
Housing, Husbandry and Welfare of a “Classic” Fish Model, the Paradise Fish (Macropodus opercularis)

 ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Original Housing and Husbandry Conditions
2.2. Breeding Conditions
2.3. Improved Housing Conditions
2.4. Behavioral Observations
2.5. Health Diagnostics
2.6. Anesthesia and Analgesia
2.7. Data Visualization
3. Results
3.1. Environmental Factors and Optimal Housing Conditions
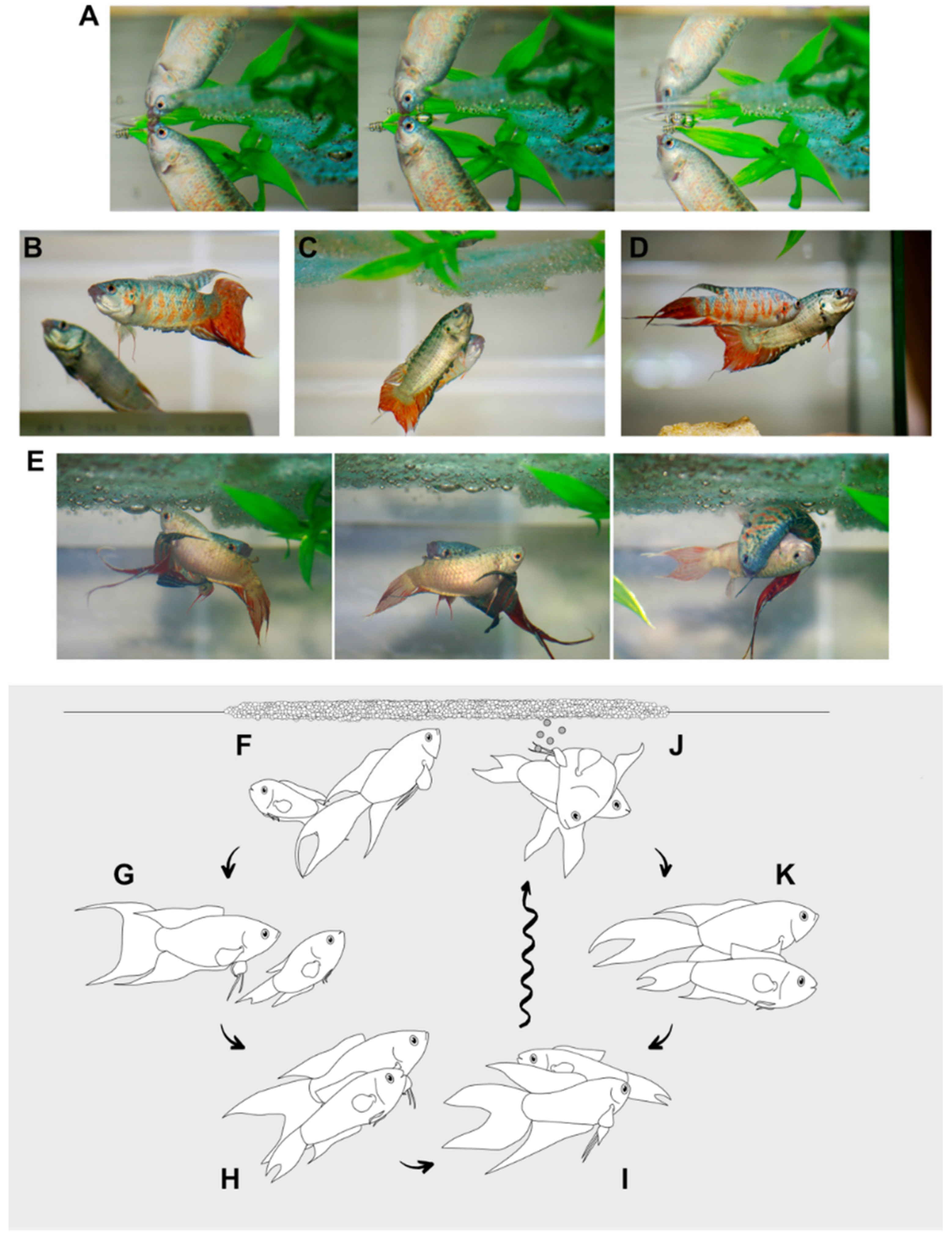
3.2. Social Behavior and Courtship
3.2.1. Species-Specific Behavioral Elements
Frontal Spread Display and Opercular Erection (OPE)
Lateral Spread Display (LSD) and Tail Beating (TAB)
Biting (BIT)
Chasing
Oblique Movements (OBM) and Vertical Waggle (VEW)
Attack and Mouth-Lock (MOU)
3.2.2. Reproductive and Courtship Behaviors
3.3. Effects of Adverse Conditions on Paradise Fish Welfare
3.3.1. Incidence of Stress-Related Disease Symptoms
Results for Pathology and Bacteriology
Results for Molecular Diagnostics (PCR)
3.3.2. Anesthesia and Analgesia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element | Abbreviation | Description | Source |
|---|---|---|---|
| Active elements | |||
| Escape | ESC | Moving rapidly away from strong stimulus source. | [16] |
| Swim | SWI | Normal or fast locomotion with no special orientation. | [16] |
| Move | MOV | Slow, short-range locomotion with no special orientation. | [16] |
| Creeping | CRE | Slow, forward movement propelled only by the pectoral fin. | [16] |
| Staccato | STA | Quick starts and sudden stops, with no special orientation. | [16] |
| Leaping | LEP | Quick move, propelled by the caudal fin. | [16] |
| Erratic movement | ERA | Zig-zag locomotion performed on the bottom. | [16] |
| Jumping | JUP | Quick jump from a confined space, out of the water | [16] |
| Air gulping | A-G | Air gulping at the surface. | [16] |
| Air bubbling | A-B | Similar to air gulping but a few big bubbles are released through the gills or mouth. Most often associated with nest-building. | [16] |
| Approach | APR | Slow movement, oriented to a particular object. | [16] |
| Pick | PIC | Object picked with the jaw. | [16] |
| Back | BAC | Slow, backward locomotion. | [16] |
| Passive elements | |||
| Floating | FLO | The fish is floating 1–2 cm under the surface. | [16] |
| Hanging in midwater | HIM | The fish is floating > 2 cm under the surface. | [16] |
| Resting | RES | Fish rests at the bottom, with pectoral fins fanning slowly. | [16] |
| Freezing | FRZ | Fish is motionless at the bottom. | [16] |
| Freezing under the surface | FUS | Fish is periodically motionless near the surface, and gulps air in-between. | [16] |
| Social elements | |||
| Social orientation | ORI | Slow locomotion around a conspecific. | [16] |
| Oblique plan floating position 1 | OBF | Submission posture, the body is 20–40 degrees inclined from the horizontal plane, only pectoral fins are fanning, other fins are closed. | [16] |
