
PDLIM5 Affects Chicken Skeletal Muscle Satellite Cell Proliferation and Differentiation via the p38-MAPK Pathway

 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Animals and Cell Culture
2.3. Vector Construction, RNA Oligonucleotides, and Cell Transfection
2.4. RNA Extraction and Expression Detection
2.5. CCK-8 and EdU (5-ethynyl-2′-deoxyuridine) Assay
2.6. Immunofluorescence and Western Blot
2.7. Transcriptome Analysis
2.8. Statistical Analysis
3. Results
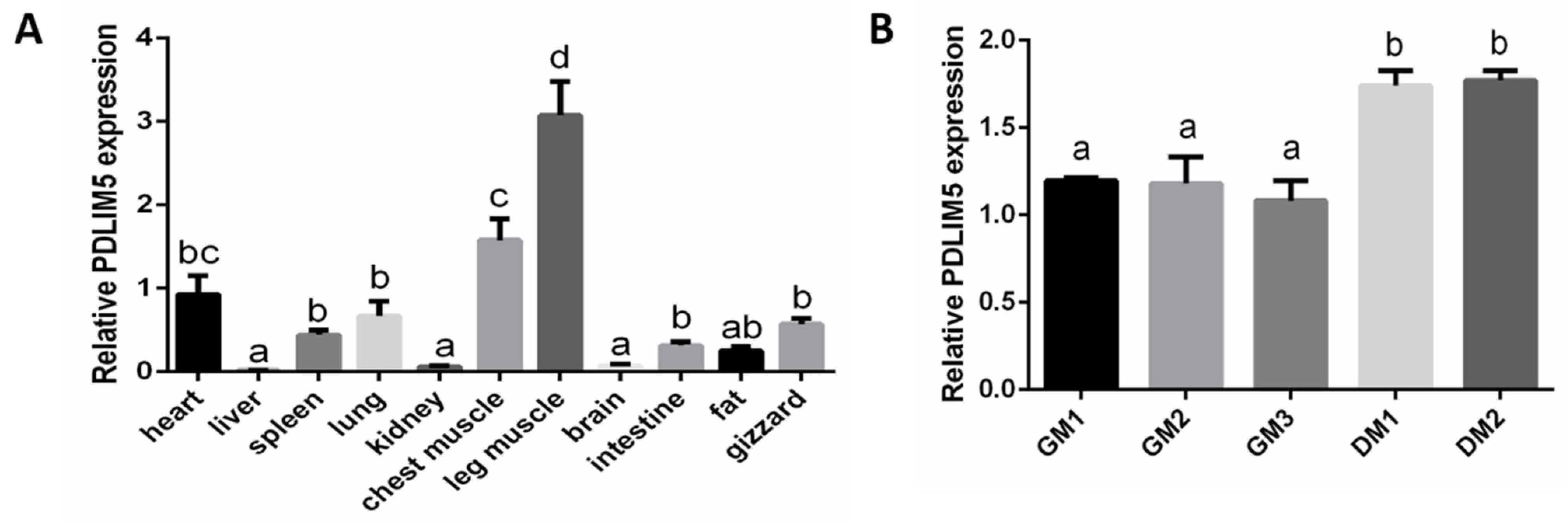
3.1. PDLIM5 Tissue Distribution in Vhicken
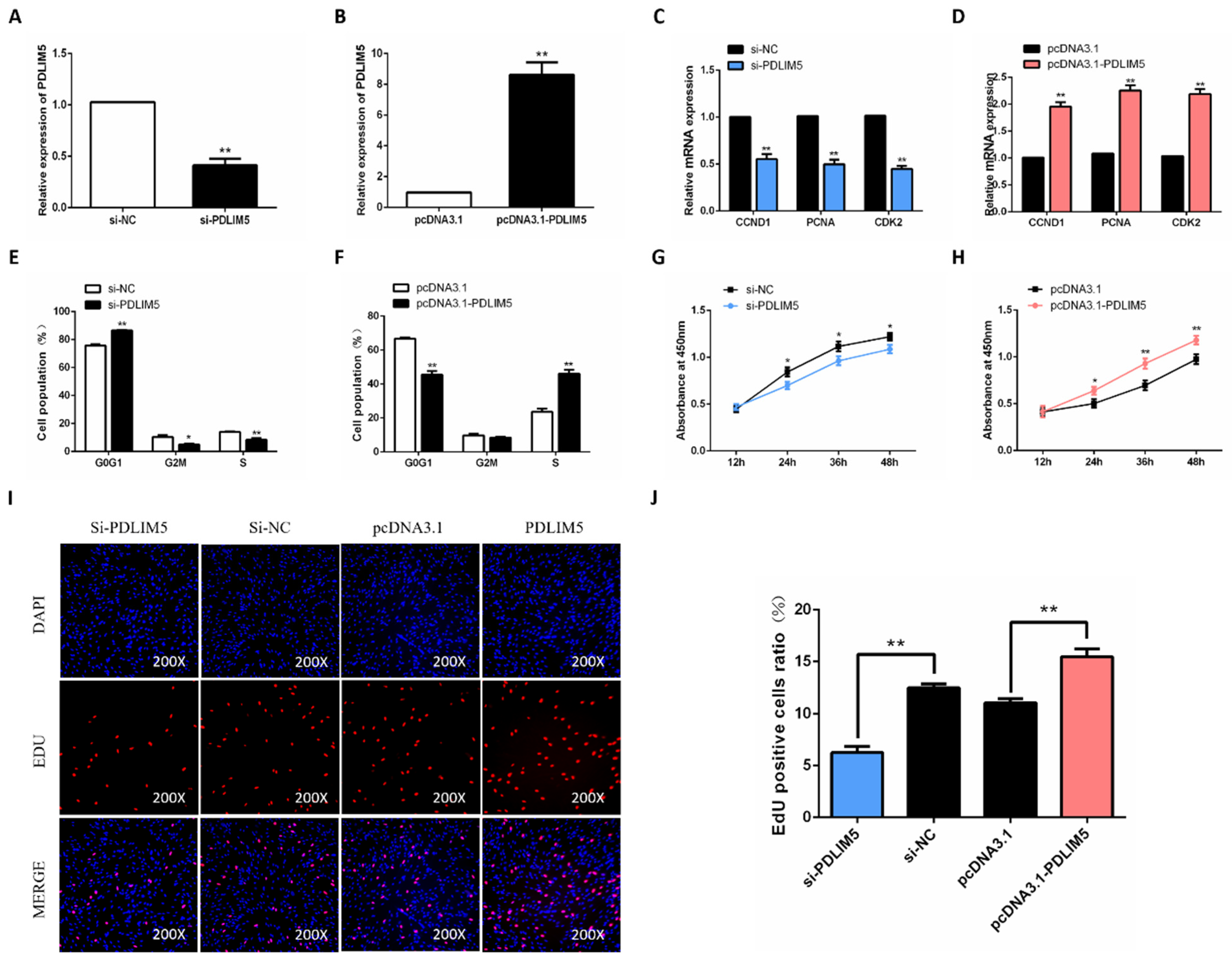
3.2. PDLIM5 Increases Proliferation of Chicken SMSCs
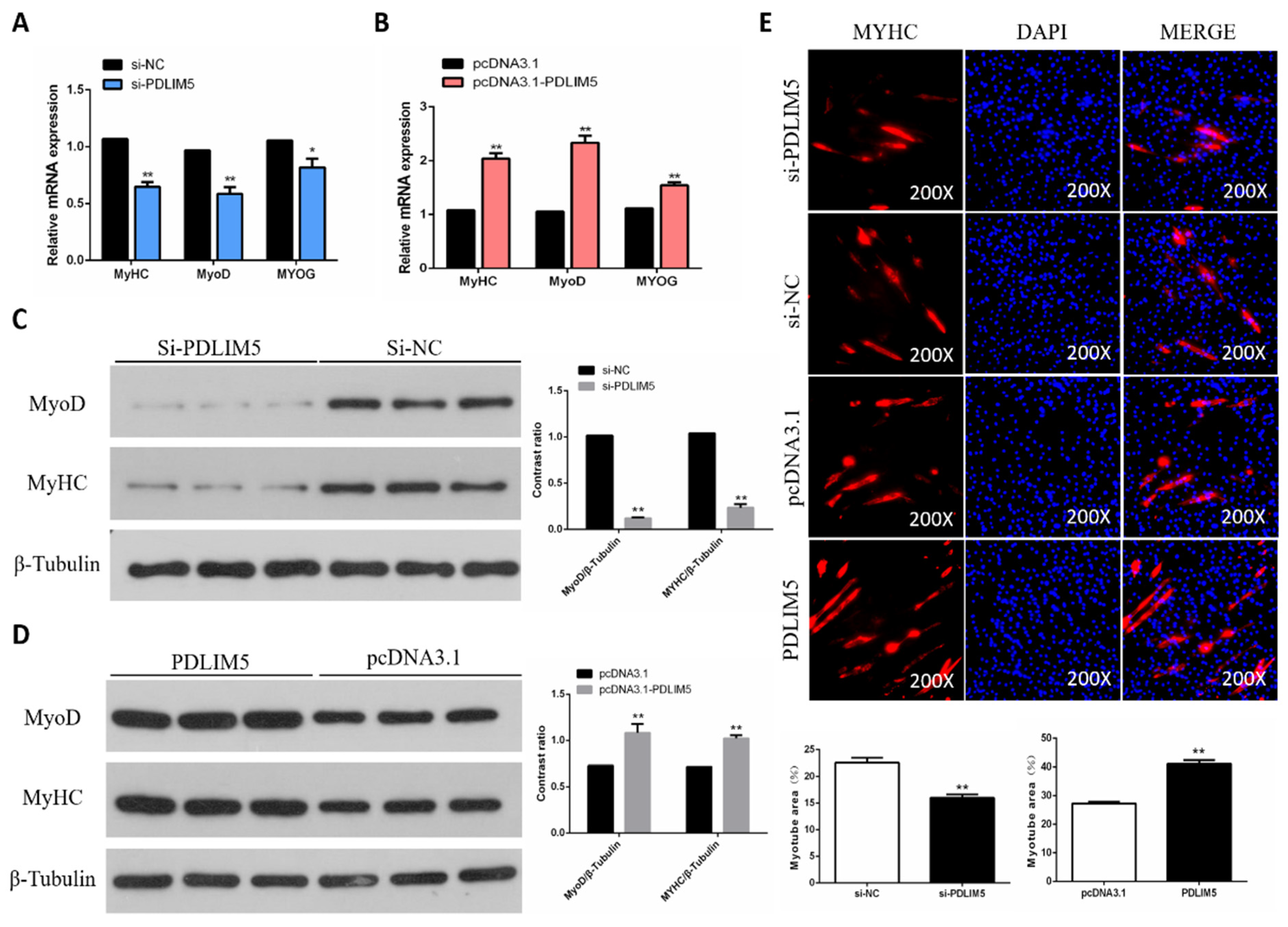
3.3. PDLIM5 Increased Differentiation of Chicken SMSCs
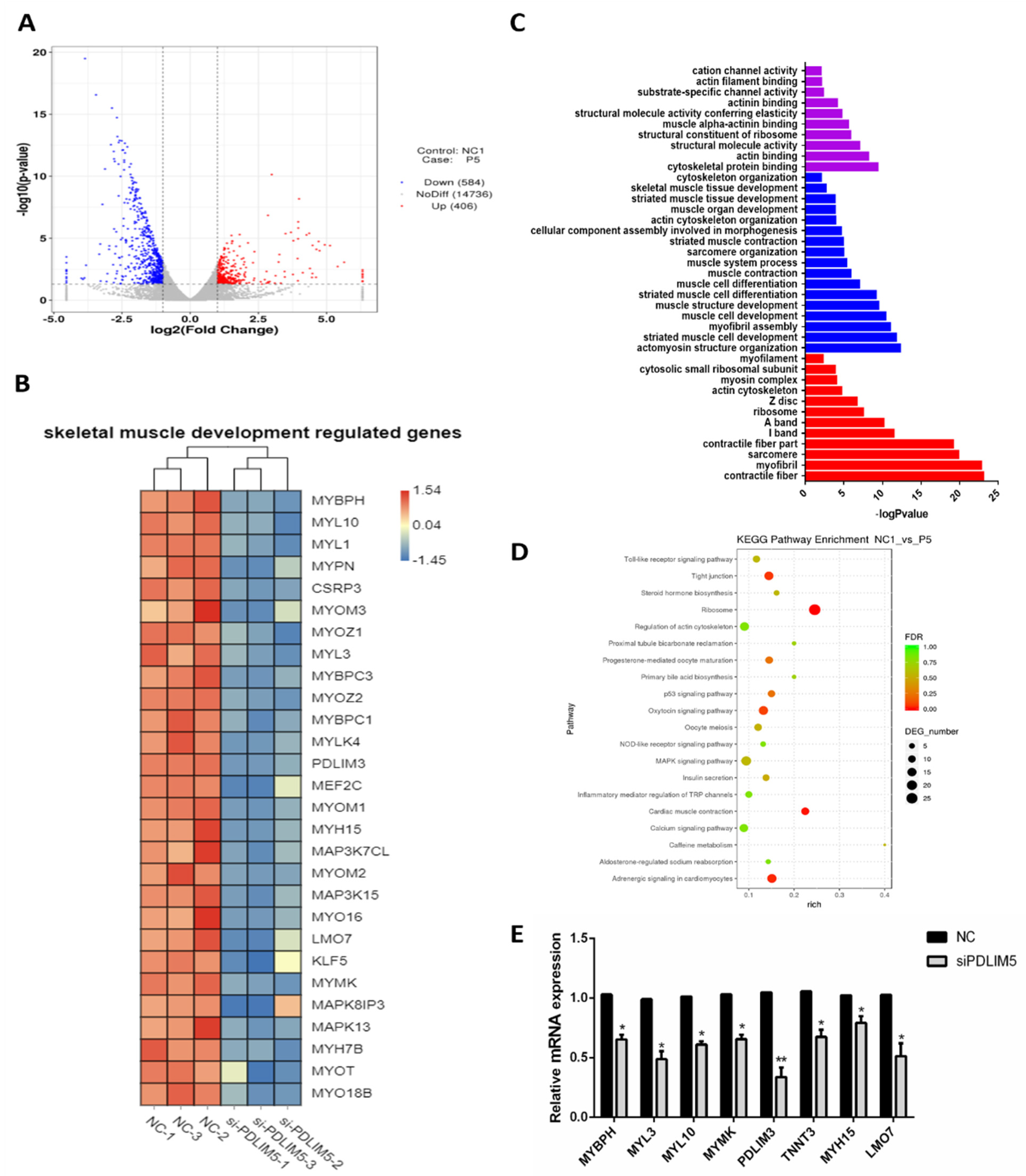
3.4. Gene Expression Analysis of PDLIM5 Silenced Cells
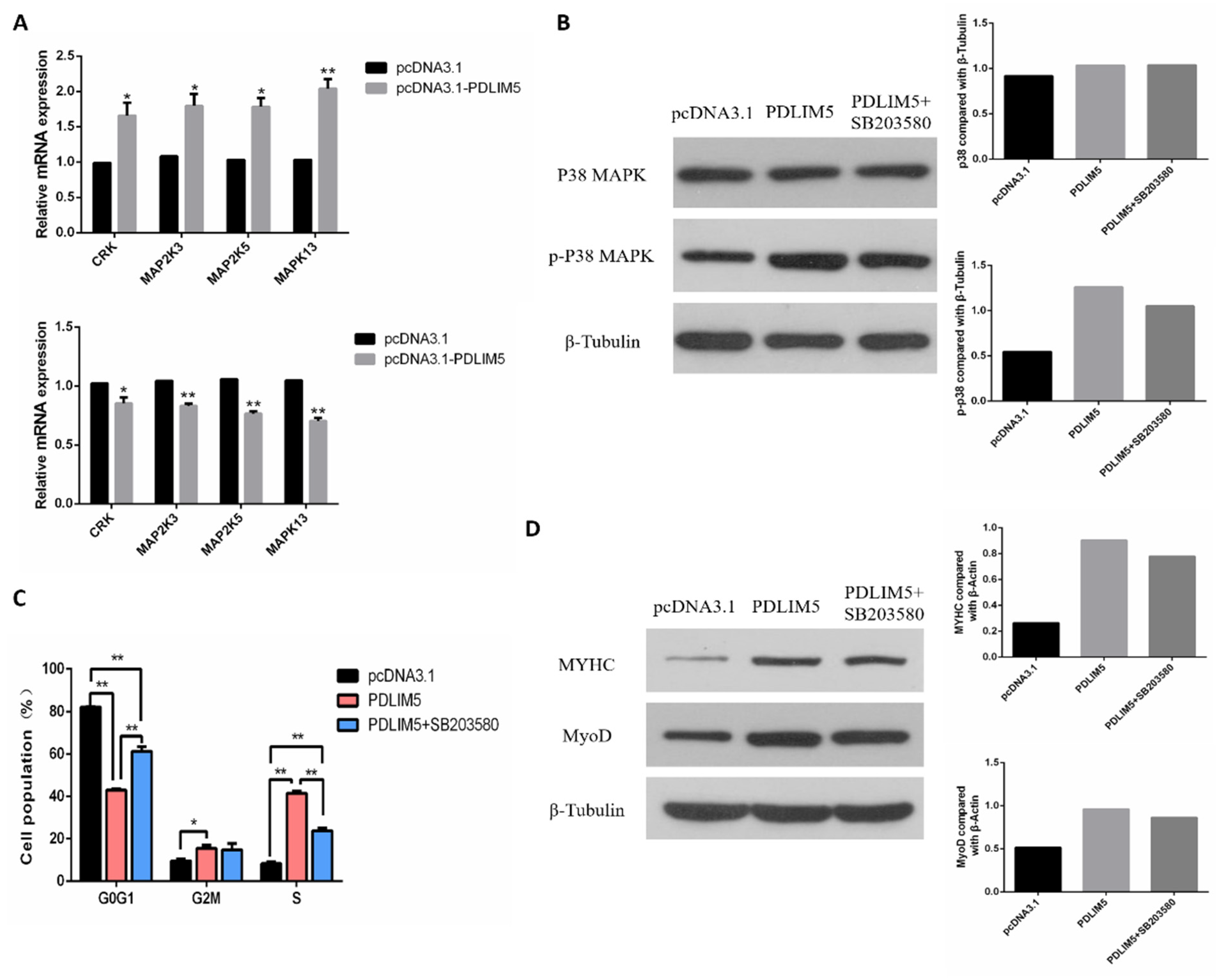
3.5. PDLIM5 Regulated the SMSCs via p38-MAPK Signaling Pathway
4. Discussion
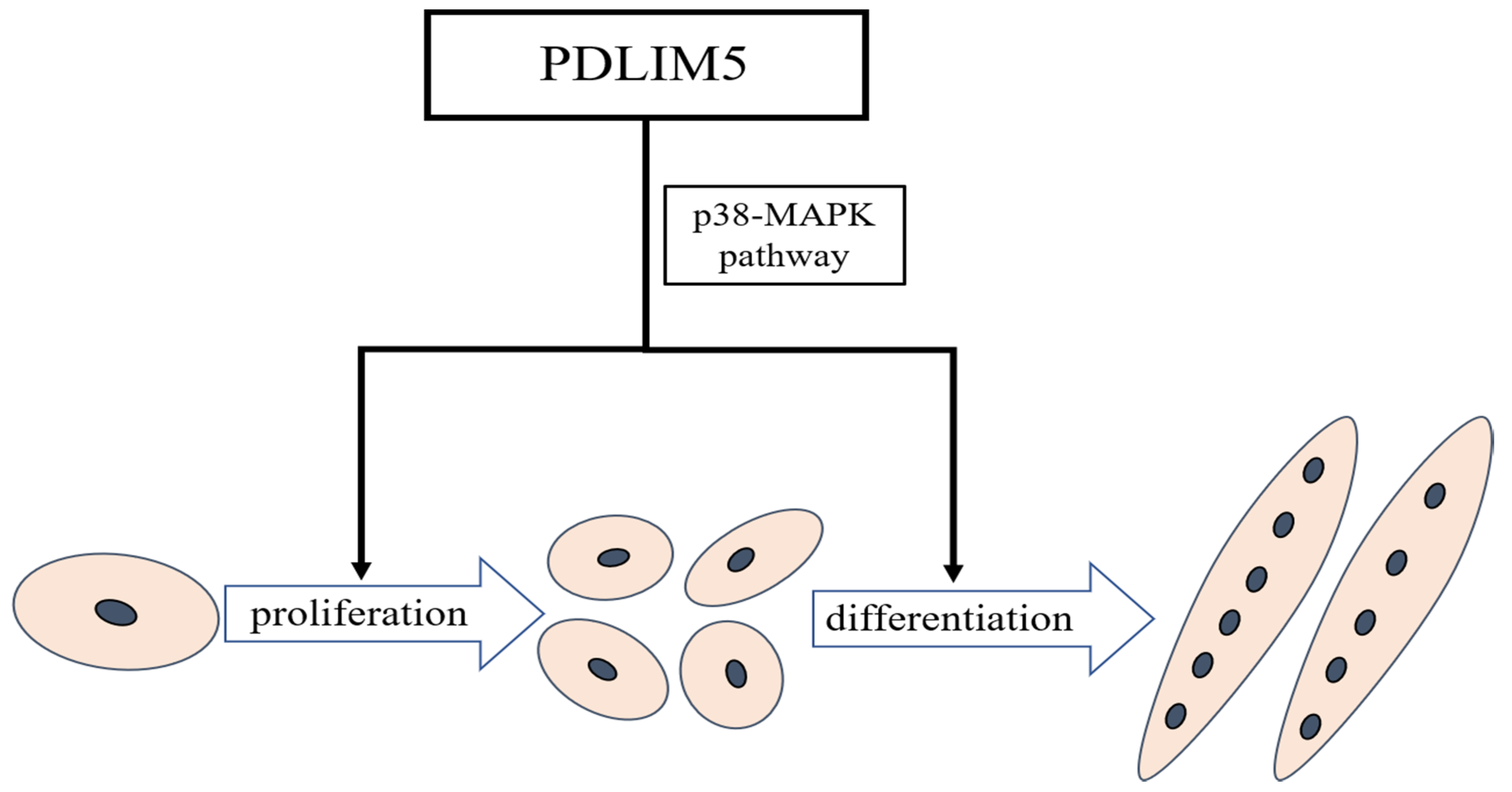
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Li, W.; Liu, D.; Tang, S.; Li, D.; Han, R.; Tian, Y.; Li, H.; Li, G.; Liu, X.; Kang, X.; et al. A multiallelic indel in the promoter region of the cyclin-dependent kinase inhibitor 3 gene is significantly associated with body weight and carcass traits in chickens. Poult. Sci. 2019, 98, 556–565. [Google Scholar] [CrossRef]
