
Ontogeny of OPN4, OPN5, GnRH and GnIH mRNA Expression in the Posthatch Male and Female Pekin Duck (Anas platyrhynchos domesticus) Suggests OPN4 May Have Additional Functions beyond Reproduction

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Tissue and RNA Collection
2.3. qRT-PCR
2.4. Hormone Analyses
2.5. Statistical Analyses
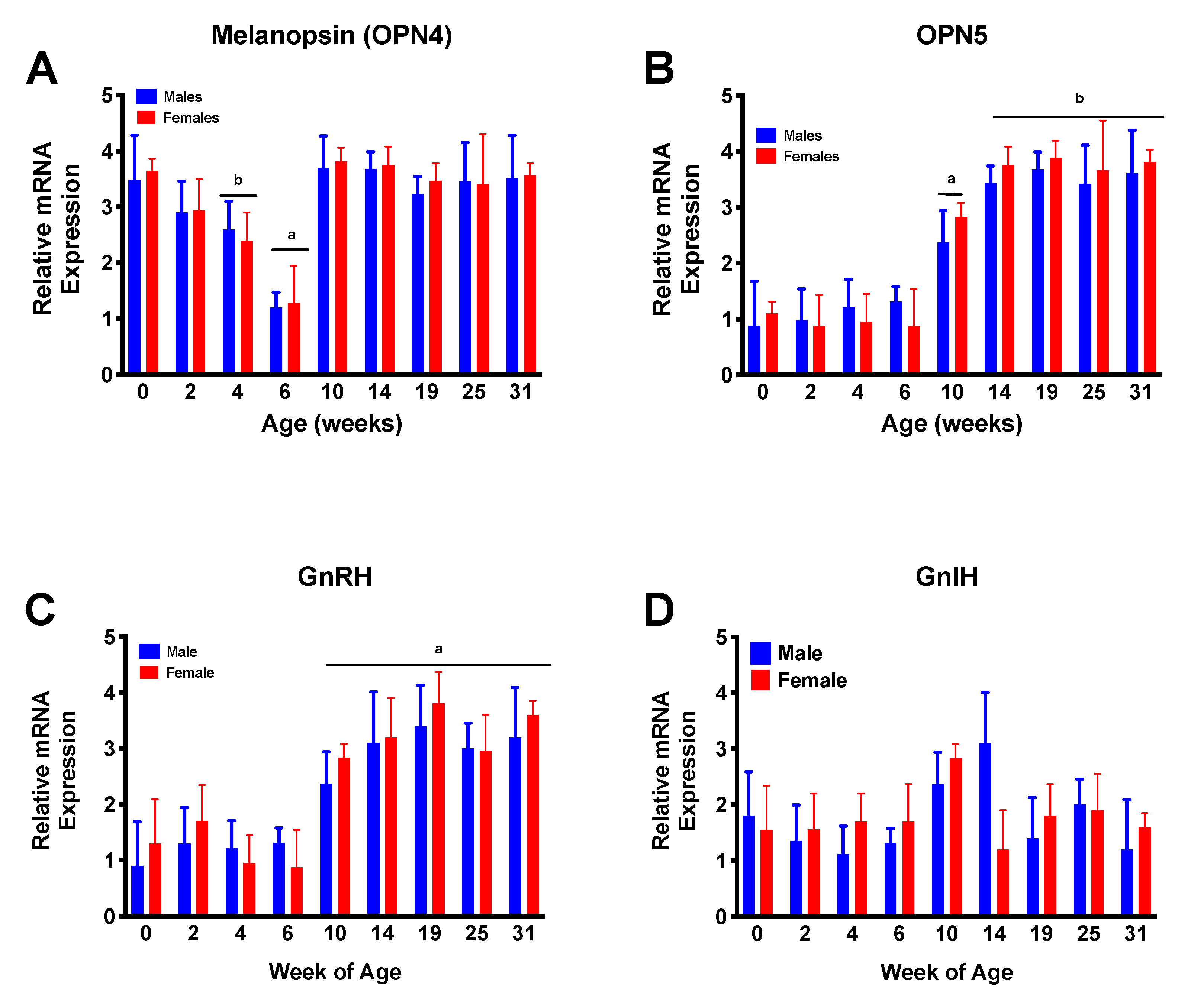
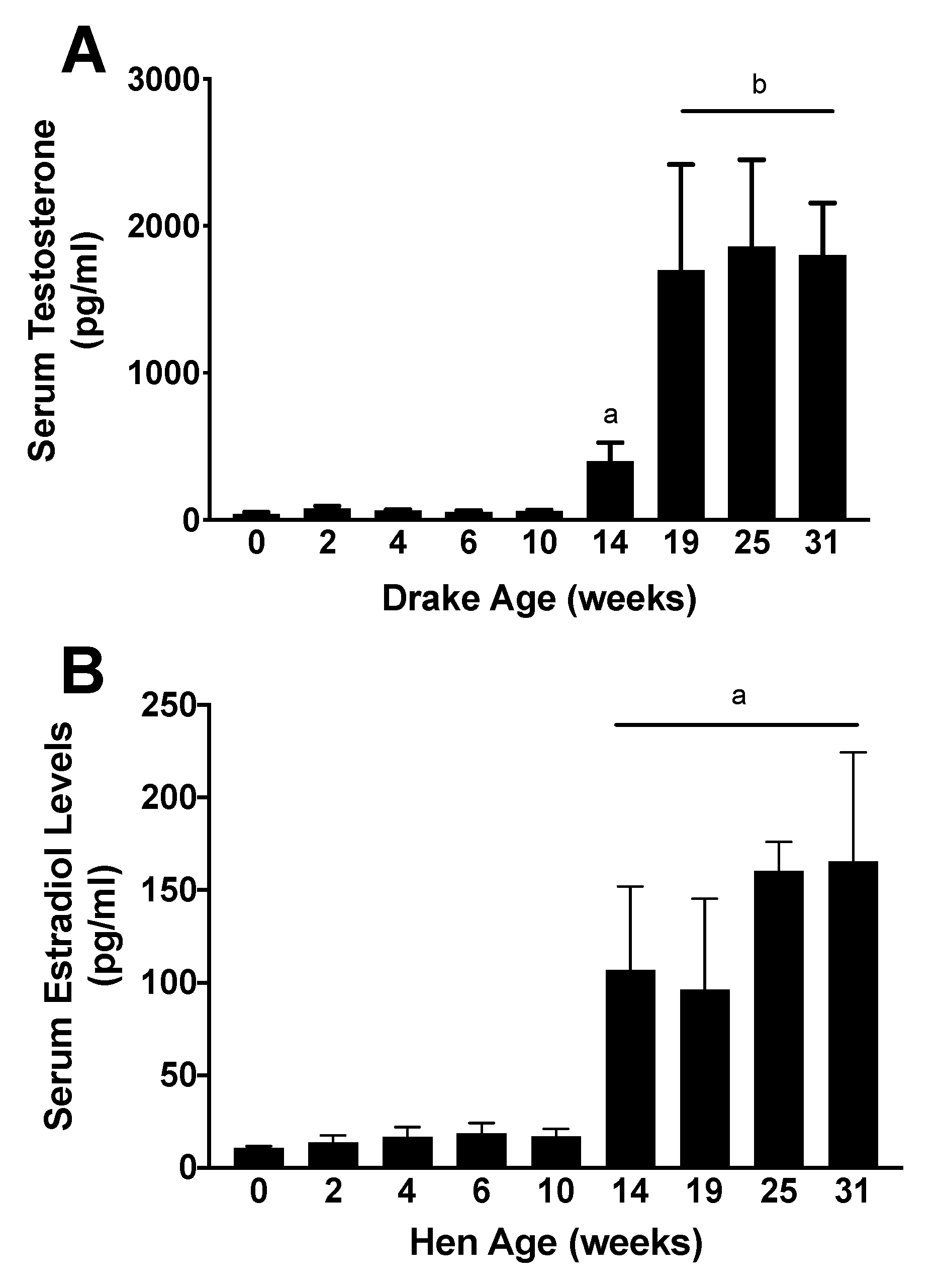
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benoit, J.; Assenmacher, I.; Walter, F.X. Gonadotropic activity of the hypophysis of the domestic duck during seasonal testicular regression and prepuberty. C. R. Seances. Soc. Biol. Fil. 1950, 144, 1403–1407. [Google Scholar] [PubMed]
- Benoit, J.; Assenmacher, I.; Brard, E. Action of prolonged permanent lighting on the testicular development of the Pekin duck. Arch. Anat. Microsc. Morphol. Exp. 1959, 48, 5–11. [Google Scholar] [PubMed]
- Benoit, J.; Assenmacher, I. Hypothalamo-hypophyseal relationship and its role in gonadostimulation in domestic duck. J. Physiol. 1951, 43, 643–645. [Google Scholar]
- Wells, M.C.; Lehner, P.N.; Bolen, E.G.; Rylander, M.K. Comparison of scotopic sensitivity in diurnal (Anas platyrhynchos) and crepuscular (Dendrocygna autumnalis) ducks. J. Comp. Physiol. Psychol. 1975, 88, 940–944. [Google Scholar] [CrossRef]
- Campbell, C.L.; Colton, S.; Haas, R.; Rice, M.; Porter, A.; Schenk, A.; Meelker, A.; Fraley, S.M.; Fraley, G.S. Effects of different wavelengths of light on the biology, behavior, and production of grow-out Pekin ducks. Poult. Sci. 2015, 94, 1751–1757. [Google Scholar] [CrossRef]
- House, G.; Sobotik, E.; Nelson, J.; Archer, G. The effect of raising Pekin ducks two spectra of LED light on production, stress, and behavior. In Proceedings of the Poultry Science Association Annual Meeting, San Antonio, TX, USA, 23–26 July 2018; p. 347. [Google Scholar]
- Porter, L.; Porter, A.; Potter, H.; Alenciks, E.; Fraley, S.M.; Fraley, G.S. Low light intensity in Pekin duck breeder barns has a greater impact on the fertility of drakes than hens. Poult. Sci. 2018, 97, 4262–4271. [Google Scholar] [CrossRef]
- Benoit, J. The Role of the Eye and of the Hypothalamus in the Photostimulation of Gonads in the Duck. Ann. N. Y. Acad. Sci. 1964, 117, 204–216. [Google Scholar] [CrossRef]
- Cherry, P.; Morris, T.R. Domestic Duck Production: Science and Practice; CABI: Wallingford, UK; Oxfordshire, UK; Cambridge, MA, USA, 2008; ISBN 9780851990545. [Google Scholar]
- Haas, R.; Alenciks, E.; Frazier, K.; Fraley, G.S. The Maintenance of Reproductive Status in Pekin Drakes Requires Both Red and Blue Wavelengths of Light: Relationship to Opsin-Related Proteins in the Hypothalamus. Poult. Sci. 2017, 96, 2908–2919. [Google Scholar] [CrossRef]
