissue expression and regulation after infection with Edwardsiella ictaluri. Dev. Comp. Immunol. 2008, 32, 693–705. [Google Scholar] [CrossRef]
- Kuperman, D.A.; Lewis, C.C.; Woodruff, P.G.; Rodriguez, M.W.; Yang, Y.H.; Dolganov, G.M.; Fahy, J.V.; Erle, D.J. Dissecting asthma using focused transgenic modeling and functional genomics. J. Allergy Clin. Immunol. 2005, 116, 305–311. [Google Scholar] [CrossRef]
- Kilpatrick, D.C. Animal lectins: A historical introduction and overview. Biochim. Biophys. Acta. 2002, 1572, 187–197. [Google Scholar] [CrossRef]
- Tosi, M.F. Innate immune responses to infection. J. Allergy Clin. Immunol. 2005, 116, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wang, J.F.; Zhao, Y.Q.; Zhang, J.Y.; Bai, C.C.; Zhang, C.Q.; Zhang, C.; Li, K.; Zhang, H.; Du, X.; et al. Identification of an amphioxus intelectin homolog that preferably agglutinates grampositive over gram-negative bacteria likely due to different binding capacity to LPS and PGN. Fish Shellfish Immunol. 2012, 33, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhang, C.; Zhang, Y.; Li, K.; Xu, L.; Guo, L.; Kong, Y.; Feng, L. Characterization and comparative analyses of two amphioxus intelectins involved in the innate immune response. Fish Shellfish Immunol. 2013, 34, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Yonekawa, A.; Saijo, S.; Hoshino, Y.; Miyake, Y.; Ishikawa, E.; Suzukawa, M.; Inoue, H.; Tanaka, M.; Yoneyama, M.; Oh-Hora, M.; et al. Dectin-2 is a direct receptor for mannose-capped lipoarabinomannan of mycobacteria. Immunity 2014, 41, 402–413. [Google Scholar] [CrossRef]
- Komiya, T.; Tanigawa, Y.; Hirohashi, S. Cloning of the novel gene intelectin, which is expressed in intestinal paneth cells in mice. Biochem. Biophys. Res. Commun. 1998, 251, 759–762. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Baum, L.G.; Moremen, K.; Pierce, M. The X-lectins: A new family with homology to the Xenopus laevis oocyte lectin XL-35. Glycoconj. J. 2004, 21, 443–450. [Google Scholar] [CrossRef]
- Peatman, E.; Baoprasertkul, P.; Terhune, J.; Xu, P.; Nandi, S.; Kucuktas, H.; Li, P.; Wang, S.; Somridhivej, B.; Dunham, R.; et al. Expression analysis of the acute phase response in channel catfish (Ictalurus punctatus) after infection with a Gram-negative bacterium. Dev. Comp. Immunol. 2007, 31, 1183–1196. [Google Scholar] [CrossRef]
- Russell, S.; Young, K.M.; Smith, M.; Hayes, M.A.; Lumsden, J.S. Identification, cloning and tissue localization of a rainbow trout (Oncorhynchus mykiss) intelectin-like protein that binds bacteria and chitin. Fish Shellfish Immunol. 2008, 25, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Cao, Z.; Su, P.; Zhang, H.B.; Li, M.Z.; Lin, Y.Q.; Zhao, D.; Shen, Y.; Jinj, C.; Chen, S.; et al. Characterization and comparative analyses of zebrafish intelectins: Highly conserved sequences, diversified structures and functions. Fish Shellfish Immunol. 2009, 26, 396–405. [Google Scholar] [CrossRef]
