
Swine Inflammation and Necrosis Syndrome Is Associated with Plasma Metabolites and Liver Transcriptome in Affected Piglets

 , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animals and Experimental Design
2.3. Clinical Scoring
2.4. Sample Collection
2.5. RNA Extraction, Microarray Hybridization, and Bioinformatic Analysis
2.6. Validation of Microarray Data Using qPCR Analysis
2.7. Targeted Metabolite Screening
2.8. Statistics
3. Results
3.1. Clinical SINS Scores and Organ Alterations
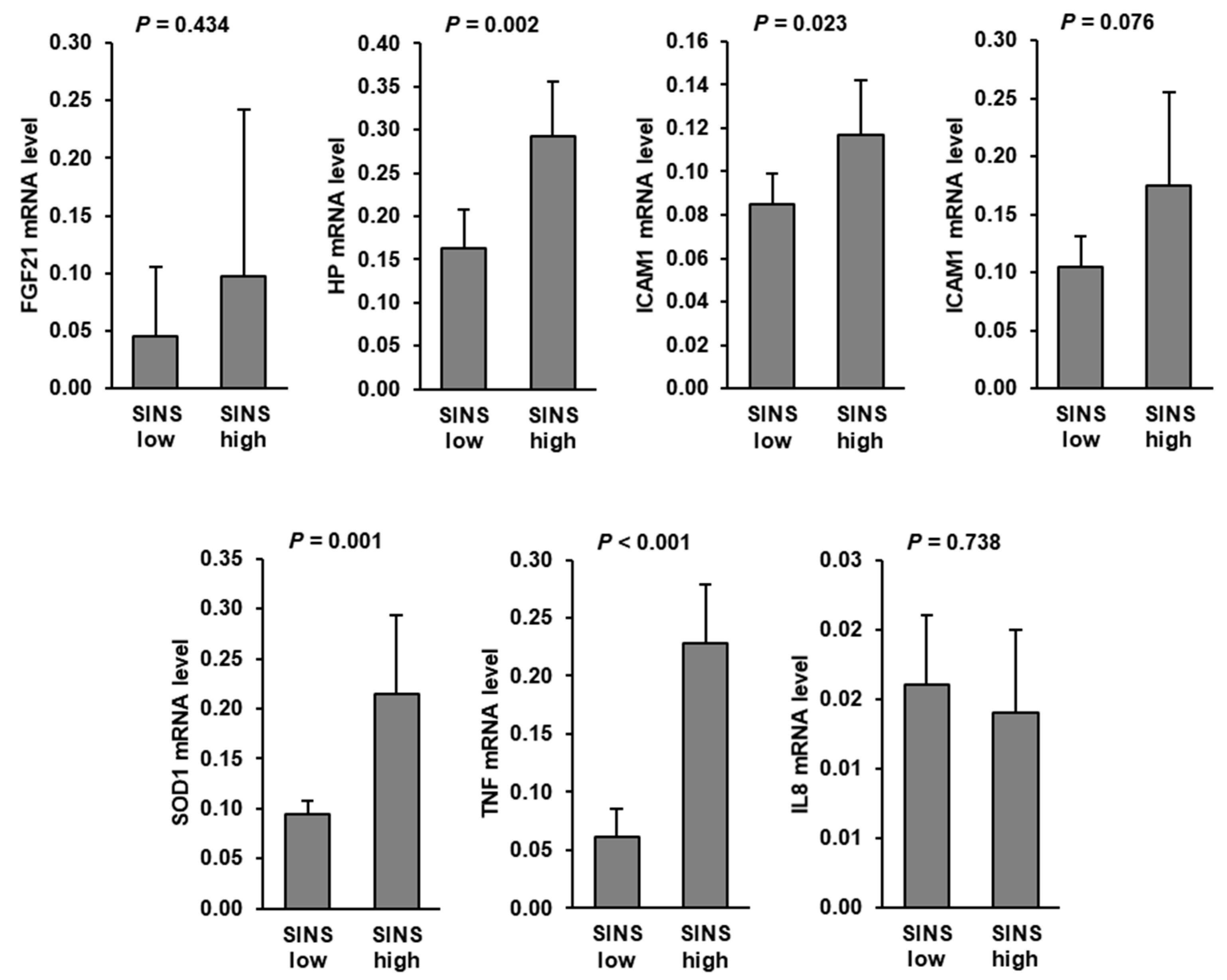
3.2. Expression of Genes Involved in Inflammation and Stress Response in the Liver of Piglets of the SINS Low Group and the SINS High Group
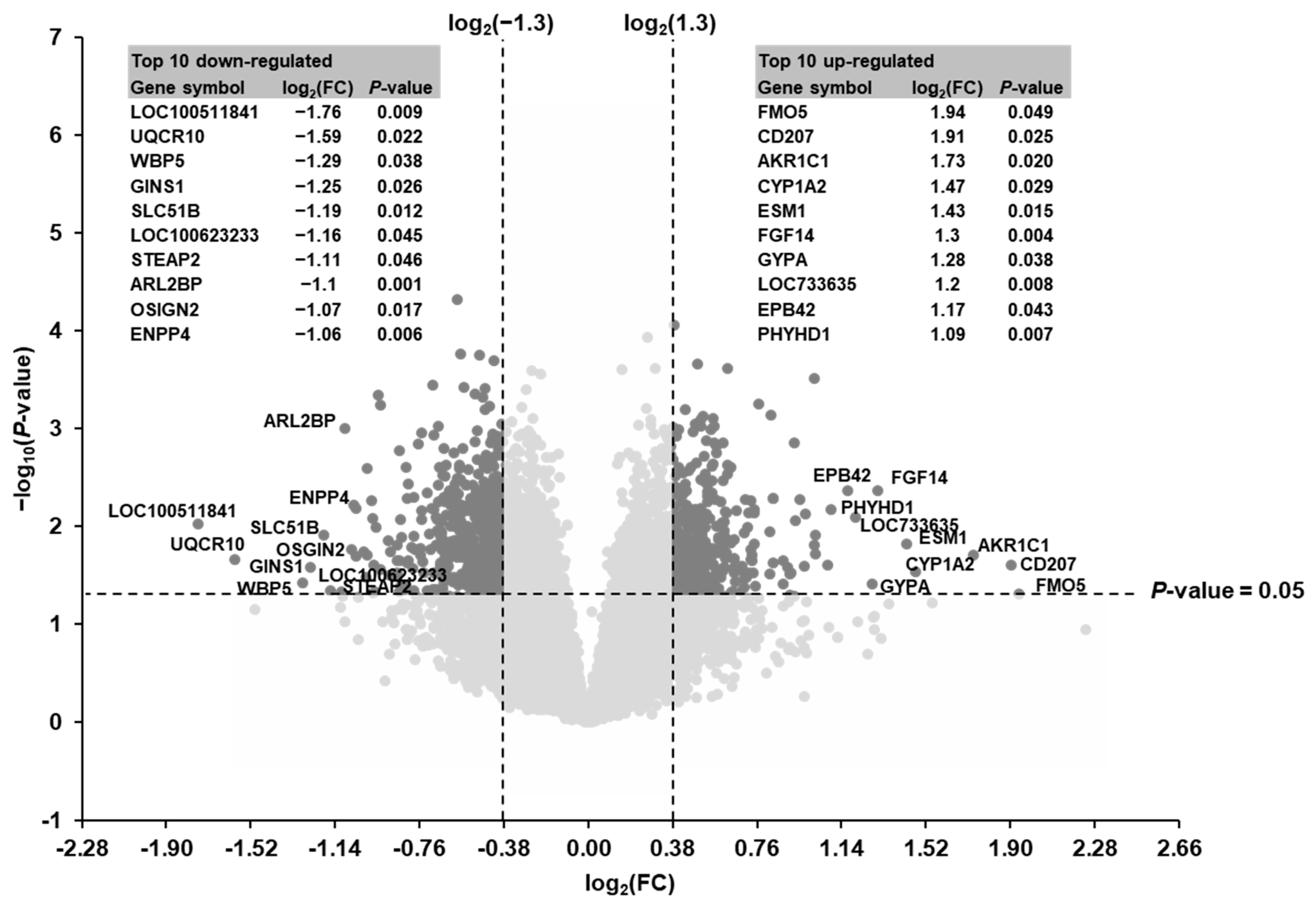
3.3. Identification of Differentially Expressed Transcripts in the Liver of Piglets between the SINS High Group and the SINS Low Group
3.4. Technical Validation of Microarray Data
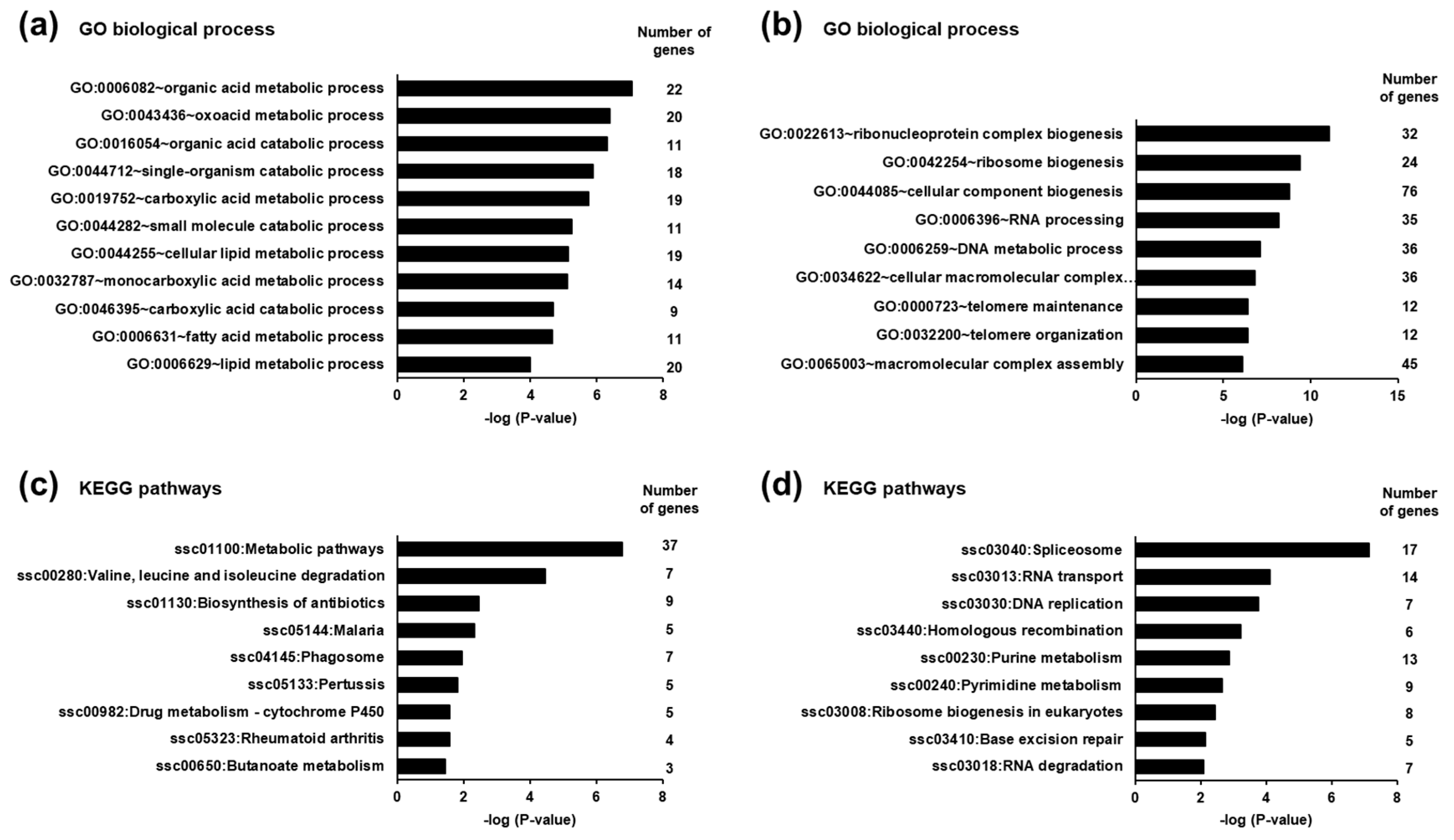
3.5. Identification of Biological Processes and Pathways Affected by the Differentially Expressed Transcripts in the Liver of Piglets between the SINS High Group and the SINS Low Group
3.6. Concentrations of Plasma Metabolites in Suckling Piglets of the SINS High Group and the SINS Low Group
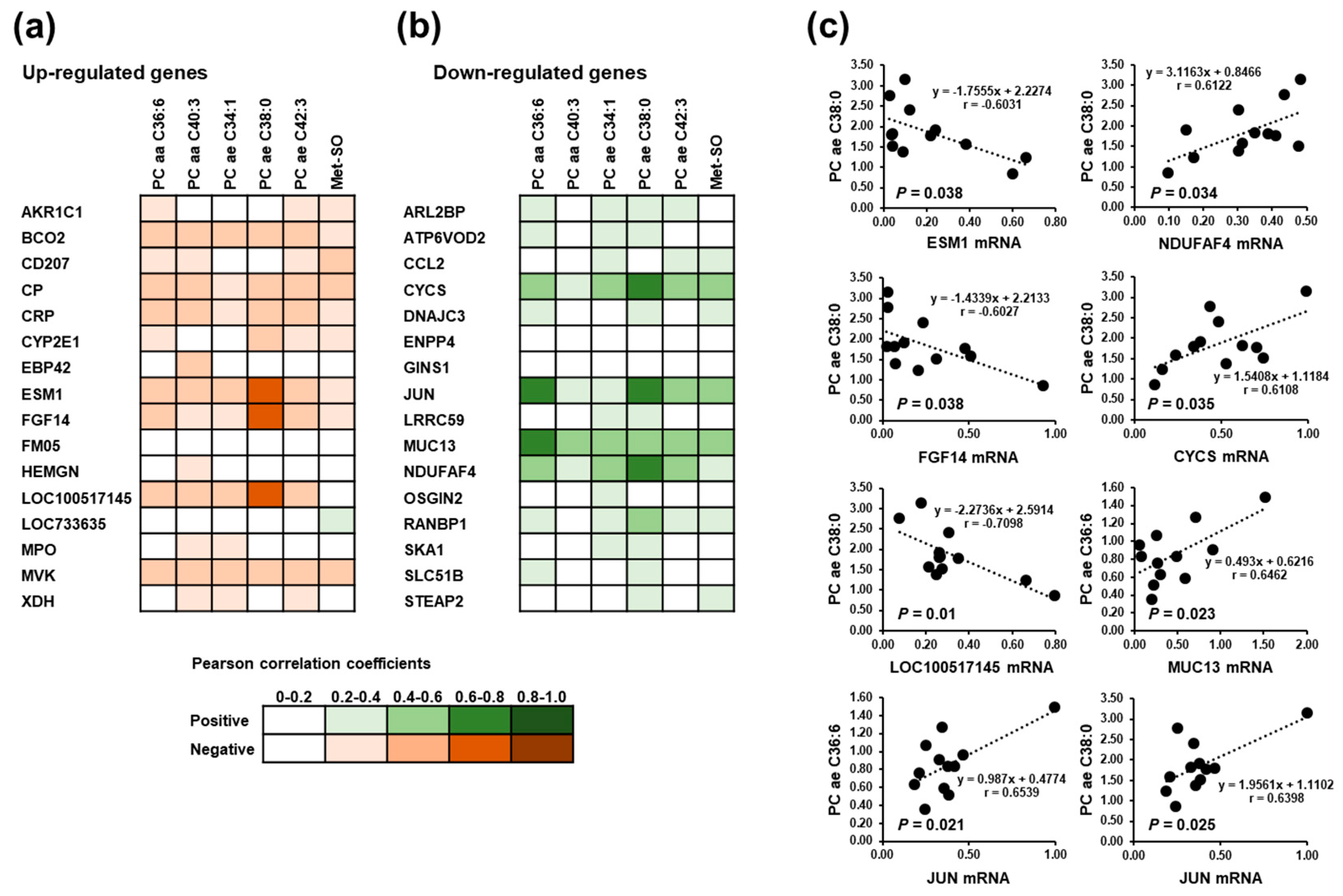
3.7. Correlations between Significantly Altered Plasma Metabolites and the Most Strongly Regulated Hepatic Genes in Suckling Piglets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reiner, G.; Lechner, M.; Eisenack, A.; Kallenbach, K.; Rau, K.; Müller, S.; Fink-Gremmels, J. Prevalence of an inflammation and necrosis syndrome in suckling piglets. Animal 2019, 13, 2007–2017. [Google Scholar] [CrossRef] [PubMed]
