
Effect of Chitosan Inclusion and Dietary Crude Protein Level on Nutrient Intake and Digestibility, Ruminal Fermentation, and N Excretion in Beef Heifers Offered a Grass Silage Based Diet


 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Dietary Treatments

{kind=link}
{kind=link}
{kind=link}
| Protein Level 1 | HP | LP | ||
|---|---|---|---|---|
| Chitosan Inclusion 2 | + | − | + | − |
| Ingredient composition, % DM | ||||
| Grass silage | 50 | 50 | 50 | 50 |
| Rolled barley | 26.75 | 26.75 | 31.55 | 31.55 |
| Protein mix 3 | 20.50 | 20.50 | 5.7 | 5.7 |
| Soyhulls | − | − | 10 | 10 |
| Molasses | 1.5 | 1.5 | 1.5 | 1.5 |
| Mineral mix 4 | 1.25 | 1.25 | 1.25 | 1.25 |
| Chemical composition, % DM | ||||
| Dry matter % | 35.95 | 35.79 | 35.58 | 35.39 |
| Crude protein | 16.38 | 16.30 | 13.27 | 13.30 |
| RDP 5 | 11.10 | 11.10 | 9.25 | 9.25 |
| RUP 5 | 5.28 | 5.28 | 3.80 | 3.80 |
| Neutral detergent fibre | 36.77 | 36.03 | 39.03 | 37.54 |
| Neutral detergent fibre forage | 51.24 | 51.24 | 51.24 | 51.24 |
| Acid detergent fibre | 22.20 | 21.89 | 24.52 | 23.91 |
| Starch | 11.96 | 11.68 | 13.77 | 14.10 |
| Ether extract | 2.87 | 2.49 | 2.12 | 2.59 |
| Gross energy, MJ/kg | 16.65 | 16.62 | 16.44 | 16.27 |
2.2. Data and Sample Collection
2.3. Chemical Analysis
2.4. Statistical Analyses
3. Results
3.1. Nutrient Intake and Digestibility
3.2. Nitrogen Intake and Output