- Xue, Z.; Pang, Y.; Liu, X.; Zheng, Z.; Xiao, R.; Jin, M.; Han, Y.; Su, P.; Lv, L.; Wang, J.; et al. First evidence of protein G-binding protein in the most primitive vertebrate: Serum lectin from lamprey (Lampetra japonica). Dev. Comp. Immunol. 2013, 41, 618–630. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Schnee, J.; Pang, M.; Wolfert, M.; Baum, L.G.; Moremen, K.W.; Pierce, M. Human homologs of the Xenopus oocyte cortical granule lectin XL35. Glycobiology 2001, 11, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Cloix, L.; Reverchon, M.; Cornuau, M.; Froment, P.; Rame, C.; Costa, C.; Froment, G.; Lecomte, P.; Chen, W.; Royère, D.; et al. Expression and regulation of INTELECTIN1 in human granulosa-lutein cells: Role in IGF-1-induced steroidogenesis through NAMPT. Biol. Reprod. 2014, 91, 50. [Google Scholar] [CrossRef]
- Wrackmeyer, U.; Hansen, G.H.; Seya, T.; Danielsen, E.M. Intelectin: A novel lipid raft-associated protein in the enterocyte brush border. Biochemistry 2006, 45, 9188–9197. [Google Scholar] [CrossRef] [PubMed]
- Peebles, R.S. The intelectins: A new link between the immune response to parasitic infections and allergic inflammation? Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 298, L288–L289. [Google Scholar] [CrossRef]
- Suzuki, Y.A.; López, V.; Lonnerdal, B. Mammalian lactoferrin receptors: Structure and function. Cell. Mol. Life Sci. 2005, 62, 2560–2575. [Google Scholar] [CrossRef]
- Pemberton, A.D.; Knight, P.A.; Wright, S.H.; Miller, R.P. Proteomic analysis of mouse jejunal epithelium and its response to infection with the intestinal nematode, Trichinella spiralis. Proteomics 2004, 4, 1101–1108. [Google Scholar] [CrossRef]
- González, J.; Geijo, M.V.; García-Pariente, C.; Verna, A.; Corpa, J.M.; Reyes, L.E.; Ferreras, M.C.; Juste, R.A.; García Marín, J.F.; Pérez, V. Histopathological classification of lesions associated with natural paratuberculosis infection in cattle. J. Comp. Path. 2005, 133, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Balseiro, A.; Pérez, V.; Juste, R.A. Chronic regional intestinal inflammatory disease: A trans-species slow infection? Comp. Immunol. Microbiol. Infect. Dis. 2019, 62, 88–100. [Google Scholar] [CrossRef]
- Vázquez, P.; Ruiz-Larrañaga, O.; Garrido, J.M.; Iriondo, M.; Manzano, C.; Agirre, M.; Estonba, A.; Juste, R.A. Genetic association analysis of paratuberculosis forms in Holstein-friesian cattle. Vet. Med. Int. 2014, 2014, 321327. [Google Scholar] [CrossRef]
- Park, H.E.; Park, H.T.; Jung, Y.H.; Yoo, H.S. Establishment a real-time reverse transcription PCR based on host biomarkers for the detection of the subclinical cases of Mycobacterium avium subsp. paratuberculosis. PLoS ONE 2017, 12, e0178336. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.P.; Tomlinson, G.; Marrie, T.J.; Tang, P.; McGeer, A.; Low, D.E.; Detsky, A.S.; Gold, W.L. Can routine laboratory tests discriminate between severe acute respiratory syndrome and other causes of community-acquired pneumonia? Clin. Infect. Dis. 2005, 40, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Bevins, C.L.; Salzman, N.H. Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nat. Rev. Microbiol. 