- Yin, H.; Zhang, S.; Gilbert, E.R.; Siegel, P.B.; Zhu, Q.; Wong, E.A. Expression profiles of muscle genes in postnatal skeletal muscle in lines of chickens divergently selected for high and low body weight. Poult. Sci. 2014, 93, 147–154. [Google Scholar] [CrossRef]
- Yablonka-Reuveni, Z.; Rivera, A.J. Temporal expression of regulatory and structural muscle proteins during myogenesis of satellite cells on isolated adult rat fibers. Dev. Biol. 1994, 164, 588–603. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.H. Relation of body size to muscle cell size and number in the chicken. Poult. Sci. 1963, 42, 283–290. [Google Scholar] [CrossRef]
- Chargé, S.B.P.; Rudnicki, M.A. Cellular and molecular regulation of muscle regeneration. Physiol. Rev. 2004, 84, 209–238. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, S.; Tokunaga, C.; Kiyohara, Y.; Higuchi, O.; Konishi, H.; Mizuno, K.; Gill, G.N.; Kikkawa, U. Protein-protein interaction of zinc finger lim domains with protein kinase c. J. Biol. Chem. 1996, 271, 31029–31032. [Google Scholar] [CrossRef] [PubMed]
- Zain, M.A.; Roffeei, S.N.; Zainal, N.Z.; Kanagasundram, S.; Mohamed, Z. Nonsynonymous polymorphisms of the pdlim5 gene association with the occurrence of both bipolar disorder and schizophrenia. Psychiatr. Genet. 2013, 23, 258–261. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cheng, H.; Kimura, K.; Peter, A.K.; Cui, L.; Ouyang, K.; Shen, T.; Liu, Y.; Gu, Y.; Dalton, N.D.; Evans, S.M. Loss of enigma homolog protein results in dilated cardiomyopathy. Circ. Res. 2010, 107, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gao, Y.; Xu, Y.; Sun, X.; Song, X.; Ma, H.; Yang, M. Si-rna-mediated knockdown of pdlim5 suppresses gastric cancer cell proliferation in vitro. Chem. Biol. Drug Des. 2015, 85, 447–453. [Google Scholar] [CrossRef]
- Edlund, K.; Lindskog, C.; Saito, A.; Berglund, A.; Pontén, F.; Göransson-Kultima, H.; Isaksson, A.; Jirström, K.; Planck, M.; Johansson, L.; et al. Cd99 is a novel prognostic stromal marker in non-small cell lung cancer. Int. J. Cancer 2012, 131, 2264–2273. [Google Scholar] [CrossRef]
- Eeckhoute, J. A cell-type-specific transcriptional network required for estrogen regulation of cyclin d1 and cell cycle progression in breast cancer. Genes Dev. 2006, 20, 2513. [Google Scholar] [CrossRef] [PubMed]
- Krcmery, J.; Camarata, T.; Kulisz, A.; Simon, H.G. Nucleocytoplasmic functions of the pdz-lim protein family: New insights into organ development. Bioessays News Rev. Mol. Cell. Dev. Biol. 2010, 32, 100–108. [Google Scholar] [CrossRef] [PubMed]