3.3. Blood Metabolites
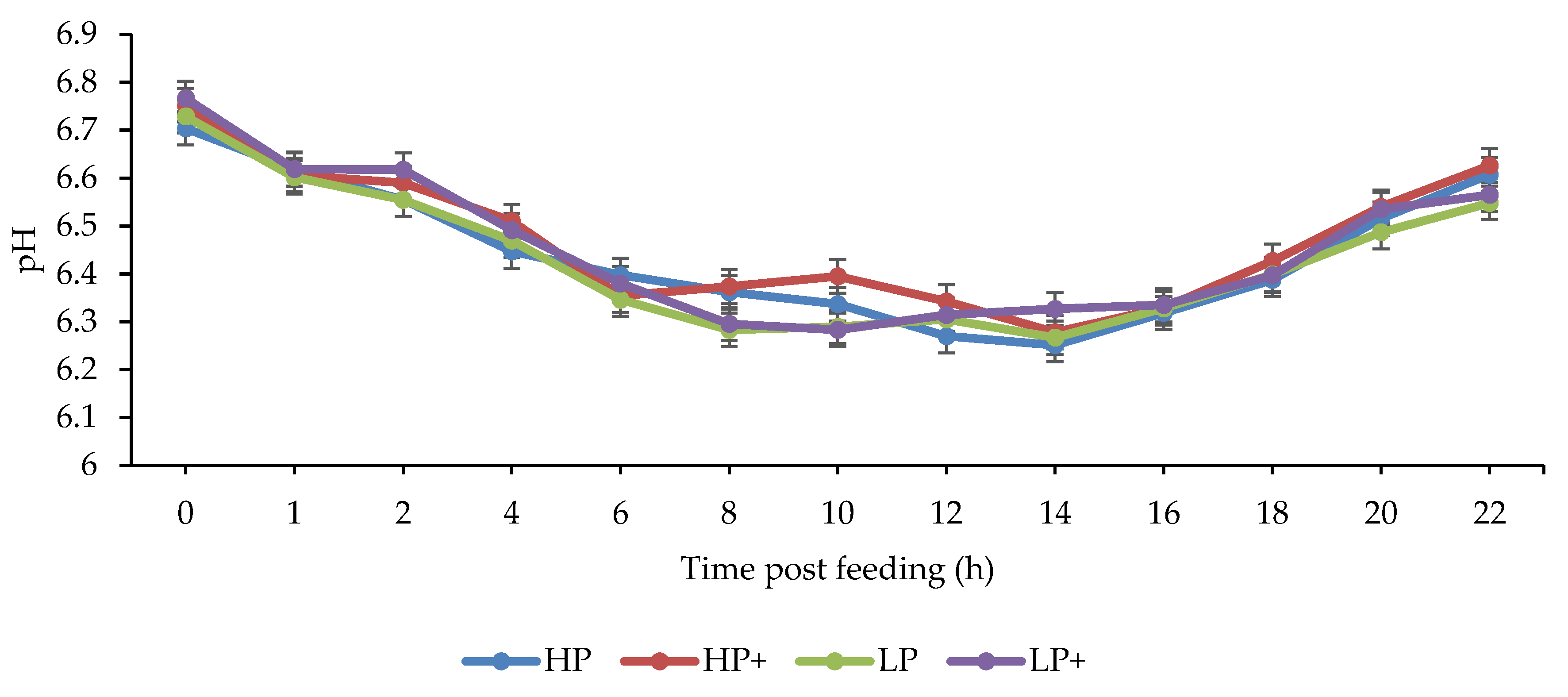
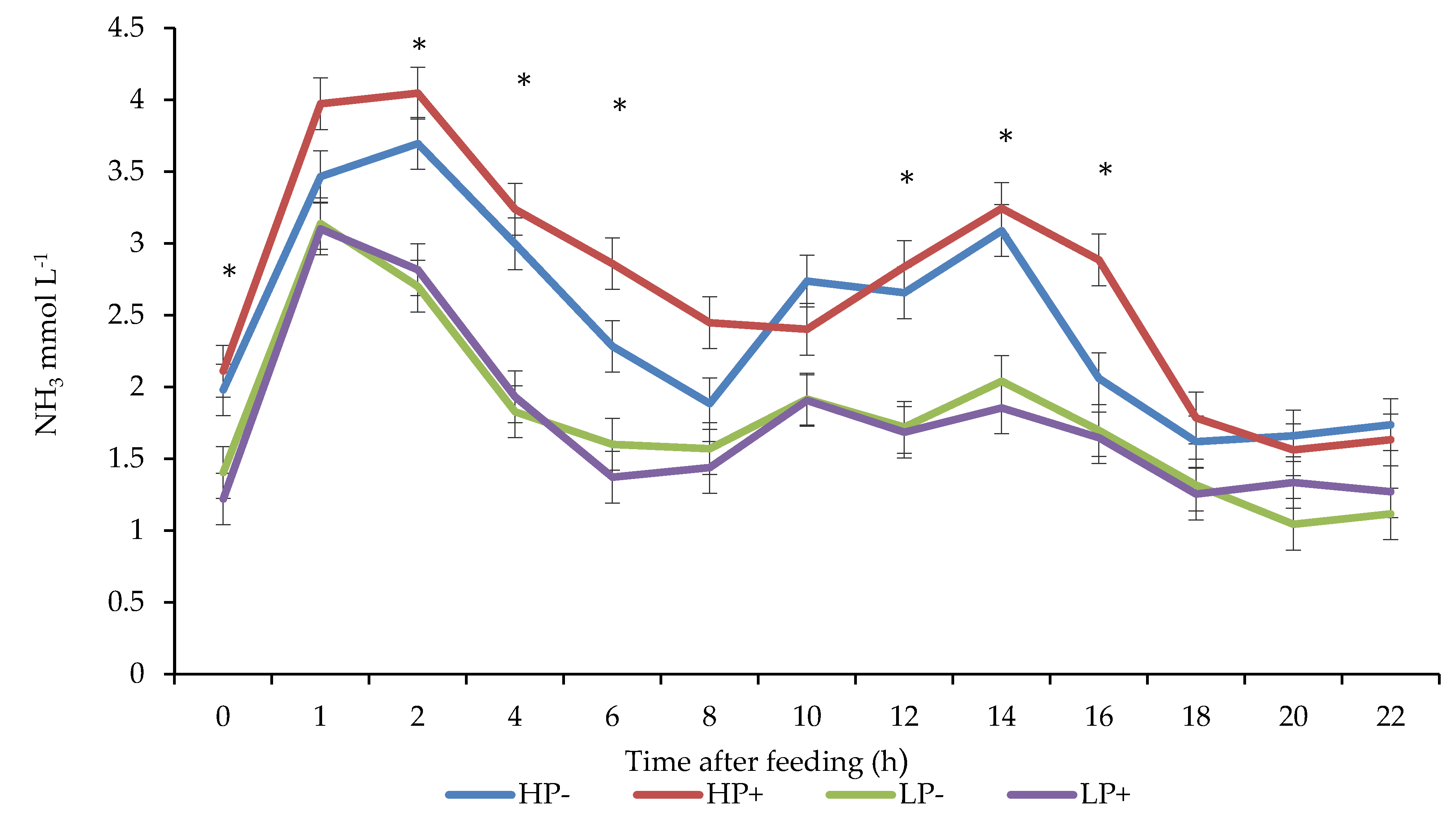
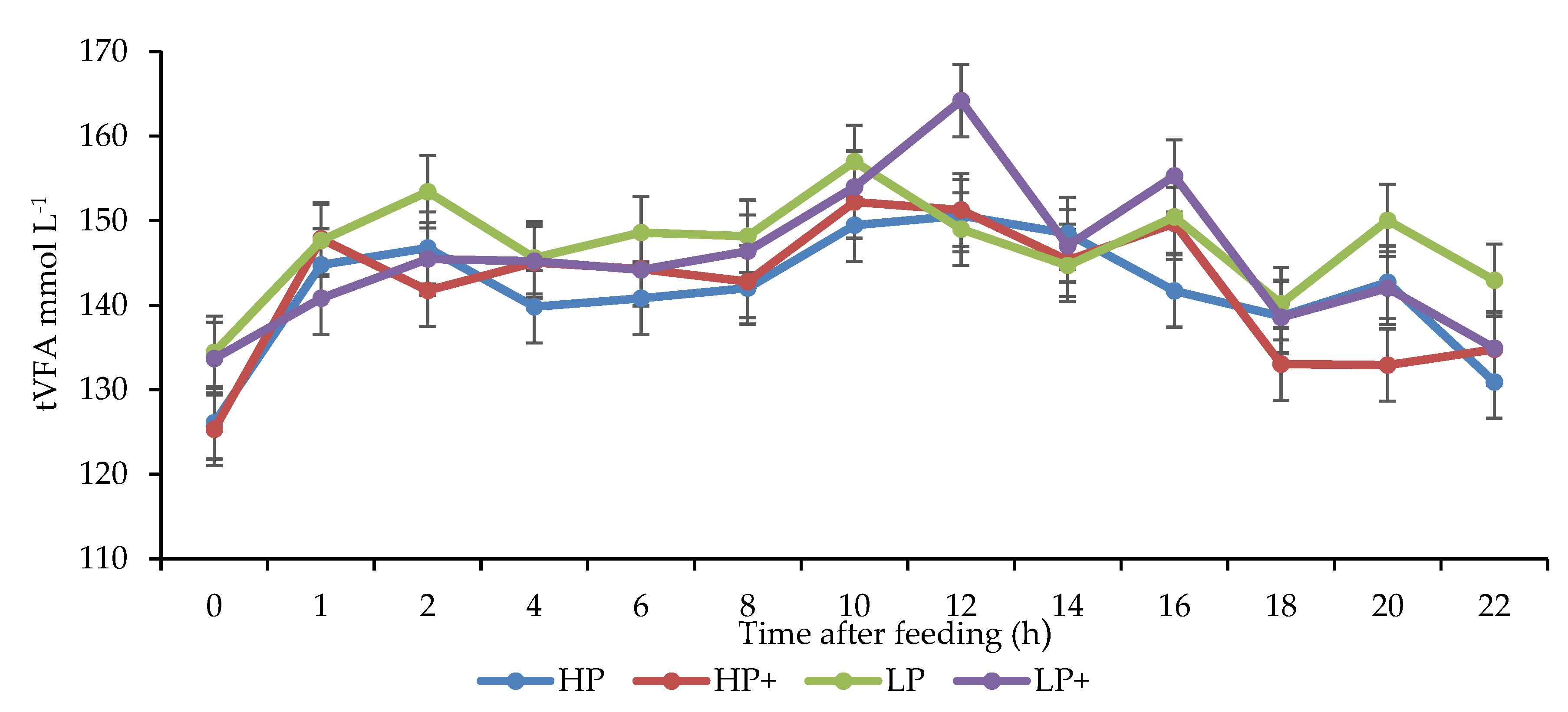
3.4. Rumen Fermentation Parameters
4. Discussion
4.1. Nutrient Intake and Digestibility
4.2. Nitrogen Intake and Output
4.3. Rumen Fermentation Parameters
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Duffy, P.; Hyde, B.; Ryan, A.M.; Murphy, J.; Quirke, B.; Fahey, D. Air Pollutant Emissions In Ireland 1990–2017 Reported to the Secretariat of the UNECE Convention on Long-Range Transboundary Air Pollution And To The European Union; Environmental Protection Agency: Washington, DC, USA, 2019; ISBN 978-1-84095-817-1.