- Colton, S.; Fraley, G.S. The effects of environmental enrichment devices on feather picking in commercially housed Pekin ducks. Poult. Sci. 2014, 93, 2143–2150. [Google Scholar] [CrossRef]
- Haas, R.; Alenciks, E.; Meddle, S.; Fraley, G.S.S.S. Expression of deep brain photoreceptors in the Pekin drake: A possible role in the maintenance of testicular function. Poult. Sci. 2017, 138, 118–123. [Google Scholar] [CrossRef]
- Guh, Y.-J.; Tamai, T.K.; Yoshimura, T. The underlying mechanisms of vertebrate seasonal reproduction. Proc. Jpn. Acad. Ser. B 2019, 95, 343–357. [Google Scholar] [CrossRef]
- Ubuka, T.; Bentley, G.E.; Tsutsui, K. Neuroendocrine regulation of gonadotropin secretion in seasonally breeding birds. Front. Neurosci. 2013, 7, 38. [Google Scholar] [CrossRef]
- Johnson, P.A.; Van Tienhoven, A. Investigations of the significance of the crepuscular LH peak in the ovulatory cycle of the hen (Gallus domesticus). J. Endocrinol. 1984. [Google Scholar] [CrossRef]
- Johnson, A.L.; Van Tienhoven, A. Plasma Concentrations of Six Steroids and LH During the Ovulatory Cycle of the Hen, Gallus domesticus1. Biol. Reprod. 1980. [Google Scholar] [CrossRef] [PubMed]
- Porter, L.; Alenciks, E.; Fraley, G.S. Gonadal regression elicited in Pekin duck drakes and hens associated with supplemental light from kerosene lanterns during the winter months. In Proceedings of the Poultry Science Association Annual Meeting, San Antonio, TX, USA, 23–26 July 2016; p. 195. [Google Scholar]
- Van Tienhoven, A. No Title. In Reproductive Physiology of Vertebrates; Cornell University Press: New York, NY, USA, 1983; pp. 80–94. [Google Scholar]
- Yoshimura, T. Neuroendocrine mechanism of seasonal reproduction in birds and mammals. Anim. Sci. J. 2010, 81, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.W.; Kuenzel, W.J. Deep-brain photoreceptors (DBPs) involved in the photoperiodic gonadal response in an avian species, Gallus gallus. Gen. Comp. Endocrinol. 2015, 211, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Nakane, Y.; Yoshimura, T. Deep brain photoreceptors and a seasonal signal transduction cascade in birds. Cell Tissue Res. 2010, 342, 341–344. [Google Scholar] [CrossRef] [PubMed]
- Kuenzel, W.J. The search for deep encephalic photoreceptors within the avian brain, using gonadal development as a primary indicator. Poult. Sci. 1993, 72, 959–967. [Google Scholar] [CrossRef] [PubMed]