- te Velthuis, A.J.; Bagowski, C.P. Pdz and lim domain-encoding genes: Molecular interactions and their role in development. Sci. World J. 2007, 7, 1470–1492. [Google Scholar] [CrossRef] [PubMed]
- Passier, R.; Richardson, J.A.; Olson, E.N. Oracle, a novel pdz-lim domain protein expressed in heart and skeletal muscle. Mech. Dev. 2000, 92, 277–284. [Google Scholar] [CrossRef]
- Cheng, H.; Chen, T.; Tor, M.; Park, D.; Zhou, Q. A high-throughput screening platform targeting pdlim5 for pulmonary hypertension. J. Biomol. Screen. 2016, 21, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.H.; Grote, E.; Mohler, W.; Vignery, A. Cell-cell fusion. FEBS Lett. 2007, 581, 2181–2193. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Rudnicki, M.A. A new look at the origin, function, and “stem-cell” status of muscle satellite cells. Dev. Biol. 2000, 218, 115–124. [Google Scholar] [CrossRef]
- Yin, H.; He, H.; Cao, X.; Shen, X.; Han, S.; Cui, C.; Zhao, J.; Wei, Y.; Chen, Y.; Xia, L.; et al. Mir-148a-3p regulates skeletal muscle satellite cell differentiation and apoptosis via the pi3k/akt signaling pathway by targeting meox2. Front. Genet. 2020, 11, 512. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Shen, X.; Cao, X.; He, H.; Han, S.; Chen, Y.; Cui, C.; Wei, Y.; Wang, Y.; Li, D.; et al. Hdac4 regulates the proliferation, differentiation and apoptosis of chicken skeletal muscle satellite cells. Animals 2020, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhang, Y.; Yu, S.; Li, S.; Jiang, W.; Zhu, Y.; Xu, Y.; Yang, C.; Tian, G.; Mi, J.; et al. Pdlim5 identified by label-free quantitative proteomics as a potential novel biomarker of papillary thyroid carcinoma. Biochem. Biophys. Res. Commun. 2018, 499, 338–344. [Google Scholar] [CrossRef]
- Liu, X.; Chen, L.; Huang, H.; Lv, J.M.; Chen, M.; Qu, F.J.; Pan, X.W.; Li, L.; Yin, L.; Cui, X.G.; et al. High expression of pdlim5 facilitates cell tumorigenesis and migration by maintaining ampk activation in prostate cancer. Oncotarget 2017, 8, 98117–98134. [Google Scholar] [CrossRef]
- Zhang, G.; Chen, F.; Wu, P.; Li, T.; He, M.; Yin, X.; Shi, H.; Duan, Y.; Zhang, T.; Wang, J.; et al. Microrna-7 targets the klf4 gene to regulate the proliferation and differentiation of chicken primary myoblasts. Front. Genet. 2020, 11, 842. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Cai, B.; Abdalla, B.A.; Zhu, X.; Zheng, M.; Han, P.; Nie, Q.; Zhang, X. Lncirs1 controls muscle atrophy via sponging mir-15 family to activate igf1-pi3k/akt pathway. J. Cachexia Sarcopenia Muscle 2019, 10, 391–410. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cao, X.; Dong, D.; Shen, X.; Cheng, J.; Jiang, R.; Yang, Z.; Peng, S.; Huang, Y.; Lan, X.; et al. Circular rna ttn acts as a mir-432 sponge to facilitate proliferation and differentiation of myoblasts via the igf2/pi3k/akt signaling pathway. Mol. Ther. Nucleic Acids 2019, 18, 966–980. [Google Scholar] [CrossRef] [PubMed]
- Berkes, C.A.; Tapscott, S.J. Myod and the transcriptional control of myogenesis. Semin. Cell Dev. Biol. 2005, 16, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Tajsharghi, H.; Oldfors, A. Myosinopathies: Pathology and mechanisms. Acta Neuropathol. 2013, 125, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Ito, J.; Hashimoto, T.; Nakamura, S.; Aita, Y.; Yamazaki, T.; Schlegel, W.; Takimoto, K.; Maturana, A.D. Splicing transitions of the anchoring protein enh during striated muscle development. Biochem. Biophys. Res. Commun. 2012, 421, 232–238. [Google Scholar] [CrossRef]
- Ito, J.; Takita, M.; Takimoto, K.; Maturana, A.D. Enigma homolog 1 promotes myogenic gene expression and differentiation of c2c12 cells. Biochem. Biophys. Res. Commun. 2013, 435, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Liu, N. Microrna-17-92 regulates myoblast proliferation and differentiation by targeting the enh1/id1 signaling axis. Cell Death Differ. 2016, 23, 1658–1669. [Google Scholar] [CrossRef]