| Lateral head-head display 2,3 | LHH | Fish are oriented in the same direction with one being slightly behind. | [18] |
| Lateral head-tail display 2,3 | LHT | Fish (usually males) are oriented in opposite directions. Sometimes they also slowly start circling. | [18] |
| Lateral head-tail display with shaking 2 | SHA | Similar to LHT but involves fast circling and descent to the bottom. | [18] |
| Frontal Display 2 | FRD | Faces the opponent with unpaired fins extended. Body angle towards the opponent close to perpendicular. | [5,18] |
| Opercular Erection 2 | OPE | Opercular erection, the opercular ocelli are visible. Usually performed by competing males during LHH, LHT or FRD. | [5,18] |
| Biting | BIT | The fish uses its jaws to inflict wounds by damaging the epidermis of the other fish | [5] |
| Charge | CHA | The fish swims fast in the direction of the opponent. | Described in the context of B. splendens [69] |
| Display at distance | DIS | Fish stay in head-tail position, with erected tailfin. Distance is larger than one body length. | [18] |
| Parallel swimming | PAS | The two fish swim closely to each other into the same direction. | [18] |
| Tail-beating | TAB | Occurs occasionally during LHH and LHT and consists of undulating thrusts of the tail towards the other fish | [5] |
| Mouth-lock | MOU | Occurs during intense fighting after FRD and OPE. Males lock their jaws and sink to the bottom of the tank. It can last from several seconds up to 2 min. | [18] |
| Lateral vibrating | VIB | Rapid undulatory movement that occurs when the pair is in head-to-head or head-to-tail position. Usually used by non-receptive females. | [5] |
| Circling | CIR | Male curves his body and slowly swims in a circle. Female follows with her snout at his dorsum. | [5] |
| Oblique plan movements | OBM | Median fins are commonly folded, the caudal fin often droops, and the fish often tilts laterally. Occasionally, the body may be tilted vertically, either upward or downward. | [5] |
| Vertical waggle | VEW | Low-frequency undulating movements of the caudal fin while the fish is in (near) vertical position. Often performed by receptive females. | [5] |
| Leading to the nest | LEN | After an LSD male swims slowly to the nest, with median fins erect. | [5] |
| Anabantoid embrace 4 | ANE | Female moves into a U-shape flexture, with her snout protruding beyond the male’s dorsal fin. The male clasps the female, quivering of the body starts in both and they roll over. Genital pores move close to the bubble nest, gametes are released. | [5,7,11] |
| Swimming inhibition | SIN | After spawning, male releases the female and both become immobile, even start to sink. | [5] |
Appendix B
Appendix B.1. Detailed Fin-Clipping Protocol
Appendix B.1.1. Required Reagents and Tools
- Lidocaine powder (Sigma L7757);