- Environmental Protection Agency. Ireland’s Environment—An Assessment 2016; Environmental Protection Agency: Washington, DC, USA; Johnstown Castle Estate, Co.: Wexford, Ireland, 2016.
- Warner, J.X.; Dickerson, R.R.; Wei, Z.; Strow, L.L.; Wang, Y.; Liang, Q. Increased atmospheric ammonia over the world′s major agricultural areas detected from space. Geophys. Res. Lett. 2017, 44, 2875–2884. [Google Scholar] [CrossRef] [PubMed]
- Aneja, V.P.; Schlesinger, W.H.; Li, Q.; Nahas, A.; Battye, W.H. Characterization of the Global Sources of Atmospheric Ammonia from Agricultural Soils. J. Geophys. Res. Atmos. 2020, 125, e2019JD031684. [Google Scholar] [CrossRef]
- Duffy, P.; Black, K.; Fahey, D.; Hyde, B.; Kehoe, A.; Murphy, J.; Quirke, B.; Ryan, A.M.; Ponzi, J. Greenhouse Gas Emissions 1990–2016 Reported to the United Nations Framework Convention on Climate Change; Environmental Protection Agency: Washington, DC, USA, 2020; ISBN 978-1-84095-884-3.
- EEA. National Emission Ceilings Directive. Available online: https://www.eea.europa.eu/themes/air/air-pollution-sources-1/national-emissionceilings/national-emission-ceilings-directive (accessed on 30 September 2020).
- Angelidis, A.; Crompton, L.; Misselbrook, T.; Yan, T.; Reynolds, C.; Stergiadis, S. Evaluation and prediction of nitrogen use efficiency and outputs in faeces and urine in beef cattle. Agric. Ecosyst. Environ. 2019, 280, 1–15. [Google Scholar] [CrossRef]
- Cole, N.A.; Todd, R.W. Nitrogen and phosphorus balance of beef cattle feedyards. In Proceedings of the Texas Animal Manure Management Issues Conference, Round Rock, TX, USA, 29–30 September 2009.
- Erickson, G.; Klopfenstein, T. Nutritional and management methods to decrease nitrogen losses from beef feedlots. J. Anim. Sci. 2010, 88, E172–E180. [Google Scholar] [CrossRef]
- Todd, R.W.; Cole, N.A.; Hagevoort, G.R.; Casey, K.D.; Auvermann, B.W. Ammonia losses and nitrogen partitioning at a southern High Plains open lot dairy. Atmos. Environ. 2015, 110, 75–83. [Google Scholar] [CrossRef]
- Jarvis, S.; Hatch, D.; Lockyer, D. Ammonia fluxes from grazed grassland: Annual losses from cattle production systems and their relation to nitrogen inputs. J. Agric. Sci. 1989, 113, 99–108. [Google Scholar] [CrossRef]
- Varel, V.H.; Nienaber, J.A.; Freetly, H.C. Conservation of nitrogen in cattle feedlot waste with urease inhibitors. J. Anim. Sci. 1999, 77, 1162–1168. [Google Scholar] [CrossRef]
- Mulligan, F.; Dillon, P.; Callan, J.; Rath, M.; O’mara, F. Supplementary concentrate type affects nitrogen excretion of grazing dairy cows. J. Dairy Sci. 2004, 87, 3451–3460. [Google Scholar] [CrossRef]
- Cole, N.A.; Defoor, P.J.; Galyean, M.L.; Duff, G.C.; Gleghorn, J.F. Effects of phase-feeding of crude protein on performance, carcass characteristics, serum urea nitrogen concentrations, and manure nitrogen of finishing beef steers12. J. Anim. Sci. 2006, 84, 3421–3432. [Google Scholar] [CrossRef]
- Vasconcelos, J.T.; Cole, N.A.; McBride, K.W.; Gueye, A.; Galyean, M.L.; Richardson, C.R.; Greene, L.W. Effects of dietary crude protein and supplemental urea levels on nitrogen and phosphorus utilization by feedlot cattle1. J. Anim. Sci. 2009, 87, 1174–1183. [Google Scholar] [CrossRef]