- House, G.M.; Sobotik, E.B.; Nelson, J.R.; Archer, G.S. A comparison of white/red and white/blue LED light fixtures on Pekin duck production, stress and behaviour. Br. Poult. Sci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.X.X.; Hu, M.D.; Guo, B.B.B.; Qu, X.L.L.; Lei, M.M.M.; Chen, R.; Chen, Z.; Shi, Z.D.D. Effect and molecular regulatory mechanism of monochromatic light colors on the egg-laying performance of Yangzhou geese. Anim. Reprod. Sci. 2019, 204, 131–139. [Google Scholar] [CrossRef]
- Nakane, Y.; Ikegami, K.; Ono, H.; Yamamoto, N.; Yoshida, S.; Hirunagi, K.; Ebihara, S.; Kubo, Y.; Yoshimura, T. A mammalian neural tissue opsin (Opsin 5) is a deep brain photoreceptor in birds. Proc. Natl. Acad. Sci. USA 2010, 107, 15264–15268. [Google Scholar] [CrossRef] [PubMed]
- Bailes, H.J.; Lucas, R.J. Human melanopsin forms a pigment maximally sensitive to blue light (λmax ≈ 479 nm) supporting activation of Gq/11 and Gi/o signalling cascades. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122987. [Google Scholar] [CrossRef]
- Potter, H.; Alenciks, E.; Frazier, K.; Porter, A.; Fraley, G.S.S. Immunolesion of melanopsin neurons causes gonadal regression in Pekin drakes (Anas platyrhynchos domesticus). Gen. Comp. Endocrinol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Glass, J.D.; Lauber, J.K. Sites and action spectra for encephalic photoreception in the Japanese quail. Am. J. Physiol 1981, 240, R220–R228. [Google Scholar] [CrossRef]
- Menaker, M. Extraretinal light perception in the sparrow, I. entrainment of the biological clock. Proc. Natl. Acad. Sci. USA 1968, 59, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Underwood, H.; Menaker, M. Photoreception in sparrows: Response to photoperiodic stimuli. Science 1970, 169, 893. [Google Scholar]
- Benoit, J.M. Chronobiologic study in the domestic duck. II. Physiological mechanism of the chronobiologic action of visible light on the gonads of the male duck. Chronobiologia 1978, 5, 158–168. [Google Scholar]
- Wang, G.; Ramenofsky, M.; Wingfield, J.C. Apparent dissociation of photoperiodic time measurement between vernal migration and breeding under dim green light conditions in Gambel’s white-crowned sparrow Zonotrichia leucophrys gambelii. Curr. Zool. 2013, 59, 349–359. [Google Scholar] [CrossRef]
- Oliver, J.; Bayle, J.D. Brain photoreceptors for the photo-induced testicular response in birds. Experientia 1982, 38, 1021–1029. [Google Scholar] [CrossRef]
- Menaker, M.; Roberts, R.; Elliott, J.; Underwood, H. Extraretinal light perception in the sparrow. 3. The eyes do not participate in photoperiodic photoreception. Proc. Natl. Acad. Sci. USA 1970, 67, 320–325. [Google Scholar] [CrossRef]