- Ethyl 3-aminobenzoate methane sulfonate/Tricaine powder (Sigma E10521);
- Ethanol, min. 70%;
- Fin-clipping board + blade + forceps;
- Anesthesia tank (anesthetic and analgesic);
- Recovery tank (analgesia and treatment tank).
Appendix B.1.2. Stock Solutions
- Lidocaine stock solution (500 mg/L) prepared fresh weekly with filtered water (or purified reverse osmosis (RO) water). Keep the stock solution in the dark or in a covered glass bottle at 4 °C.
- Tricaine (MS-222) stock solution (4 g/L final concentration) [70], containing 400 mg of MS-222 in 97.9 mL purified or reverse osmosis (RO) water: buffered to pH 7.5 with 2.1 mL of 1 M Tris (pH 9). Keep the stock solution at 4 °C for use in 1–2 weeks or at −20 °C for prolonged storage.
Appendix B.2. Procedure
References
- Yu, T.; Guo, Y. Early Normal Development of the Paradise Fish Macropodus opercularis. Russ. J. Dev. Biol. 2018, 49, 240–244. [Google Scholar] [CrossRef]
- Huang, W.B.; Chang, C.C. Effects of Parental Care and Body Size on the Reproductive Success of the Paradise Fish Macropodus opercularis (L.) in a Small Area. Zool. Stud. 2011, 50, 401–408. [Google Scholar]
- Ward, R.W. Ethology of the Paradise Fish, Macropodus opercularis I. Differences between Domestic and Wild Fish. Copeia 1967, 1967, 809. [Google Scholar] [CrossRef]
- Haung, W.B.; Cheng, F.L. Effects of Temperature and Floating Materials on Breeding by the Paradise Fish (Macropodus opercularis) in the Non-Reproductive Season. Zool. Stud. 2006, 45, 475–482. [Google Scholar]
- Hall, D.D. A Qualitative Analysis of Courtship and Reproductive Behavior in the Paradise Fish, Macropodus opercularis (Linnaeus). Z. Tierpsychol. 1968, 25, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Miklosi, A.; Csanyi, V.; Gerlai, R. Antipredator Behavior in Paradise Fish (Macropodus opercularis) Larvae: The Role of Genetic Factors and Paternal Influence. Behav. Genet. 1997, 27, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Tate, M.; McGoran, R.E.; White, C.R.; Portugal, S.J. Life in a Bubble: The Role of the Labyrinth Organ in Determining Territory, Mating and Aggressive Behaviours in Anabantoids. J. Fish Biol. 2017, 91, 723–749. [Google Scholar] [CrossRef] [PubMed]
- Watson, C.A.; DiMaggio, M.; Hill, J.E.; Tuckett, Q.M.; Yanong, R.P. Evolution, Culture, and Care for Betta splendens. EDIS 2019, 2019. [Google Scholar] [CrossRef]
- Cole, B.; Tamaru, C.S.; Bailey, R.; Brown, C. A Manual for Commercial Production of the Gourami, Trichogaster Trichopterus, A Temporary Paired Spawner; Publication Number 135-2-Acknowledgements; Center for Tropical and Subtropical Aquaculture: Honolulu, HI, USA, 1999. [Google Scholar]
- Liu, T.-C.; Cheng, C.-F.; Chang, C.-H.; Lin, S.-L.; Wang, W.-S.; Hung, S.-W.; Chen, M.-H.; Lin, C.-C.; Tu, C.-Y.; Lin, Y.-H. Highly Virulent Photobacterium Damselae Subsp. Piscicida Isolated from Taiwan Paradise Fish, Macropodus opercularis (L.), in Taiwan. Afr. J. Microbiol. Res. 2011, 5, 2107–2113. [Google Scholar] [CrossRef]