- da Silva, L.D.; Pereira, O.G.; da Silva, T.C.; Valadares Filho, S.C.; Ribeiro, K.G. Effects of silage crop and dietary crude protein levels on digestibility, ruminal fermentation, nitrogen use efficiency, and performance of finishing beef cattle. Anim. Feed Sci. Technol. 2016, 220, 22–33. [Google Scholar] [CrossRef]
- Kong, M.; Chen, X.G.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food Microbiol. 2010, 144, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Mehdizadeh, T.; Tajik, H.; Langroodi, A.M.; Molaei, R.; Mahmoudian, A. Chitosan-starch film containing pomegranate peel extract and Thymus kotschyanus essential oil can prolong the shelf life of beef. Meat Sci. 2020, 163, 108073. [Google Scholar] [CrossRef] [PubMed]
- Duran, A.; Kahve, H.I. The effect of chitosan coating and vacuum packaging on the microbiological and chemical properties of beef. Meat Sci. 2020, 162, 107961. [Google Scholar] [CrossRef]
- Belanche, A.; Ramos-Morales, E.; Newbold, C.J. In vitro screening of natural feed additives from crustaceans, diatoms, seaweeds and plant extracts to manipulate rumen fermentation. J. Sci. Food Agric. 2016, 96, 3069–3078. [Google Scholar] [CrossRef]
- Dias, A.O.C.; Goes, R.H.T.B.; Gandra, J.R.; Takiya, C.S.; Branco, A.F.; Jacaúna, A.G.; Oliveira, R.T.; Souza, C.J.S.; Vaz, M.S.M. Increasing doses of chitosan to grazing beef steers: Nutrient intake and digestibility, ruminal fermentation, and nitrogen utilization. Anim. Feed Sci. Technol. 2017, 225, 73–80. [Google Scholar] [CrossRef]
- Goiri, I.; Oregui, L.M.; Garcia-Rodriguez, A. Dose–response effects of chitosans on in vitro rumen digestion and fermentation of mixtures differing in forage-to-concentrate ratios. Anim. Feed Sci. Technol. 2009, 151, 215–227. [Google Scholar] [CrossRef]
- Goiri, I.; Garcia-Rodriguez, A.; Oregui, L.M. Effect of chitosans on in vitro rumen digestion and fermentation of maize silage. Anim. Feed Sci. Technol. 2009, 148, 276–287. [Google Scholar] [CrossRef]
- Belanche, A.; Pinloche, E.; Preskett, D.; Newbold, C.J. Effects and mode of action of chitosan and ivy fruit saponins on the microbiome, fermentation and methanogenesis in the rumen simulation technique. FEMS Microbiol. Ecol. 2016, 92. [Google Scholar] [CrossRef] [PubMed]
- Goiri, I.; Oregui, L.; Garcia-Rodriguez, A. Use of chitosans to modulate ruminal fermentation of a 50: 50 forage-to-concentrate diet in sheep. J. Anim. Sci. 2010, 88, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Araújo, A.; Venturelli, B.; Santos, M.; Gardinal, R.; Cônsolo, N.; Calomeni, G.; Freitas, J.; Barletta, R.; Gandra, J.; Paiva, P. Chitosan affects total nutrient digestion and ruminal fermentation in Nellore steers. Anim. Feed Sci. Technol. 2015, 206, 114–118. [Google Scholar] [CrossRef]
- de Paiva, P.G.; de Jesus, E.F.; Del Valle, T.A.; de Almeida, G.F.; Costa, A.G.B.V.B.; Consentini, C.E.C.; Zanferari, F.; Takiya, C.S.; da Silva Bueno, I.C.; Rennó, F.P. Effects of chitosan on ruminal fermentation, nutrient digestibility, and milk yield and composition of dairy cows. Anim. Prod. Sci. 2017, 57, 301–307. [Google Scholar] [CrossRef]
- Mingoti, R.; Freitas Jr, J.; Gandra, J.; Gardinal, R.; Calomeni, G.; Barletta, R.; Vendramini, T.; Paiva, P.; Rennó, F. Dose response of chitosan on nutrient digestibility, blood metabolites and lactation performance in holstein dairy cows. Livest. Sci. 2016, 187, 35–39. [Google Scholar] [CrossRef]