- Underwood, H.; Menaker, M. Photoperiodically significant photoreception in sparrows: Is the retina involved? Science 1970, 167, 298–301. [Google Scholar] [CrossRef] [PubMed]
- Underwood, H.; Menaker, M. Extraretinal light perception: Entrainment of the biological clock controlling lizard locomotor activity. Science 1970, 170, 190–193. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Kuenzel, W.J. A possible neural cascade involving the photoneuroendocrine system (PNES) responsible for regulating gonadal development in an avian species, Gallus gallus. Brain Res. Bull. 2008, 76, 586–596. [Google Scholar] [CrossRef] [PubMed]
- Halford, S.; Pires, S.S.; Turton, M.; Zheng, L.; Gonzalez-Menendez, I.; Davies, W.L.; Peirson, S.N.; Garcia-Fernandez, J.M.; Hankins, M.W.; Foster, R.G. VA opsin-based photoreceptors in the hypothalamus of birds. Curr. Biol. 2009, 19, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Saldanha, C.J.; Silverman, A.J.; Silver, R. Direct innervation of GnRH neurons by encephalic photoreceptors in birds. J. Biol. Rhythm. 2001, 16, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Vigh, B.; Vigh-Teichmann, I. Actual problems of the cerebrospinal fluid-contacting neurons. Microsc. Res. Tech. 1998, 41, 57–83. [Google Scholar] [CrossRef]
- Kuenzel, W.J.; Kang, S.W.; Zhou, Z.J. Exploring avian deep-brain photoreceptors and their role in activating the neuroendocrine regulation of gonadal development. Poult. Sci. 2015, 94, 786–798. [Google Scholar] [CrossRef]
- Scott, S.M.; Alm, J.; Fisher, D.A. Effect of thyroid hormone and sex status on epidermal growth factor concentrations in the submandibular gland of a congenitally hypothyroid mouse model. J. Dev. Physiol. 1986, 8, 461–466. [Google Scholar]
- Wilson, F.E. A test of the hypothesis that T3 is the “seasonality” thyroid hormone in American tree sparrows (Spizella arborea): Intracerebroventricular infusion of iopanoic acid, an inhibitor of T3 synthesis and degradation. J. Comp. Physiol. B 2001, 171, 113–119. [Google Scholar] [CrossRef]
- Scanes, C.G.; Jallageas, M.; Assenmacher, I. Seasonal variations in the circulating concentrations of growth hormone in male Peking duck (Anas platyrhynchos) and teal (Anas crecca); correlations with thyroidal function. Gen. Comp. Endocrinol. 1980, 41, 76–79. [Google Scholar] [CrossRef]
- Wilson, F.E.; Reinert, B.D. The timing of thyroid-dependent programming in seasonally breeding male American tree sparrows (Spizella arborea). Gen. Comp. Endocrinol. 1996, 103, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Kosonsiriluk, S.; Mauro, L.J.; Chaiworakul, V.; Chaiseha, Y.; El Halawani, M.E. Photoreceptive oscillators within neurons of the premammillary nucleus (PMM) and seasonal reproduction in temperate zone birds. Gen. Comp. Endocrinol. 2013, 190. [Google Scholar] [CrossRef] [PubMed]