- Chandran, B.K.S.; Jayaprakas, V.; Kumar, A.B. Breeding behaviour of spiketail paradise fish, Pseudosphromenus cupanus (Cuvier, 1831). Int. J. Pure Appl. Zool. 2013, 1, 267–276. [Google Scholar]
- Song, H.-B.; Choi, S.-S. Reproductive Ecology and Early Life History of Paradise Fish, Macropodus chinensis (Pisces; Belontidae) in Aquarium. Korean J. Limnol. 2000, 33, 282–294. [Google Scholar]
- Scheurmann, I. Aquarium Fish Breeding, 1st ed.; Baron’s Educational Series Inc. Publishing: Hauppauge, NY, USA, 1990. [Google Scholar]
- Srikrishnan, R.; Hirimuthugoda, N.; Rajapakshe, W. Evaluation of Growth Performance and Breeding Habits of Fighting Fish (Betta splendens) under 3 Diets and Shelters. J. Surv. Fish. Sci. 2017, 3, 50–65. [Google Scholar] [CrossRef]
- Motilan, Y.; Nishikanta, K. Captive Breeding of Thick-Lipped Gourami, Trichogaster labiosua (Day) by Gradual Increasing Aquarium Water Temperature and Their Early Life Stages. Int. J. Sci. Res. Publ. 2014, 5(3), 1–4. [Google Scholar]
- Csányi, V.; Tóth, P.; Altbäcker, V.; Dóka, A.; Gervai, J. Behavioural Elements of the Paradise Fish (Macropodus opercularis). I. Regularities of Defensive Behaviour. Acta Biol. Hung. 1985, 36, 93–114. [Google Scholar]
- Csányi, V.; Tóth, P.; Altbäcker, V.; Dóka, A.; Gervai, J. Behavioural Elements of the Paradise Fish (Macropodus opercularis). II. A Functional Analysis. Acta Biol. Hung. 1985, 36, 115–130. [Google Scholar]
- Miklosi, A.; Haller, J.; Csanyi, V. The Influence of Opponent-Related and Outcome-Related Memory on Repeated Aggressive Encounters in the Paradise Fish (Macropodus opercularis). Biol. Bull. 1995, 188, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Miklósi, Á.; Haller, J.; Csányi, V. Learning about the Opponent during Aggressive Encounters in Paradise Fish (Macropodus opercularis L.): When It Takes Place? Behav. Process. 1997, 40, 97–105. [Google Scholar] [CrossRef]
- Haller, J.; Miklósi, Á.; Csányi, V.; Makara, G.B. Behavioral Tactics Control the Energy Costs of Aggression: The Example of Macropodus opercularis. Aggress. Behav. 1996, 22, 437–446. [Google Scholar] [CrossRef]
- Csányi, V.; Csizmadia, G.; Miklosi, A. Long-Term Memory and Recognition of Another Species in the Paradise Fish. Anim. Behav. 1989, 37, 908–911. [Google Scholar] [CrossRef]
- Miklósi, A.; Topál, J.; Csányi, V. Development of Open-field and Social Behavior of the Paradise Fish (Macropodus opercularis L.). Dev. Psychobiol. 1992, 25, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Miklósi, Á.; Haller, J.; Csányi, V. Different Duration of Memory for Conspecific and Heterospecific Fish in the Paradise Fish (Macropodus opercularis L.). Ethology 2010, 90, 29–36. [Google Scholar] [CrossRef]