- Del Valle, T.A.; de Paiva, P.G.; de Jesus, E.F.; de Almeida, G.F.; Zanferari, F.; Costa, A.G.; Bueno, I.C.; Rennó, F.P. Dietary chitosan improves nitrogen use and feed conversion in diets for mid-lactation dairy cows. Livest. Sci. 2017, 201, 22–29. [Google Scholar] [CrossRef]
- Vendramini, T.H.A.; Takiya, C.S.; Silva, T.; Zanferari, F.; Rentas, M.F.; Bertoni, J.; Consentini, C.E.C.; Gardinal, R.; Acedo, T.S.; Rennó, F.P. Effects of a blend of essential oils, chitosan or monensin on nutrient intake and digestibility of lactating dairy cows. Anim. Feed Sci. Technol. 2016, 214, 12–21. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Council, N.R. Nutrient Requirements of Dairy Cattle: Seventh Revised Edition, 2001; The National Academies Press: Washington, DC, USA, 2001; p. 405. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis; AOAC: Arlington, VA, USA, 1990. [Google Scholar]
- McCleary, B.; Solah, V.; Gibson, T. Quantitative measurement of total starch in cereal flours and products. J. Cereal Sci. 1994, 20, 51–58. [Google Scholar] [CrossRef]
- Van Soest, P.V.; Robertson, J.; Lewis, B. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Weatherburn, M. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Wencelova, M.; Varadyova, Z.; Mihalikova, K.; Kisidayova, S.; Jalc, D. Evaluating the effects of chitosan, plant oils, and different diets on rumen metabolism and protozoan population in sheep. Turk. J. Vet. Anim. Sci. 2014, 38, 26–33. [Google Scholar] [CrossRef]
- Bach, A.; Calsamiglia, S.; Stern, M.D. Nitrogen metabolism in the rumen. J. Dairy Sci. 2005, 88, E9–E21. [Google Scholar] [CrossRef]
- Ivan, M. Comparison of duodenal flow and digestibility in fauna-free sheep inoculated with Holotrich protozoa, Entodinium monofauna or total mixed protozoa population. Br. J. Nutr. 2008, 101, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Garrido-Maestu, A.; Jeong, K.C. Application, mode of action, and in vivo activity of chitosan and its micro-and nanoparticles as antimicrobial agents: A review. Carbohydr. Polym. 2017, 176, 257–265. [Google Scholar] [CrossRef]
- Tamminga, S. A review on environmental impacts of nutritional strategies in ruminants. J. Anim. Sci. 1996, 74, 3112–3124. [Google Scholar] [CrossRef] [PubMed]
- Colmenero, J.O.; Broderick, G. Effect of dietary crude protein concentration on milk production and nitrogen utilization in lactating dairy cows. J. Dairy Sci. 2006, 89, 1704–1712. [Google Scholar] [CrossRef]
- Reynolds, C.K.; Kristensen, N.B. Nitrogen recycling through the gut and the nitrogen economy of ruminants: An asynchronous symbiosis1. J. Anim. Sci. 2008, 86, E293–E305. [Google Scholar] [CrossRef] [PubMed]
- Menezes, A.; Valadares Filho, S.; de Silva, L.C.; Pacheco, M.; Pereira, J.; Rotta, P.; Zanetti, D.; Detmann, E.; Silva, F.; Godoi, L. Does a reduction in dietary crude protein content affect performance, nutrient requirements, nitrogen losses, and methane emissions in finishing Nellore bulls? Agric. Ecosyst. Environ. 2016, 223, 239–249. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Dairy Cattle 6; National Academy Sciences: Washington, DC, USA, 1985.