2011, 9, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, S.; Yamashita, M.; Hoffman, D.R.; Nishiyama, A.; Shinohara, T.; Ohtsu, T.; Shibata, Y. Capture of heat-killed Mycobacterium bovis bacillus Calmette-Guérin by intelectin-1 deposited on cell surfaces. Glycobiology 2009, 19, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Voehringer, D.; Stanley, S.A.; Cox, J.S.; Completo, G.C.; Lowary, T.L.; Locksley, R.M. Nippostrongylus brasiliensis: Identification of intelectin-1 and -2 as Stat6-dependent genes expressed in lung and intestine during infection. Exp. Parasitol. 2007, 116, 458–466. [Google Scholar] [CrossRef]
- Chen, L.; Yan, J.; Sun, W.; Zhang, Y.; Sui, C.; Qi, J.; Du, Y.; Feng, L. A zebrafish intelectin ortholog agglutinates both Gram-negative and Gram-positive bacteria with binding capacity to bacterial polysaccharide. Fish Shellfish Immunol. 2016, 55, 729–736. [Google Scholar] [CrossRef]
- Pisanu, S.; Cubeddu, T.; Cacciotto, C.; Pilicchi, Y.; Pagnozzi, D.; Uzzau, S.; Rocca, S.; Addis, M.F. Characterization of paucibacillary ileal lesions in sheep with subclinical active infection by Mycobacterium avium subsp. paratuberculosis. Vet. Res. 2018, 49, 117. [Google Scholar] [CrossRef]
- Espinosa, J.; de la Morena, R.; Benavides, J.; García-Pariente, C.; Fernández, M.; Tesouro, M.; Arteche, N.; Vallejo, R.; Ferreras, M.C.; Pérez, V. Assessment of acute-phase protein response associated with the different pathological forms of bovine paratuberculosis. Animals 2020, 10, 1925. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Animal ID | Age (years) | Diagnostic Method | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Clinical Signs | Histological Classification | ZN | Infection Status | IDEXX ELISA | Real Time PCR | MAP Isolation | ||||
| Feces | Tissues | Feces | Tissues | |||||||
| 4 | 3.26 | ND | No lesions | - | Non infected | NEG | NEG | NEG | NEG | NEG |
| 13 | 0.81 | ND | No lesions | - | Non infected | NEG | NEG | NEG | NEG | NEG |
| 80 | 2.86 | NO | No lesions | - | Non infected | NEG | NEG | NEG | NEG | NEG |
| 94 | 2.70 | NO | No lesions | - | Non infected | NEG | NEG | NEG | NEG | NEG |
| 113 | 1.27 | NO | No lesions | - | Non infected | NEG | NEG | NEG | NEG | NEG |
| 8 | 4.71 | NO | Focal | + | Infected | NEG | NEG | POS | NEG | POS |
| 28 | 8.39 | NO | Focal | - | Infected | NEG | POS | POS | NEG | POS |
| 31 | 5.33 | NO | Focal | - | Infected | NEG | NEG | POS | NEG | NEG |
| 44 | 5.45 | NO | Focal | - | Infected | NEG | NEG | NEG | NEG | POS |
| 52 | 6.59 | NO | Focal | - | Infected | POS | POS | POS | NEG | POS |
| 11 | 7.31 | NO | Multifocal | + | Infected | NEG | NEG | NEG | NEG | POS |
| 46 | 2.75 | YES | Multifocal | + | Infected | NEG | NEG | NEG | NEG | NEG |
| 64 | 3.62 | NO | Multifocal | + | Infected | POS | POS | POS | POS | POS |
| 97 | 2.96 | YES | Multifocal | ++ | Infected | POS | POS | POS | NEG | POS |
| 110 | 4.03 | NO | Multifocal | - | Infected | NEG | NEG | POS | NEG | POS |
| 26 | 5.46 | NO | D. intermediate | + | Infected | POS | POS | POS | NEG | POS |