- Miklósi, Á.; Pongrácz, P.; Csányi, V. The Ontogeny of Antipredator Behavior in Paradise Fish Larvae (Macropodus opercularis) IV. The Effect of Exposure to Siblings. Dev. Psychobiol. 1997, 30, 283–291. [Google Scholar] [CrossRef]
- Kassel, J.; Davis, R.E. Early Behavioral Experience and Adult Social Behavior in the Paradise Fish, Macropodus opercularis L. Behav. Biol. 1975, 15, 343–351. [Google Scholar] [CrossRef]
- Davis, R.E.; Kassel, J. The Ontogeny of Agonistic Behavior and the Onset of Sexual Maturation in the Paradise Fish, Macropodus opercularis (Linnaeus). Behav. Biol. 1975, 14, 31–39. [Google Scholar] [CrossRef]
- Huntingford, F.A.; Adams, C.; Braithwaite, V.A.; Kadri, S.; Pottinger, T.G.; Sandøe, P.; Turnbull, J.F. Current Issues in Fish Welfare. J. Fish Biol. 2006, 68, 332–372. [Google Scholar] [CrossRef]
- Sloman, K.A.; Bouyoucos, I.A.; Brooks, E.J.; Sneddon, L.U. Ethical Considerations in Fish Research. J. Fish Biol. 2019, 94, 556–577. [Google Scholar] [CrossRef]
- Pleeging, C.C.F.; Moons, C.P.H. Potential Welfare Issues of the Siamese Fighting Fish (Betta splendens) at the Retailer and in the Hobbyist Aquarium. Vlaams Diergeneeskd. Tijdschr. 2017, 86, 213–223. [Google Scholar] [CrossRef]
- Brown, C. Fish Intelligence, Sentience and Ethics. Animal Cognition; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1–17. [Google Scholar] [CrossRef]
- Chandroo, K.P.; Yue, S.; Moccia, R.D. An Evaluation of Current Perspectives on Consciousness and Pain in Fishes. Fish Fish. 2004, 5, 281–295. [Google Scholar] [CrossRef]
- Brown, C.; Laland, K.; Krause, J.; Brown, C. (Eds.) Fish Cognition and Behavior; Wiley-Blackwell: Oxford, UK, 2011. [Google Scholar] [CrossRef]
- Demski, L.S. The Pallium and Mind/Behavior Relationships in Teleost Fishes. Brain. Behav. Evol. 2013, 82, 31–44. [Google Scholar] [CrossRef]
- Håstein, T.; Scarfe, A.D.; Lund, V.L. Science-Based Assessment of Welfare: Aquatic Animals. Oie Rev. Sci. Tech. 2005, 24, 529–547. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goodfellow, M. Nucleic Acid Techniques in Bacterial Systematics; John Wiley & Sons, Ltd.: Chichester, UK; New York, NY, USA, 1991. [Google Scholar]
- Mocho, J.P. Three-Dimensional Screen: A Comprehensive Approach to the Health Monitoring of Zebrafish. Zebrafish 2016, 13, S132–S137. [Google Scholar] [CrossRef] [PubMed]
- Mocho, J.P.; Martin, D.J.; Millington, M.E.; Saavedra Torres, Y. Environmental Screening of Aeromonas hydrophila, Mycobacterium spp., and Pseudocapillaria tomentosa in Zebrafish Systems. J. Vis. Exp. 2017, 2017, 1–9. [Google Scholar] [CrossRef]
- Schroeder, P.G.; Sneddon, L.U. Exploring the Efficacy of Immersion Analgesics in Zebrafish Using an Integrative Approach. Appl. Anim. Behav. Sci. 2017, 187, 93–102. [Google Scholar] [CrossRef]
- Deakin, A.G.; Buckley, J.; AlZu’bi, H.S.; Cossins, A.R.; Spencer, J.W.; Al’Nuaimy, W.; Young, I.S.; Thomson, J.S.; Sneddon, L.U. Automated Monitoring of Behaviour in Zebrafish after Invasive Procedures. Sci. Rep. 2019, 9, 9042. [Google Scholar] [CrossRef] [PubMed]