- Keren, A.; Tamir, Y.; Bengal, E. The p38 mapk signaling pathway: A major regulator of skeletal muscle development. Mol. Cell. Endocrinol. 2006, 252, 224–230. [Google Scholar] [CrossRef]
- Jones, N.C.; Tyner, K.J.; Nibarger, L.; Stanley, H.M.; Cornelison, D.D.W.; Olwin, F.B.B. The p38α/β mapk functions as a molecular switch to activate the quiescent satellite cell. J. Cell Biol. 2005, 169, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Zhao, J.; He, H.; Chen, Y.; Wang, Y.; Li, D.; Zhu, Q. Gga-mir-3525 targets pdlim3 through the mapk signaling pathway to regulate the proliferation and differentiation of skeletal muscle satellite cells. Int. J. Mol. Sci. 2020, 21, 5573. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.H.; Mak, G.W.; Li, M.S.; Tsui, S.K. The lim-only protein fhl2 regulates interleukin-6 expression through p38 mapk mediated nf-κb pathway in muscle cells. Cytokine 2012, 59, 286–293. [Google Scholar] [CrossRef]
- Liu, Z.; Han, S.; Wang, Y.; Cui, C.; Zhu, Q.; Jiang, X.; Yang, C.; Du, H.; Yu, C.; Li, Q.; et al. The lim-only protein fhl2 is involved in autophagy to regulate the development of skeletal muscle cell. Int. J. Biol. Sci. 2019, 15, 838–846. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′–3′) |