- Pérez, J.H.; Furlow, J.D.; Wingfield, J.C.; Ramenofsky, M. Regulation of vernal migration in Gambel’s white-crowned sparrows: Role of thyroxine and triiodothyronine. Horm. Behav. 2016, 84, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Reinert, B.D.; Wilson, F.E. The thyroid and the hypothalamus-pituitary-ovarian axis in American tree sparrows (Spizella arborea). Gen. Comp. Endocrinol 1996, 103, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Mishra, I.; Bhardwaj, S.K.; Malik, S.; Kumar, V. Concurrent hypothalamic gene expression under acute and chronic long days: Implications for initiation and maintenance of photoperiodic response in migratory songbirds. Mol. Cell. Endocrinol. 2017, 439, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Hayase, S.; Aoki, N.; Takehara, A.; Ishigohoka, J.; Matsushima, T.; Wada, K.; Homma, K.J. Sex differences in brain thyroid hormone levels during early post-hatching development in zebra finch (Taeniopygia guttata). PLoS ONE 2017, 12, e0169643. [Google Scholar] [CrossRef]
- Schew, W.A.; McNabb, F.M.; Scanes, C.G. Comparison of the ontogenesis of thyroid hormones, growth hormone, and insulin-like growth factor-I in ad libitum and food-restricted (altricial) European starlings and (precocial) Japanese quail. Gen. Comp. Endocrinol. 1996, 101, 304–316. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Aoki, N.; Kitajima, T.; Iikubo, E.; Katagiri, S.; Matsushima, T.; Homma, K.J. Thyroid hormone determines the start of the sensitive period of imprinting and primes later learning. Nat. Commun. 2012. [Google Scholar] [CrossRef]
- Sirsat, T.S. The Role of Thyroid Hormone Across Avian Development Spectrum: Investigations on Systemic Development, Metabolism and Ontogeny of Endothermy; University of North Texas: Denton, YX, USA, 2017. [Google Scholar]
- Sirsat, T.S.; Crossley, D.A.; Crossley, J.L.; Dzialowski, E.M. Thyroid hormone manipulation influences development of cardiovascular regulation in embryonic Pekin duck, Anas platyrhynchos domestica. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2018. [Google Scholar] [CrossRef]
- Spiers, D.E.; McNabb, R.A.; McNabb, F.M.A. The development of thermoregulatory ability, heat seeking activities, and thyroid function in hatchling Japanese quail (Coturnix coturnix japonica). J. Comp. Physiol. 1974. [Google Scholar] [CrossRef]
- Olson, J.M.; McNabb, F.M.A.; Jablonski, M.S.; Ferris, D.V. Thyroid development in relation to the development of endothermy in the red-winged blackbird (Agelaius phoeniceus). Gen. Comp. Endocrinol. 1999. [Google Scholar] [CrossRef]