- Sneddon, L.U. Evolution of Nociception and Pain: Evidence from Fish Models. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374. [Google Scholar] [CrossRef] [PubMed]
- Sneddon, L. Clinical Anesthesia and Analgesia in Fish. J. Exot. Pet Med. 2012, 21, 32–43. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 5 July 2019).
- Wickham, H. ggplot2 — Elegant Graphics for Data Analysis. Available online: https://www.springer.com/gp/book/9780387981413 (accessed on 5 March 2020). [CrossRef]
- Mandal, S.C.; Sahu, N.P.; Kohli, M.P.S.; Das, P.; Gupta, S.K.; Munilkumar, S. Replacement of Live Feed by Formulated Feed: Effect on the Growth and Spawning Performance of Siamese Fighting Fish (Betta splendens, Regan, 1910). Aquac. Res. 2010, 41, 1707–1716. [Google Scholar] [CrossRef]
- Puello-Cruz, A.C.; Velasco-Blanco, G.; Martínez-Rodríguez, I.E.; Felix-Ramos, E.; Voltolina, D. Growth and Survival of Siamese Fighting Fish, Betta splendens, Larvae at Low Salinity and With Different Diets. J. World Aquac. Soc. 2010, 41, 823–828. [Google Scholar] [CrossRef]
- Sipaúba-Tavares, L.H.; Appoloni, A.M.; Fernandes, J.B.K.; Millan, R.N. Alimentação Do Peixe-de-Briga, Betta splendens (Regan, 1910) Em Tanque Externo de Cultivo: Dieta Natural e Formulada. Braz. J. Biol. 2016, 76, 292–299. [Google Scholar] [CrossRef] [PubMed]
- McMenamin, S.K.; Parichy, D.M. Metamorphosis in Teleosts. In Current Topics in Developmental Biology; Academic Press Inc.: Cambridge, MA, USA, 2013; Volume 103, pp. 127–165. [Google Scholar] [CrossRef]
- Sifa, L.; Mathias, J.A. The Critical Period of High Mortality of Larvae Fish -A Discussion Based on Current Research. Chin. J. Oceanol. Limnol. 1987, 5, 80–96. [Google Scholar] [CrossRef]
- Lawrence, C. The Husbandry of Zebrafish (Danio rerio): A Review. Aquaculture 2007, 269, 1–20. [Google Scholar] [CrossRef]
- Reed, B.; Jennings, M. Guidance on the Housing and Care of Zebrafish Danio rerio; RSPCA: West Sussex, UK, 2010. [Google Scholar]
- Vargesson, N. Zebrafish. In Manual of Animal Technology; Barnett, S.W., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2007. [Google Scholar]
- Varga, M.; Ralbovszki, D.; Balogh, E.; Hamar, R.; Keszthelyi, M.; Tory, K. Zebrafish Models of Rare Hereditary Pediatric Diseases. Diseases 2018, 6, 43. [Google Scholar] [CrossRef] [PubMed]
- MacRae, C.A.; Peterson, R.T. Zebrafish as Tools for Drug Discovery. Nat. Rev. Drug Discov. 2015, 14, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Kinth, P.; Mahesh, G.; Panwar, Y. Mapping of Zebrafish Research: A Global Outlook. Zebrafish 2013, 10, 510–517. [Google Scholar] [CrossRef]
- Patton, E.E.; Tobin, D.M. Spotlight on Zebrafish: The next Wave of Translational Research. DMM Dis. Model. Mech. 2019, 12, dmm039370. [Google Scholar] [CrossRef] [PubMed]