|---|---|
| Si-PDLIM5 | GGAUAAGUCAGACGGGAUTT AUCCCGUCUGUACUUAUCCTT |
| Si-NC | UUCUCCGAACGUGUCACGUTT ACGUGACACGUUCGGAGAATT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, H.; Yin, H.; Yu, X.; Zhang, Y.; Ma, M.; Li, D.; Zhu, Q. PDLIM5 Affects Chicken Skeletal Muscle Satellite Cell Proliferation and Differentiation via the p38-MAPK Pathway. Animals 2021, 11, 1016. https://doi.org/10.3390/ani11041016
He H, Yin H, Yu X, Zhang Y, Ma M, Li D, Zhu Q. PDLIM5 Affects Chicken Skeletal Muscle Satellite Cell Proliferation and Differentiation via the p38-MAPK Pathway. Animals. 2021; 11(4):1016. https://doi.org/10.3390/ani11041016
Chicago/Turabian StyleHe, Haorong, Huadong Yin, Xueke Yu, Yao Zhang, Menggen Ma, Diyan Li, and Qing Zhu. 2021. "PDLIM5 Affects Chicken Skeletal Muscle Satellite Cell Proliferation and Differentiation via the p38-MAPK Pathway" Animals 11, no. 4: 1016. https://doi.org/10.3390/ani11041016
APA StyleHe, H., Yin, H., Yu, X., Zhang, Y., Ma, M., Li, D., & Zhu, Q. (2021). PDLIM5 Affects Chicken Skeletal Muscle Satellite Cell Proliferation and Differentiation via the p38-MAPK Pathway. Animals, 11(4), 1016. https://doi.org/10.3390/ani11041016