- Reiner, G.; Kühling, J.; Lechner, M.; Schrade, H.J.; Saltzmann, J.; Muelling, C.; Daenicke, S.; Loewenstein, F. Swine Inflammation and Necrosis Syndrome is influenced by husbandry and quality of sow in suckling piglets, weaners and fattening pigs. Porcine Health Manag. 2020, 6, 32. [Google Scholar] [CrossRef]
- Kühling, J.; Eisenhofer, K.; Lechner, M.; Becker, S.; Willems, H.; Reiner, G. The effects of boar on susceptibility to swine inflammation and necrosis syndrome in piglets. Porcine Health Manag. 2021, 7, 15. [Google Scholar] [CrossRef] [PubMed]
- EFSA; Panel on Animal Health and Welfare (AHAW). Statement on the use of animal-based measures to assess the welfare of animals. EFSA J. 2012, 10, 2767. [Google Scholar]
- EFSA. Scientific Opinion concerning a multifactorial approach on the use of animal and non-animal-based measures to assess the welfare of pigs. EFSA J. 2014, 12, 3702. [Google Scholar]
- Penny, R.H.C.; Edwards, M.J.; Mulley, R. Clinical observations of necrosis of skin of suckling piglets. Aust. Vet. J. 1971, 47, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Blowey, R.; Done, S.H. Tail necrosis in pigs. Pig J. 2003, 5L, 155–163. [Google Scholar]
- Santi, M.; Gheller, N.B.; Mores, T.J.; Marques, B.M.; Gonçalves, M.A.D.; Gava, D.; Zlotowski, P.; Driemeier, D.; Barcellos, D.E.S.N. Tail Necrosis in Piglets—Case Report. Available online: http://hdl.handle.net/11299/155093 (accessed on 15 November 2018).
- Reiner, G.; Lechner, M. Inflammation and necrosis syndrome (SINS) in swine. CAB Rev. 2019, 14, 1–8. [Google Scholar] [CrossRef]