- Müller, F. Comparative Aspects of Alternative Laboratory Fish Models. Zebrafish 2005, 2, 47–54. [Google Scholar] [CrossRef]
- Schartl, M. Beyond the Zebrafish: Diverse Fish Species for Modeling Human Disease. DMM Dis. Model. Mech. 2014, 7, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Gervai, J.; Csányi, V. Behavior-Genetic Analysis of the Paradise Fish, Macropodus Opercularis. I. Characterization of the Behavioral Responses of Inbred Strains in Novel Environments: A Factor Analysis. Behav. Genet. 1985, 15, 503–519. [Google Scholar] [CrossRef] [PubMed]
- Csányi, V.; Gerlai, R. Open-Field Behavior and the Behavior-Genetic Analysis of the Paradise Fish (Macropodus opercularis). J. Comp. Psychol. 1988, 102, 326–336. [Google Scholar] [CrossRef]
- Csányi, V. Ethological Analysis of Predator Avoidance by the Paradise Fish (Macropodus opercularis L.): II. Key Stimuli in Avoidance Learning. Anim. Learn. Behav. 1986, 14, 101–109. [Google Scholar] [CrossRef]
- Matthews, B.J.; Vosshall, L.B. How to Turn an Organism into a Model Organism in 10 ‘easy’ Steps. J. Exp. Biol. 2020, 223, jeb218198. [Google Scholar] [CrossRef]
- Williams, T.D.; Readman, G.D.; Owen, S.F.; Williams, T. Key Issues Concerning Environmental Enrichment for Laboratory-Held Fish Species. Animals 2009, 43, 107–120. [Google Scholar] [CrossRef]
- Tort, L. Stress and Immune Modulation in Fish. Dev. Comp. Immunol. 2011, 35, 1366–1375. [Google Scholar] [CrossRef] [PubMed]
- Wendelaar Bonga, S.E. The Stress Response in Fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Hirayama, K. Water Control by Filtration in Closed Culture Systems. Aquaculture 1974, 4, 369–385. [Google Scholar] [CrossRef]
- Manuel, R.; Gorissen, M.; Stokkermans, M.; Zethof, J.; Ebbesson, L.O.E.; Van Den Vis, H.; Flik, G.; Van Den Bos, R. The Effects of Environmental Enrichment and Age-Related Differences on Inhibitory Avoidance in Zebrafish (Danio rerio Hamilton). Zebrafish 2015, 12, 152–165. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.M. Psychological Factors in Stress and Disease. Sci. Am. 1972, 226, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Clotfelter, E.D.; Ardia, D.R.; McGraw, K.J. Red Fish, Blue Fish: Trade-Offs between Pigmentation and Immunity in Betta splendens. Behav. Ecol. 2007, 18, 1139–1145. [Google Scholar] [CrossRef]
- Magnadottir, B. Immunological Control of Fish Diseases. Mar. Biotechnol. 2010, 12, 361–379. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. A Guide for the Laboratory Use of Zebrafish (Danio rerio), 4th ed.; University of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]




| Water Quality Parameters and Husbandry Routine | |
|---|---|
| Air temperature | 24.0–28.0 °C |
| pH | 7.0–7.5 |
| Conductivity | 350–450 µS |
| NO2− | <0.06 mg/L |
| NO3− | <15–20 mg/L |
| NH3 (ammonia) | <0.05 mg/L |
| Fish feeding < 5 months | 3× daily |
| Fish feeding > 5 months | 2× daily |
| Tank cleaning larvae | <1 month—daily |