- Anne McNabb, F.M.; Olson, J.M. Development of thermoregulation and its hormonal control in precocial and altricial birds. Avian Poult. Biol. Rev. 1996, 7, 111–125. [Google Scholar]
- McNaughton, F.J.; Dawson, A.; Goldsmith, A.R. Puberty in birds: The reproductive system of starlings does not respond to short days until birds are fully grown. J. Endocrinol. 1992. [Google Scholar] [CrossRef] [PubMed]
- Dawson, A.; King, V.M.; Bentley, G.E.; Ball, G.F. Photoperiodic control of seasonality in birds. J. Biol. Rhythms 2001, 16. [Google Scholar] [CrossRef] [PubMed]
- MacDougall-Shackleton, S.A.; Stevenson, T.J.; Watts, H.E.; Pereyra, M.E.; Hahn, T.P. The evolution of photoperiod response systems and seasonal GnRH plasticity in birds. Integr. Comp. Biol. 2009, 49, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Bentley, G.E.; Ubuka, T.; McGuire, N.L.; Calisi, R.; Perfito, N.; Kriegsfeld, L.J.; Wingfield, J.C.; Tsutsui, K. Gonadotrophin-inhibitory hormone: A multifunctional neuropeptide. J. Neuroendocr. 2009, 21, 276–281. [Google Scholar] [CrossRef]
- Fraley, G.S.S.; Coombs, E.; Gerometta, E.; Colton, S.; Sharp, P.J.J.; Li, Q.; Clarke, I.J.J. Distribution and sequence of gonadotropin-inhibitory hormone and its potential role as a molecular link between feeding and reproductive systems in the Pekin duck (Anas platyrhynchos domestica). Gen. Comp. Endocrinol. 2013, 184, 103–110. [Google Scholar] [CrossRef]
- Shimizu, M.; Bedecarrats, G.Y. Activation of the chicken gonadotropin-inhibitory hormone receptor reduces gonadotropin releasing hormone receptor signaling. Gen. Comp. Endocrinol. 2010, 167, 331–337. [Google Scholar] [CrossRef]
- Tsutsui, K.; Saigoh, E.; Ukena, K.; Teranishi, H.; Fujisawa, Y.; Kikuchi, M.; Ishii, S.; Sharp, P.J. A novel avian hypothalamic peptide inhibiting gonadotropin release. Biochem. Biophys. Res. Commun. 2000, 275, 661–667. [Google Scholar] [CrossRef]
- Ubuka, T.; McGuire, N.L.; Calisi, R.M.; Perfito, N.; Bentley, G.E. The control of reproductive physiology and behavior by gonadotropin-inhibitory hormone. Integr. Comp. Biol. 2008, 48, 560–569. [Google Scholar] [CrossRef]
- McConn, B.; Wang, G.; Yi, J.; Gilbert, E.R.; Osugi, T.; Ubuka, T.; Tsutsui, K.; Chowdhury, V.S.; Furuse, M.; Cline, M.A. Gonadotropin-inhibitory hormone-stimulation of food intake is mediated by hypothalamic effects in chicks. Neuropeptides 2014, 48, 327–334. [Google Scholar] [CrossRef]