- Schrauwen, E.; Thoonen, H.; Hoorens, J.; Houvenaghel, A. Pathophysiological effects of endotoxin infusion in young piglets. Br. Vet. J. 1986, 142, 364–370. [Google Scholar] [CrossRef]
- Guillou, D.; Demey, V.; Chacheyras-Durand, F.; Le Treut, Y. Mise en evidence du transfer des endotoxines de la truie vers sa portée dans le context du syndrome de dysgalactie post-partum. J. Rech. Porc. 2013, 45, 269–270. [Google Scholar]
- Van Limbergen, T.; Devreese, M.; Croubels, S.; Broekaert, N.; Michiels, A.; DeSaeger, S.; Maes, D. Role of mycotoxins in herds with and without problems with tail necrosis in neonatal pigs. Vet. Rec. 2017, 181, 539. [Google Scholar] [CrossRef]
- Nordgreen, J.; Edwards, S.A.; Boyle, L.A.; Bolhuis, J.E.; Veit, C.; Sayyari, A.; Marin, D.E.; Dimitrov, I.; Janczak, A.M.; Valros, A. A proposed role for proinflammatory cytokines in damaging behavior in pigs. Front. Vet. Sci. 2020, 7, 646. [Google Scholar] [CrossRef] [PubMed]
- Fox, E.S.; Thomas, P.; Broitman, S.A. Clearance of Gut-Derived Endotoxins by the Liver: Release and Modification of 3H, 14C-Lipopolysaccharide by Isolated Rat Kupffer Cells. Gastroenterology 1989, 96, 456–461. [Google Scholar] [CrossRef]
- Atif, M.S.; Warner, S.; Oo, Y.H. Linking the gut and liver: Crosstalk between regulatory T cells and mucosa-associated invariant T cells. Hepatol. Int. 2018, 12, 305–314. [Google Scholar] [CrossRef]