| Tank cleaning juvenile | >1 month—weekly |
| Tank cleaning >5 months (adults) | weekly (clean and siphon), with half water volume change |
| Adult fish holding density | 10–12 fish/30 L |
| Light cycle | 13 h Light:11 h Dark |
| 3 L Tanks, 10 Larvae/3 dL, Artemia from 3 wpf | 3 L Tanks, 7 Larvae/3 dL, Artemia from 3 wpf | 3 L Tanks, 5 Larvae/3 dL, Artemia from 6 dpf | 3 L Tanks, 2–4 Larvae/3 dL, Artemia from 6 dpf | 5 L Tanks, 1–1.5 Larvae/dL with Sponge-Water Filter | 10 L Tanks, 1–2 Larvae/dL with Sponge-Water Filter |
|---|---|---|---|---|---|
| ~1% (4/400) | ~9% (31/350) | ~28% (99/350) | ~36% (65/180) | 0% 1 (0/360) | 0% 1 (0/450) |
| Behavior | Male | Female |
|---|---|---|
| Nest-building + Courtship | Spread fins (DIS or LHT) Opercular erection (OPE) Brightening body color Territorial defense (BIT) Bubble nest build (A-B) Intense lateral spread display (LHH or LHT) Leads female to nest (LEN) | Refusal: Spread fins (DIS or LHT), Opercular erection (OPE), Swim away, hide (SWI or ESC) Acceptance: Pale body color, Oblique movements (OBM), Vertical waggle (VEW), Follow male |
| Spawning | Circle under nest (CIR) Anabantoid embrace (ANE) Ejaculation + Sinking (SIN) Release female | Circle under nest (CIR) Anabantoid embrace (ANE) Egg release + sinking (SIN) |
| Nest care/Guard | Collect eggs and put them into the nest (PIC) Blow more bubble (A-B) Chase away female (BIT) Guard nest (BIT) Clean and move eggs and embryos (PIC) | Either tries to help the male and is chased away (ESC) or Uninterested in nest care from start |
| Pathogen | Bacteriology | PCR |
|---|---|---|
| Aeromonas hydrophila | + | + |
| Pseudomonas alcaligenes | + | ND |
| Aerococcus spp. | + | ND |
| Comamonas spp. | + | ND |
| Mycobacterium spp. | ND | + |
| Mycobacterium chelonae | ND | + |
| Mycobacterium fortuitum | ND | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rácz, A.; Adorján, G.; Fodor, E.; Sellyei, B.; Tolba, M.; Miklósi, Á.; Varga, M. Housing, Husbandry and Welfare of a “Classic” Fish Model, the Paradise Fish (Macropodus opercularis). Animals 2021, 11, 786. https://doi.org/10.3390/ani11030786
Rácz A, Adorján G, Fodor E, Sellyei B, Tolba M, Miklósi Á, Varga M. Housing, Husbandry and Welfare of a “Classic” Fish Model, the Paradise Fish (Macropodus opercularis). Animals. 2021; 11(3):786. https://doi.org/10.3390/ani11030786
Chicago/Turabian StyleRácz, Anita, Gábor Adorján, Erika Fodor, Boglárka Sellyei, Mohammed Tolba, Ádám Miklósi, and Máté Varga. 2021. "Housing, Husbandry and Welfare of a “Classic” Fish Model, the Paradise Fish (Macropodus opercularis)" Animals 11, no. 3: 786. https://doi.org/10.3390/ani11030786
APA StyleRácz, A., Adorján, G., Fodor, E., Sellyei, B., Tolba, M., Miklósi, Á., & Varga, M. (2021). Housing, Husbandry and Welfare of a “Classic” Fish Model, the Paradise Fish (Macropodus opercularis). Animals, 11(3), 786. https://doi.org/10.3390/ani11030786