| 30 | 4.14 | YES | D.intermediate | + | Infected | NEG | NEG | NEG | NEG | NEG |
| 68 | 6.01 | YES | D. intermediate | ++ | Infected | POS | POS | POS | POS | POS |
| 88 | 2.92 | YES | D. histiocytic | +++ | Infected | POS | POS | POS | NEG | POS |
| 103 | 10.39 | YES | D. histiocytic | + | Infected | POS | POS | POS | NEG | POS |
| Animal ID | Histopathological Classification | Infection Status | Mean Number | Mean Value |
|---|---|---|---|---|
| per Cow * | per Type ** | |||
| 4 | No lesions | Non-infected | 0.1 ± 0.0 | |
| 13 | No lesions | Non-infected | 184.5 ± 82.9 | |
| 80 | No lesions | Non-infected | 2.2 ± 4.7 | 41.0 ± 7.1 |
| 94 | No lesions | Non-infected | 16.7 ± 17.9 | |
| 113 | No lesions | Non-infected | 1.7 ± 4.4 | |
| 8 | Focal | Infected | 80.7 ± 89.6 | |
| 28 | Focal | Infected | 174.9 ± 116.4 | |
| 31 | Focal | Infected | 146.6 ± 71.5 | 116.9 ± 16.1 |
| 44 | Focal | Infected | 30.3 ± 67.8 | |
| 52 | Focal | Infected | 152.1 ± 153.4 | |
| 11 | Multifocal | Infected | 253.3 ± 210.8 | |
| 46 | Multifocal | Infected | 28.5 ± 38.9 | |
| 64 | Multifocal | Infected | 154.3 ± 122.3 | 108.7 ± 19.9 |
| 97 | Multifocal | Infected | 37.8 ± 45.5 | |
| 110 | Multifocal | Infected | 69.4 ± 67.1 | |
| 26 | Diffuse intermediate | Infected | 34.6 ± 40.3 | |
| 30 | Diffuse intermediate | Infected | 15.8 ± 26.7 | |
| 68 | Diffuse intermediate | Infected | 46.0 ± 66.0 | 76.50 ± 13.8 |
| 88 | Diffuse histiocytic | Infected | 90.6 ± 107.5 | |
| 103 | Diffuse histiocytic | Infected | 195.7 ± 103.5 |
| ROC Analysis | AUC | p Value | CUT OFF | SE (%) | SP (%) | DV |
|---|---|---|---|---|---|---|
| Focal vs. Control | 0.800 | 0.134 | >167 | 100.00 | 80.00 | 0.90 |
| Multifocal vs. Control | 0.840 | 0.039 | >167 | 100.00 | 80.00 | 0.90 |
| Diffuse vs. Control | 0.800 | 0.073 | >22 | 100.00 | 60.00 | 0.80 |
| Patent vs. Control | 0.820 | 0.046 | >167 | 90.00 | 80.00 | 0.85 |
| Lesions vs. Control | 0.813 | 0.069 | >167 | 93.33 | 80.00 | 0.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanco Vázquez, C.; Balseiro, A.; Alonso-Hearn, M.; Juste, R.A.; Iglesias, N.; Canive, M.; Casais, R. Bovine Intelectin 2 Expression as a Biomarker of Paratuberculosis Disease Progression. Animals 2021, 11, 1370. https://doi.org/10.3390/ani11051370
Blanco Vázquez C, Balseiro A, Alonso-Hearn M, Juste RA, Iglesias N, Canive M, Casais R. Bovine Intelectin 2 Expression as a Biomarker of Paratuberculosis Disease Progression. Animals. 2021; 11(5):1370. https://doi.org/10.3390/ani11051370
Chicago/Turabian StyleBlanco Vázquez, Cristina, Ana Balseiro, Marta Alonso-Hearn, Ramón A. Juste, Natalia Iglesias, Maria Canive, and Rosa Casais. 2021. "Bovine Intelectin 2 Expression as a Biomarker of Paratuberculosis Disease Progression" Animals 11, no. 5: 1370. https://doi.org/10.3390/ani11051370
APA StyleBlanco Vázquez, C., Balseiro, A., Alonso-Hearn, M., Juste, R. A., Iglesias, N., Canive, M., & Casais, R. (2021). Bovine Intelectin 2 Expression as a Biomarker of Paratuberculosis Disease Progression. Animals, 11(5), 1370. https://doi.org/10.3390/ani11051370