- Medzhitov, R. Toll-like receptors and innate immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Ravin, H.A.; Rowley, D.; Jenkins, C.; Fine, J. On the absorption of bacterial endotoxin from the gastro-intestinal tract of the normal and shocked animal. J. Exp. Med. 1960, 112, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Ganey, P.E.; Roth, R.A. Concurrent inflammation as a determinant of susceptibility to toxicity from xenobiotic agents. Toxicology 2001, 169, 195–208. [Google Scholar] [CrossRef]
- Pestka, J.J.; Zhou, H.R.; Moon, Y.; Chung, Y.J. Cellular and molecular mechanisms for immune modulation by deoxynivalenol and other trichothecenes: Unravelling a paradoxon. Toxicol. Lett. 2004, 153, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of two mycotoxins deoxynivalenol and fumonisin on pig intestinal health. Porcine Health Manag. 2016, 2, 21. [Google Scholar] [CrossRef] [PubMed]
- Gessner, D.K.; Winkler, A.; Koch, C.; Dusel, G.; Liebisch, G.; Ringseis, R.; Eder, K. Analysis of hepatic transcript profile and plasma lipid profile in early lactating dairy cows fed grape seed and grape marc meal extract. BMC Genom. 2017, 18, 253. [Google Scholar] [CrossRef]
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics 2003, 4, 249–264. [Google Scholar] [CrossRef]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef]
- Gessner, D.K.; Schwarz, A.; Meyer, S.; Wen, G.; Most, E.; Zorn, H.; Ringseis, R.; Eder, K. Insect Meal as Alternative Protein Source Exerts Pronounced Lipid-Lowering Effects in Hyperlipidemic Obese Zucker Rats. J. Nutr. 2019, 149, 566–577. [Google Scholar] [CrossRef]