- Mould, F.; Ørskov, E.; Gauld, S.A. Associative effects of mixed feeds. II. The effect of dietary addition of bicarbonate salts on the voluntary intake and digestibility of diets containing various proportions of hay and barley. Anim. Feed Sci. Technol. 1983, 10, 31–47. [Google Scholar] [CrossRef]
- Dijkstra, J.; Ellis, J.; Kebreab, E.; Strathe, A.; López, S.; France, J.; Bannink, A. Ruminal pH regulation and nutritional consequences of low pH. Anim. Feed Sci. Technol. 2012, 172, 22–33. [Google Scholar] [CrossRef]
- Sannes, R.; Messman, M.; Vagnoni, D. Form of Rumen-Degradable Carbohydrate and Nitrogen on Microbial Protein Synthesis and Protein Efficiency of Dairy Cows1. J. Dairy Sci. 2002, 85, 900–908. [Google Scholar] [CrossRef]
- Allison, M. Nitrogen metabolism of ruminal micro-organisms. In Physiology of Digestion and Metabolism in the Ruminant; Oriel Press: Newcastle upon Tyne, UK, 1970; pp. 456–463. [Google Scholar]
- Kang-Meznarich, J.H.; Broderick, G.A. Effects of Incremental Urea Supplementation on Ruminal Ammonia Concentration and Bacterial Protein Formation2. J. Anim. Sci. 1980, 51, 422–431. [Google Scholar] [CrossRef]
- Chen, H.-C.; Chang, C.-C.; Mau, W.-J.; Yen, L.-S. Evaluation of N-acetylchitooligosaccharides as the main carbon sources for the growth of intestinal bacteria. FEMS Microbiol. Lett. 2002, 209, 53–56. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Haraki, H.; Gandra, J.; Oliveira, E.; Takiya, C.; Goes, R.; Gabriel, A.; Rodrigues, G.; Gandra, E.; Pereira, T.; Damiani, J. Effects of chitosan and whole raw soybeans on feeding behavior and heat losses of Jersey heifers. Iran. J. Appl. Anim. Sci. 2018, 8, 397–405. [Google Scholar]
- Dado, R.; Allen, M. Intake limitations, feeding behavior, and rumen function of cows challenged with rumen fill from dietary fiber or inert bulk. J. Dairy Sci. 1995, 78, 118–133. [Google Scholar] [CrossRef]
- Baumann, T.A.; Lardy, G.P.; Caton, J.S.; Anderson, V.L. Effect of energy source and ruminally degradable protein addition on performance of lactating beef cows and digestion characteristics of steers1. J. Anim. Sci. 2004, 82, 2667–2678. [Google Scholar] [CrossRef] [PubMed][Green Version]



| Crude Protein 1 | HP | LP | SEM | CP | CHI | CP×CHI | ||
|---|---|---|---|---|---|---|---|---|
| Chitosan2 | + | − | + | − | ||||
| DMI (kg d−1) | 12.31 | 13.14 | 12.14 | 12.33 | 0.373 | 0.206 | 0.183 | 0.395 |
| Apparent total tract digestibility % | ||||||||
| Dry matter | 67.50 | 69.95 | 67.22 | 69.29 | 0.818 | 0.427 | <0.001 | 0.745 |
| Organic matter | 69.97 | 72.27 | 70.03 | 71.78 | 0.504 | 0.675 | <0.001 | 0.589 |
| Crude protein | 68.83 | 71.47 | 61.36 | 64.38 | 0.009 | <0.001 | 0.003 | 0.826 |
| NDF 3 | 48.00 | 50.59 | 49.08 | 50.59 | 1.313 | 0.688 | 0.133 | 0.689 |
| Nitrogen | ||||||||
| Intake (g d 1) | 323.8 | 327.9 | 262.3 | 261.9 | 0.009 | <0.001 | 0.850 | 0.819 |
| Output (g−1) | ||||||||
| Urine | 166.1 | 159.8 | 106.1 | 113.8 | 0.009 | <0.001 | 0.942 | 0.444 |
| Faecal | 100.4 | 93.5 | 101.9 | 93.8 | 0.003 | 0.797 | 0.041 | 0.865 |