- Tobari, Y.; Son, Y.L.; Ubuka, T.; Hasegawa, Y.; Tsutsui, K. A New Pathway Mediating Social Effects on the Endocrine System: Female Presence Acting via Norepinephrine Release Stimulates Gonadotropin-Inhibitory Hormone in the Paraventricular Nucleus and Suppresses Luteinizing Hormone in Quail. J. Neurosci. 2014, 34, 9803–9811. [Google Scholar] [CrossRef]
- Ramesh, R.; Kuenzel, W.J.; Buntin, J.D.; Proudman, J.A. Identification of growth-hormone- and prolactin-containing neurons within the avian brain. Cell Tissue Res. 2000, 299, 371–383. [Google Scholar] [CrossRef]
- Ernst, D.K.; Lynn, S.E.; Bentley, G.E. Differential response of GnIH in the brain and gonads following acute stress in a songbird. Gen. Comp. Endocrinol. 2016, 227, 51–57. [Google Scholar] [CrossRef]
- Calisi, R.M.; Rizzo, N.O.; Bentley, G.E. Seasonal differences in hypothalamic EGR-1 and GnIH expression following capture-handling stress in house sparrows (Passer domesticus). Gen. Comp. Endocrinol. 2008, 157, 283–287. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Target | Gene | Forward | Reverse |
|---|---|---|---|
| DBP | OPN4 | CTCGCCATAGAACATCCGCA | ACTGAACAGGCTACTCCCCTT |
| OPN5 | TTT CTC ACC GCT GGA TCT TT | CAG GCA GAT AAA GGC ATG GTG T | |
| Reproductive | GnRH-1 | ATC GCA AAC GAA ATG GAA AG | CTG GCT TCT CCT TCG ATC AG |
| GnIH | TAA CAC CGC ATG GTA TGT GC | CTC CTC TGC TCT TCC TCC AA | |
| Housekeeping | GAPDH | GGTTGTCTCCTGCGACTTCA | TCCTTGGATGCCATGTGGAC |
| β-actin | CAC AAT GTA CCC GGG CAT CG | ACA TCT GCT GGA AGG TGG AC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Wyk, B.; Fraley, G. Ontogeny of OPN4, OPN5, GnRH and GnIH mRNA Expression in the Posthatch Male and Female Pekin Duck (Anas platyrhynchos domesticus) Suggests OPN4 May Have Additional Functions beyond Reproduction. Animals 2021, 11, 1121. https://doi.org/10.3390/ani11041121
Van Wyk B, Fraley G. Ontogeny of OPN4, OPN5, GnRH and GnIH mRNA Expression in the Posthatch Male and Female Pekin Duck (Anas platyrhynchos domesticus) Suggests OPN4 May Have Additional Functions beyond Reproduction. Animals. 2021; 11(4):1121. https://doi.org/10.3390/ani11041121
Chicago/Turabian StyleVan Wyk, Brooke, and Gregory Fraley. 2021. "Ontogeny of OPN4, OPN5, GnRH and GnIH mRNA Expression in the Posthatch Male and Female Pekin Duck (Anas platyrhynchos domesticus) Suggests OPN4 May Have Additional Functions beyond Reproduction" Animals 11, no. 4: 1121. https://doi.org/10.3390/ani11041121
APA StyleVan Wyk, B., & Fraley, G. (2021). Ontogeny of OPN4, OPN5, GnRH and GnIH mRNA Expression in the Posthatch Male and Female Pekin Duck (Anas platyrhynchos domesticus) Suggests OPN4 May Have Additional Functions beyond Reproduction. Animals, 11(4), 1121. https://doi.org/10.3390/ani11041121