- Ringseis, R.; Zeitz, J.O.; Weber, A.; Koch, C.; Eder, K. Hepatic transcript profiling in early-lactation dairy cows fed rumen-protected niacin during the transition from late pregnancy to lactation. J. Dairy Sci. 2019, 102, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Chiappisi, E.; Ringseis, R.; Eder, K.; Gessner, D.K. Effect of endoplasmic reticulum stress on metabolic and stress signaling and kidney-specific functions in Madin-Darby bovine kidney cells. J. Dairy Sci. 2017, 100, 6689–6706. [Google Scholar] [CrossRef] [PubMed]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. In Bioinformatics Methods and Protocols; Krawetz, S., Misener, S., Eds.; Humana Press: Totowa, NJ, USA, 2000; pp. 365–386. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef]
- Betz, P. Histological and enzyme histochemical parameters for the age estimation of human skin wounds. Int. J. Legal Med. 1994, 107, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.G.; Maubert, M.E.; Anton, L.; Heiser, L.M.; Elovitz, M.A. The tracking of lipopolysaccharide through the feto-maternal compartment and the involvement of maternal TLR4 in inflammation-induced fetal brain injury. Am. J. Reprod. Immunol. 2019, 82, e13189. [Google Scholar] [CrossRef]
- Puri, P.; Wiest, M.M.; Patnaik, M.; Cheung, O.; Sargeant, C.C.; Mirshahi, F.; Watkins, S.M.; Sanyal, A.Y. The plasma lipidomic signature of nonalcoholic steatohepatitis: Differential levels of n-3 and n-6 polyunsaturated fatty acids and their lipoxygenase products. Hepatology 2007, 46, 310A–311A. [Google Scholar]
- Allard, J.; Aghdassi, E.; Mohammed, S.; Raman, M.; Avand, G.; Arendt, B.M.; Jalali, P.; Kandasamy, T.; Prayitno, N.; Sherman, M.; et al. Nutritional assessment and hepatic fatty acid composition in non-alcoholic fatty liver disease (NAFLD): A cross-sectional study. J. Hepatol. 