| Total | 266.5 | 253.3 | 208.1 | 207.6 | 0.010 | <0.001 | 0.502 | 0.534 |
| % total excreted4 | ||||||||
| % urine | 62.15 | 62.51 | 51.09 | 54.76 | 1.790 | <0.001 | 0.272 | 0.366 |
| % faecal | 37.85 | 37.49 | 48.91 | 45.24 | 1.790 | <0.001 | 0.272 | 0.366 |
| Nitrogen recovery5 | ||||||||
| Urine | 0.52 | 0.49 | 0.41 | 0.44 | 0.029 | 0.008 | 0.976 | 0.291 |
| Faecal | 0.31 | 0.28 | 0.39 | 0.36 | 0.009 | <0.001 | 0.003 | 0.826 |
| Blood metabolites | ||||||||
| Urea 6 | 5.48 | 5.31 | 4.08 | 3.88 | 0.147 | <0.001 | 0.209 | 0.933 |
| Creatinine 7 | 116.9 | 116.7 | 123.6 | 119.5 | 2.145 | 0.031 | 0.312 | 0.371 |
| Total Protein 8 | 81.45 | 81.63 | 79.91 | 81.84 | 1.419 | 0.636 | 0.462 | 0.534 |
| Glucose 6 | 3.60 | 3.56 | 3.61 | 3.57 | 0.078 | 0.911 | 0.611 | 0.956 |
| Crude Protein 1 | HP | LP | SEM | CP | CHI | CP × CHI | ||
|---|---|---|---|---|---|---|---|---|
| Chitosan 2 | + | − | + | − | ||||
| pH | 6.47 | 6.44 | 6.45 | 6.43 | 0.025 | 0.163 | 0.002 | 0.870 |
| mmol L−1 | ||||||||
| NH3 | 2.69 a | 2.45 b | 1.75 c,d | 1.78 d | 0.084 | <0.001 | 0.023 | 0.004 |
| Acetic | 111.2 | 110.7 | 115.3 | 117.3 | 1.62 | <0.001 | 0.497 | 0.240 |
| Propionic | 14.26 | 14.36 | 13.70 | 13.44 | 0.234 | <0.001 | 0.564 | 0.200 |
| Butyric | 9.24 | 9.33 | 9.00 | 9.25 | 0.206 | 0.338 | 0.355 | 0.633 |
| Iso-butyric | 1.87 a | 1.75 a | 1.68 b | 1.86 a,b | 0.113 | 0.490 | 0.792 | 0.052 |
| Valeric | 2.23 | 2.23 | 2.20 | 2.21 | 0.054 | 0.259 | 0.799 | 0.849 |
| Iso-valeric | 3.51 | 3.48 | 3.66 | 3.61 | 0.092 | 0.474 | 0.930 | 0.956 |
| Total VFA 3 | 142.2 | 141.8 | 145.6 | 147.1 | 1.98 | <0.001 | 0.640 | 0.449 |
| Ac:Pr 4 | 8.51 a | 7.80 a | 9.07 b | 9.58 b | 0.586 | <0.001 | 0.403 | 0.005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirwan, S.F.; Pierce, K.M.; Serra, E.; McDonald, M.; Rajauria, G.; Boland, T.M. Effect of Chitosan Inclusion and Dietary Crude Protein Level on Nutrient Intake and Digestibility, Ruminal Fermentation, and N Excretion in Beef Heifers Offered a Grass Silage Based Diet. Animals 2021, 11, 771. https://doi.org/10.3390/ani11030771
Kirwan SF, Pierce KM, Serra E, McDonald M, Rajauria G, Boland TM. Effect of Chitosan Inclusion and Dietary Crude Protein Level on Nutrient Intake and Digestibility, Ruminal Fermentation, and N Excretion in Beef Heifers Offered a Grass Silage Based Diet. Animals. 2021; 11(3):771. https://doi.org/10.3390/ani11030771
Chicago/Turabian StyleKirwan, Stuart F., Karina M. Pierce, Eleonora Serra, Michael McDonald, Gaurav Rajauria, and Tommy M. Boland. 2021. "Effect of Chitosan Inclusion and Dietary Crude Protein Level on Nutrient Intake and Digestibility, Ruminal Fermentation, and N Excretion in Beef Heifers Offered a Grass Silage Based Diet" Animals 11, no. 3: 771. https://doi.org/10.3390/ani11030771
APA StyleKirwan, S. F., Pierce, K. M., Serra, E., McDonald, M., Rajauria, G., & Boland, T. M. (2021). Effect of Chitosan Inclusion and Dietary Crude Protein Level on Nutrient Intake and Digestibility, Ruminal Fermentation, and N Excretion in Beef Heifers Offered a Grass Silage Based Diet. Animals, 11(3), 771. https://doi.org/10.3390/ani11030771