2008, 48, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Chiappini, F.; Coilly, A.; Kadar, H.; Gual, P.; Tran, A.; Desterke, C.; Samuel, D.; Duclos-Vallee, J.C.; Touboul, D.; Bertrand-Michel, J.; et al. Metabolism dysregulation induces a specific lipid signature of nonalcoholic steatohepatitis in patients. Sci. Rep. 2017, 7, 46658. [Google Scholar] [CrossRef] [PubMed]
- Song, M.J.; Malhi, H. The unfolded protein response and hepatic lipid metabolism in non-alcoholic fatty liver disease. Pharmacol. Ther. 2019, 203, 107401. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Part | Clinical Signs | SINS Low | SINS High | p-Value |
|---|---|---|---|---|
| Tail base | no bristles | 0.0 | 100.0 | 0.001 |
| swelling | 0.0 | 100.0 | 0.001 | |
| redness | 0.0 | 16.7 | ||
| exudation | 0.0 | 16.7 | ||
| necrosis | 0.0 | 0.0 | ||
| any | 0.0 | 100.0 | 0.001 | |
| Tail | swelling | 0.0 | 67.7 | 0.03 |
| scab formation | 0.0 | 83.3 | 0.008 | |
| rhagades | 0.0 | 67.7 | 0.03 | |
| exudation | 0.0 | 100.0 | 0.001 | |
| necrosis | 0.0 | 16.7 | ||
| annular constriction of tail tissue | 0.0 | 0.0 | ||
| bleeding | 0.0 | 0.0 | ||
| loss | 0.0 | 0.0 | ||
| any | 0.0 | 100.0 | 0.001 | |
| Ears | no bristles | 0.0 | 33.3 | |
| veins combusted | 0.0 | 83.3 | 0.008 | |
| necrosis | 0.0 | 0.0 | ||
| any | 0.0 | 100.0 | 0.001 | |
| Face | eye edema | 0.0 | 33.3 | |
| nasal edema | 0.0 | 16.7 | ||
| any | 0.0 | 33.3 | ||
| Teats | veins combusted | 0 | 33.3 | |
| swelling | 0 | 83.3 | 0.008 | |
| redness | 16.7 | 33.3 | ||
| necrosis | 0 | 16.7 | ||
| any | 16.7 | 100.0 | 0.008 | |
| Thighs | veins combusted | 33.3 | 83.3 | |
| Navel | inflammation | 50.0 | 33.3 | |
| Coronary bands | redness | 50.0 | 100.0 | |
| exudation | 66.7 | 100.0 | ||
| necrosis | 0.0 | 50.0 | ||
| any | 66.7 | 100.0 | ||
| Claw wall | bulging | 0.0 | 88.3 | 0.008 |
| bleeding | 0.0 | 0.0 | ||
| any | 0.0 | 88.3 | 0.008 | |
| Soles | redness | 0.0 | 100.0 | 0.001 |
| detachment | 0.0 | 16.7 | ||
| any | 0.0 | 100.0 | 0.001 | |
| Heels | swelling | 100.0 | 66.7 | |
| redness | 83.3 | 83.3 | ||
| cracks | 16.7 | 0.0 | ||
| detachment | 0.0 | 0.0 | ||
| any | 100.0 | 100.0 | ||
| SINS-Score | 0.8 ± 0.6 | 12.0 ± 1.2 | <0.001 |
| Gene Symbol | log2(FC) SINS High vs. SINS Low | p-Value | ||
|---|---|---|---|---|
| Microarray | qPCR | Microarray | qPCR | |
| FMO5 | 1.94 | 1.37 | 0.049 | 0.067 |
| AKR1C1 | 1.73 | 1.57 | 0.020 | 0.017 |
| ESM1 | 1.43 | 2.38 | 0.015 | 0.016 |
| FGF14 | 1.30 | 2.47 | 0.004 | 0.017 |
| AKR1C2 | 1.20 | 0.90 | 0.008 | 0.097 |
| EPB42 | 1.17 | 1.52 | 0.004 | 0.037 |
| HEMGN | 1.02 | 1.70 | 0.012 | 0.110 |
| CRP | 0.97 | 1.28 | 0.025 | 0.053 |
| BCO2 | 0.93 | 1.10 | 0.009 | 0.040 |
| C3 | 0.91 | 0.93 | 0.030 | 0.080 |
| MPO | 0.90 | 1.79 | 0.026 | 0.086 |
| XDH | 0.88 | 1.48 | 0.023 | 0.009 |
| CYP2E1 | 0.87 | 1.51 | 0.039 | 0.012 |
| MVK | 0.77 | 0.93 | 0.001 | 0.001 |
| CP | 0.75 | 1.23 | 0.040 | 0.011 |
| GINS1 | −1.25 | −0.93 | 0.026 | 0.054 |
| SLC51B | −1.19 | −1.46 | 0.012 | 0.022 |
| PCLAF | −1.16 | −0.85 | 0.045 | 0.120 |
| STEAP2 | −1.11 | −0.86 | 0.046 | 0.134 |
| ARL2BP | −1.10 | −0.88 | 0.001 | 0.029 |
| ENPP4 | −1.06 | −0.48 | 0.006 | 0.278 |
| DNAJC9 | −1.04 | −0.69 | 0.020 | 0.077 |
| SKA1 | −1.00 | −0.30 | 0.033 | 0.487 |
| NDUFAF4 | −1.00 | −0.49 | 0.003 | 0.153 |
| CCL2 | −0.93 | −0.40 | 0.028 | 0.363 |
| MUC13 | −1.01 | −1.37 | 0.018 | 0.085 |
| ATP6V0D2 | −1.00 | −0.58 | 0.020 | 0.085 |
| RANBP1 | −0.98 | −0.70 | 0.005 | 0.067 |
| CYCS | −0.94 | −0.55 | <0.001 | 0.250 |
| LRRC59 | −0.86 | −0.79 | 0.046 | 0.123 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ringseis, R.; Gessner, D.K.; Loewenstein, F.; Kuehling, J.; Becker, S.; Willems, H.; Lechner, M.; Eder, K.; Reiner, G. Swine Inflammation and Necrosis Syndrome Is Associated with Plasma Metabolites and Liver Transcriptome in Affected Piglets. Animals 2021, 11, 772. https://doi.org/10.3390/ani11030772
Ringseis R, Gessner DK, Loewenstein F, Kuehling J, Becker S, Willems H, Lechner M, Eder K, Reiner G. Swine Inflammation and Necrosis Syndrome Is Associated with Plasma Metabolites and Liver Transcriptome in Affected Piglets. Animals. 2021; 11(3):772. https://doi.org/10.3390/ani11030772
Chicago/Turabian StyleRingseis, Robert, Denise K. Gessner, Frederik Loewenstein, Josef Kuehling, Sabrina Becker, Hermann Willems, Mirjam Lechner, Klaus Eder, and Gerald Reiner. 2021. "Swine Inflammation and Necrosis Syndrome Is Associated with Plasma Metabolites and Liver Transcriptome in Affected Piglets" Animals 11, no. 3: 772. https://doi.org/10.3390/ani11030772
APA StyleRingseis, R., Gessner, D. K., Loewenstein, F., Kuehling, J., Becker, S., Willems, H., Lechner, M., Eder, K., & Reiner, G. (2021). Swine Inflammation and Necrosis Syndrome Is Associated with Plasma Metabolites and Liver Transcriptome in Affected Piglets. Animals, 11(3), 772. https://doi.org/10.3390/ani11030772